猪伪狂犬病毒gD蛋白抗原表位的克隆及表达
2016-02-06王寅彪邓瑞广李学伍河南科技大学动物科技学院河南洛阳47003河南省农业科学院河南省动物免疫学重点实验室河南郑州45000
刘 芳,张 冲,,王寅彪,赵 朴,邓瑞广,李学伍*(.河南科技大学 动物科技学院,河南 洛阳 47003; .河南省农业科学院/河南省动物免疫学重点实验室,河南 郑州 45000)
猪伪狂犬病毒gD蛋白抗原表位的克隆及表达
刘 芳1,张 冲1,2,王寅彪2,赵 朴2,邓瑞广2,李学伍2*
(1.河南科技大学 动物科技学院,河南 洛阳 471003; 2.河南省农业科学院/河南省动物免疫学重点实验室,河南 郑州 450002)
依据GenBank中猪伪狂犬病毒(PRV) BJ/YT株gD基因设计引物,扩增gD蛋白抗原表位,其设计扩增长度为702 bp,将扩增片段克隆到pMD19-T载体获得pMD19-T-gD。 利用BamHⅠ和Hind Ⅲ限制性内切酶双酶切pMD19-T-gD质粒,进行琼脂糖凝胶电泳并回收酶切片段,将回收片段与原核表达载体pET-28a连接,成功构建了pET-28a-gD表达载体。将表达载体转化感受态细胞Rosetta,IPTG诱导表达后,进行SDS-PAGE电泳,结果显示,在25 ku处出现特异性蛋白质条带。Western blot分析结果表明,表达产物能够被PRV的阳性血清识别。综上,成功克隆并表达了gD蛋白抗原表位,且其表达产物具有较好的生物学活性。
猪伪狂犬病病毒;gD基因; 抗原表位; 蛋白质表达
猪伪狂犬病(PR)是严重威胁我国养猪业健康发展的主要传染病之一[1-3]。临床表现为妊娠母猪的繁殖功能障碍和新生仔猪的体温升高、脑脊髓炎及神经症状。患病仔猪的死亡率较高,有时甚至高达100%[4-7]。成年猪感染后多能耐过,无明显临床症状,但终生带毒且成为最危险的潜在传染源。猪群一旦感染猪伪狂犬病毒(PRV)将很难根除。因此,伪狂犬病被列为生猪产业的第三大疫病[8]。
PRV属于孢疹病毒科,猪孢疹病毒属,可在多种组织细胞如扁桃体、鼻黏膜、局部淋巴结和肺脏等组织中增殖[9]。PRV是线性双股DNA病毒,基因组全长约150 kb,由长独特区(UL)、短独特区(US)及位于US两端的末端重复序列(TR)和内部重复序列(IR)组成[10],基因组中平均GC含量高达74%且结构复杂。目前已发现11种PRV糖蛋白[11],其中gD糖蛋白是成熟的PRV粒子囊膜表面的主要生物学活性糖蛋白,也是PRV增殖所必需的糖蛋白,在病毒粒子与宿主细胞间的不可逆吸附过程中发挥着重要的作用[12]。现已有多个PRV毒株的gD序列在GenBank登录,其全长约为1 250 bp,GC含量为74.5%,能够编码约400个氨基酸的糖蛋白[13]。鉴于PRV gD蛋白的重要性,本研究表达了gD抗原表位,并测定了其生物学活性,旨在为猪伪狂犬病疫苗免疫效果检测方法的建立奠定基础。
1 材料和方法
1.1 材料
1.1.1 菌种和载体 大肠杆菌DH5α感受态细胞购自天根生物(北京)有限公司,pMD19-T 克隆载体购自宝生物工程(大连)有限公司。表达载体pET-28a、Rosetta感受态细胞、PRV阳性血清及阴性血清、羊抗猪IgG、His单抗等均由河南省动物免疫学重点实验室保存。
1.1.2 主要试剂 DNA提取试剂盒,DNA胶回收试剂盒,LATaqDNA聚合酶,ExTap预混酶,限制性内切酶BamHⅠ、HindⅢ,dNTP及2×GC Buffer等均购自宝生物工程(大连)有限公司。
1.2 方法
1.2.1 PRV 基因组DNA的制备 PK-15 细胞形成单层后,弃去生长液,接种 PRV病毒液200 μL,吸附2 h后加维持液,当70%细胞出现病变和脱落时,收集所有细胞培养物。取500 μL细胞培养物,加入94 μL 10%(m/V)SDS、6 μL蛋白酶K,颠倒混匀,55 ℃水浴加热30 min;再加入等体积酚-氯仿,颠倒混匀,12 000 r/min离心5 min;取上清,再加入等体积氯仿,颠倒混匀,12 000 r/min离心5 min;取上清加入2倍体积冷乙醇颠倒混匀,-20 ℃静置10~20 min,12 000 r/min离心10 min;弃去上清,沉淀用70%酒精洗涤1次,置超净台挥发干净;用25 μL含RNase (20 μg/mL) 的灭菌去离子水溶解,立即使用或置-20 ℃保存。
1.2.2 PRV基因组DNA的鉴定 根据文献[14]的方法,在gD基因保守序列区域设计合成1对引物,用于鉴定伪狂犬病毒DNA,其引物序列如下为p1:5′-CACGGAGGACGAGCTGGGGCT-3′,p2:5′-GTCCACGCCCCGCTTGAAGCT-3′,预期扩增片段的长度为217 bp。
以PRV病毒DNA作为模板,PCR反应体系为:ExTap预混酶10 μL,上、下游引物各1 μL,模板DNA 1 μL,ddH2O 7 μL。反应程序为: 95 ℃ 5 min,94 ℃ 40 s,65 ℃ 30 s,72 ℃ 45 s,35个循环;72 ℃ 5 min;取PCR产物于2%琼脂糖凝胶上电泳,用凝胶成像系统拍照并记录结果。
1.2.3 gD蛋白抗原表位的扩增 依据GenBank中PRV 毒株(BJ/YT)gD蛋白的基因序列设计引物,引物由生工生物工程(上海)股份有限公司合成,引物序列为gD-1:5′-CCGGGATCCCCCCAGGTTCCCATACACTC-3′,gD-2:5′-CCCAAGCTTCTATTACGGCGTCAGGAATCGCATCAC-3′,预期扩增片段长度为702 bp。
以提取的PRV DNA为模板,用引物gD-1、gD-2进行PCR扩增。反应体系20 μL:LATaq酶0.2 μL,上、下游引物各0.5 μL,dNTP 1 μL,模板DNA 1 μL,2×GC Buffer 10 μL,ddH2O 6.8 μL 。反应条件: 95 ℃预变性5 min;94 ℃ 30 s,68 ℃ 30 s,72 ℃ 90 s,30个循环;72 ℃延伸10 min。取PCR产物进行琼脂糖凝胶电泳,用凝胶成像系统拍照记录结果并进行回收。
1.2.4 gD蛋白抗原表位的克隆 将回收的DNA片段与pMD19-T载体16 ℃连接过夜,将10 μL连接产物加入DH5α感受态细胞中,轻轻混匀,冰浴30 min,42 ℃热激90 s,迅速置冰上1~2 min,随后加入1 mL LB培养液,37 ℃、220 r/min振荡培养1 h。3 000 r/min离心5 min,弃上清,加入200 μL LB培养液悬浮细胞沉淀,涂布LB平板(含100 μg/mL Ampicillin、24 μg/mL IPTG、40 μg/mL X-gal),37 ℃过夜培养至蓝白菌落出现,挑取白色菌落进行PCR及双酶切鉴定pMD19-T-gD质粒。
1.2.5 gD蛋白抗原表位表达载体的构建 利用BamHⅠ、Hind Ⅲ双酶切pMD19-T-gD质粒和pET-28a表达载体,用连接酶连接回收的目的片段和表达载体。将连接产物转化Rosetta感受态细胞,挑取单个菌落进行PCR及双酶切鉴定pET-28a-gD质粒,并对其进行序列测定。
1.2.6 gD蛋白抗原表位的诱导表达 挑取阳性菌落过夜培养后,取100 μL菌液接种于100 mL LB培养液中(含100 μg/mL Ampicillin),37 ℃、230 r/min培养至OD600为0.6~1.0,加入终浓度为1.0 mmol/L的IPTG,37 ℃诱导表达8 h。收集1 mL菌液分装于1.5 mL的离心管中,12 000 r/min离心5 min,沉淀用100 μL PBS缓冲液重悬,加入25 μL 5×SDS上样Buffer,煮沸8 min。将未诱导的重组菌作为对照进行SDS-PAGE电泳。
1.2.7 gD蛋白抗原表位表达产物SDS-PAGE电泳 配制15%的分离胶,取上述诱导表达菌液200~300 μL于1.5 mL eppendorf 管中,8 000 r/min离心1~2 min,弃去上清,用20 μL PBS缓冲液重悬,加入5 μL 5×Buffer(用前加热)。放入浮漂中,煮沸8~10 min。12 000 r/min离心2 min,取上清上样。考马斯蓝染色,脱色后观察。
1.2.8 gD蛋白抗原表位Western blot检测 取硝酸纤维素膜(NC膜),依据所切下蛋白质胶的大小,小心裁剪。将膜、蛋白质胶、滤纸分别用甲醇、转膜缓冲液浸泡处理好之后按照滤纸、NC膜、SDS-PAGE胶、滤纸的顺序放入电转仪,15 V电转1 h。取出样品,SDS-PAGE胶用考马斯亮蓝继续染色,NC膜用5%的脱脂奶封闭过夜,依次加入一抗(His单抗、PRV阳性血清及阴性血清)、二抗(辣根过氧化物酶标记的羊抗鼠IgG抗体及羊抗猪IgG抗体)进行处理,并用AEC显色方法进行显色。
2 结果与分析
2.1 gD蛋白抗原表位的PCR扩增及克隆
以PRV病毒培养物的总DNA为模板,用引物p1、p2进行PCR扩增获得与预期大小一致的特异性目的条带,证明了模板DNA中PRV DNA的存在(图1),为目的序列的扩增奠定了基础。
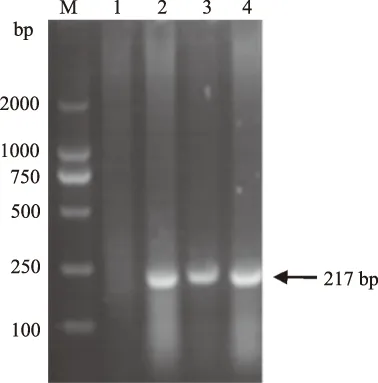
M.DNA Marker DL2000; 1.正常细胞DNA模板扩增产物; 2—4.病毒培养物DNA模板扩增产物图1 PRV DNA模板PCR扩增
以上述鉴定正确的DNA为模板,利用引物gD-1、gD-2进行扩增获得与预期大小一致的特异性目的条带(图2),成功扩增了gD蛋白的抗原表位。回收扩增产物并将其克隆于pMD19-T载体,成功构建了pMD19T-gD重组质粒。
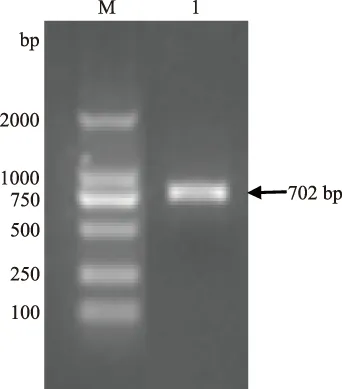
M.DNA Marker DL2000; 1.gD 抗原表位图2 gD抗原表位扩增结果
2.2 pET-28a-gD表达载体鉴定
2.2.1 酶切鉴定结果 培养重组工程菌,提取制备pET-28a-gD重组质粒,经限制性内切酶BamH Ⅰ、Hind Ⅲ酶切后,琼脂糖凝胶电泳可见702 bp的目的条带和5 900 bp的载体条带,与预期片段的大小一致(图3)。
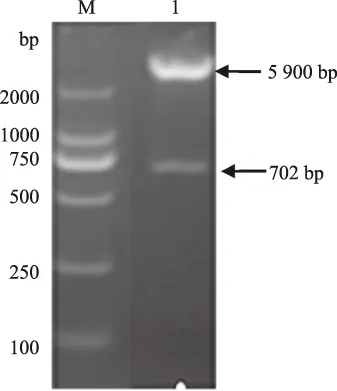
M.DNA Marker DL2000; 1.pET-28a-gD 双酶切产物图3 pET-28a-gD 表达载体双酶切鉴定
2.2.2 PCR鉴定结果 以pET-28a-gD表达载体质粒为模板,利用引物gD-1、gD-2进行PCR扩增, 扩增产物经琼脂糖电泳,结果显示扩增出了约702 bp大小的特异条带,与预期片段的大小一致(图4)。
以上结果表明,目的片段已插入表达载体。
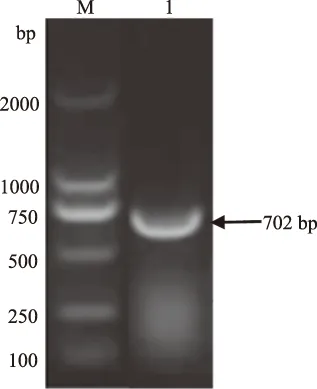
M.DNA Marker DL2000; 1.pET-28a-gD PCR 扩增产物图4 pET-28a-gD质粒PCR鉴定
2.3 gD蛋白抗原表位片段序列测定结果
对pET-28a-gD表达载体进行序列测定分析,结果显示,所插入的目片段的序列完整,在表达载体的阅读框内能够获得正确的阅读,其编码的氨基酸序列与天然gD抗原表位氨基酸序列一致。结果表明,成功的构建了pET-28a-gD表达载体。
2.4 gD蛋白抗原表位SDS-PAGE鉴定结果
重组阳性菌用IPTG于37 ℃诱导培养后,经SDS-PAGE电泳,结果显示,诱导重组菌比未诱导重组菌多出1条蛋白质条带,其分子质量约25 ku,与预期表达蛋白的大小一致(图5)。
2.5 gD蛋白抗原表位Western blot分析结果
gD蛋白抗原表位经SDS-PAGE电泳后,将凝胶上的蛋白质条带电转移到NC膜上,并进行Wes-tern blot分析。结果显示,在25 ku大小的位置呈现特异性反应条带(图6)。表明,表达产物能被伪狂犬阳性血清特异性识别,具有生物学活性。
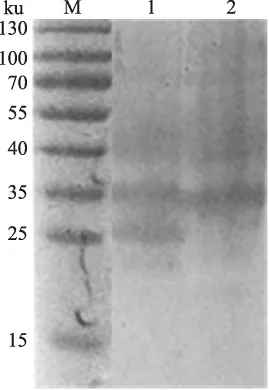
M.广谱彩虹预染蛋白Marker; 1.IPTG诱导菌; 2.IPTG未诱导菌图5 gD蛋白抗原表位SDS-PAGE 电泳
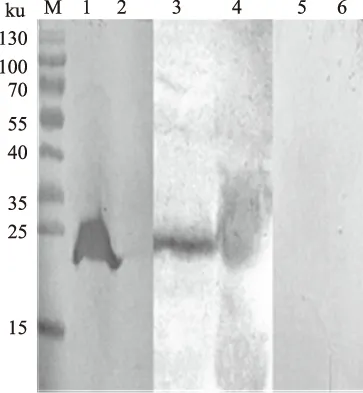
M.预染蛋白质Marker; 1.His单抗(诱导); 2.His单抗(未诱导); 3.PRV 阳性血清(诱导); 4.PRV 阳性血清(未诱导);5.PRV 阴性血清(诱导); 6.PRV 阴性血清(未诱导)图6 gD蛋白抗原表位Western blot分析结果
3 结论与讨论
PRV gD蛋白作为成熟病毒粒子囊膜表面主要糖蛋白,能诱导机体产生中和抗体,而这些抗体无论在有无补体存在的情况下,都具有中和PRV的能力[15],在机体保护方面发挥主要作用。表达gD糖蛋白,利用表达蛋白建立ELISA检测方法,检测免疫猪血清中的对应抗体效价,是评价疫苗的免疫效果的有效方法之一[16]。
由于PRV病毒基因组的GC含量较高,使得目的片段的扩增难度增加,常用的ExTaq酶扩增方法往往无法得到目的片段,使用LATaq酶及GC Buffer方可有效扩增[17]。前人研究表明,gD蛋白存在一定数量的线性抗原表位[18-19],这些抗原表位能够被不同的单克隆抗体识别,具有较强的抗原反应原性。因此,体外表达抗原表位蛋白用于疫苗免疫效果的评价,是一种行之有效的方法。此外,该方法也可用于缺失疫苗免疫猪群的鉴别诊断。在试验过程中发现PRV蛋白的表达有一定的难度,构建的表达载体往往不能够表达,其主要原因可能是PRV 蛋白与工程菌体的兼容性较小所致。因此,在研究PRV蛋白体外表达时最好采用真核表达系统。本研究采用原核表达系统成功表达了PRV gD抗原表位,为猪群PRV免疫效价的检测奠定了基础。
[1] 殷震,刘景华.动物病毒学[M].1版.北京:科学出版祉,1985:705.
[2] 樊振华,姚敬明,孟帆,等.山西部分种猪场猪伪狂犬病分子流行病学调研[J].山西农业科学,2012,40(9):989-992.
[3] 杨庆芳,宁官保,李俊达.猪伪狂犬病病毒的分离鉴定[J].山西农业科学,2011,39(8):886-889.
[4] 陆承平.兽医微生物学[M].3版.北京:中国农业出版社,2001:472.
[5] 余波,周思旋,谭诗文,等.猪伪狂犬病毒野毒株SYBR Green Ⅰ实时荧光定量PCR诊断试剂盒的研制[J].河南农业科学,2014,43(6):128-131,144.
[6] 高晓云,顾阳,潘鑫龙,等.猪伪狂犬病病毒河南分离株gE全基因的克隆与序列分析[J].华北农学报,2015,3(1):137-141.
[7] 顾阳,高晓云,程琨,等.鉴别猪伪狂犬病病毒强毒与疫苗毒双重PCR检测方法的建立[J].华北农学报,2014,29(2):94-97.
[8] 李柠摘.伪狂犬病病毒[J].国外兽医学——畜禽疾病,1995,16(3):29-32.
[9] 杜念兴.兽医免疫学[M].北京:中国农业出版社,1995.
[10] 殷震.动物病毒学[M].北京:科学出版社,1997:988-1009.
[11] 杨庆芳,宁关宝.猪伪狂犬病的研究进展[J].中国畜牧兽医,2010,7(9):15-18.
[12] 母安雄,谷根林,蒋建一,等.规模猪场主要疫病抗体水平监测及免疫效果分析[J].畜牧与兽医,2002,34(3):29-30.
[13] 白挨泉,王晓清,甄劲松,等.广东部分集约化猪场猪伪狂犬病毒野毒感染的血清学调查[J].中国畜牧兽医,2005,32(3):55-58.
[14] 李学伍,陈焕春,杨艳艳,等.PCR法对伪狂犬病病猪不同部位的检测及敏感性试验[J].中国畜禽传染病,1998,20(6):365-368.
[15] Hampl H,Ben-Porat T,Ehrlicher L,etal.Characterization of envelope proteins of pseudorabies virus[J].J Viro1,1984,52:583-590.
[16] 费流芳,伍时达,吴虹,等.猪瘟与伪狂犬病混合感染的防制[J].中国兽医科技,1995,33(4):33.
[17] 陈焕春,方六荣,何启盖,等.猪伪狂犬病病毒鄂A株的分离鉴定[J].畜牧兽医学报,1998,29(2):156-161.
[18] 范伟兴,魏荣,张雪莲,等.PRV LA 株gD基因的序列测定及其重复高变区的发现[J].中国兽医学报,2003,23(4):350-352.
[19] 陈焕春,周复春,力六荣,等.伪狂犬病病毒鄂A株TK-gG/LacZ突变株的构建[J].病毒学报,2001,17(1):69-74.
Cloning and Expression of Epitopes of gD Protein of Pseudorabies Virus
LIU Fang1,ZHANG Chong1,2,WANG Yinbiao2,ZHAO Pu2,DENG Ruiguang2,LI Xuewu2*
(1.College of Animal Science and Technology,Henan University of Science and Technology,Luoyang 471003,China;2.Henan Key Laboratory of Animal Immunology/Henan Academy of Agricultural Sciences,Zhengzhou 450002,China)
According to the pseudorabies virus strain BJ/YTgDgene sequences in GenBank,a pair of primers were designed .The B-cell epitopes of gD was amplified,and the length of PCR product was 702 bp.The amplified fragment was cloned into the cloning vector pMD19-T successfully named pMD19-T-gD.The fragment was digested byBamH I andHindIII restriction enzymes and then cloned into the expression vector pET-28a named pET-28a-gD.The expression of protein was induced with IPTG,specific protein bands appeared at the site of 25 ku by SDS-PAGE electrophoresis.Western blot analysis showed that the expression product could be identified by pseudorabies virus positive serum .In this study,gD protein epitope was cloned and expressed successfully,and the expression product had good biological activity.
pseudorabies virus;gDgene; epitope; protein expression
2015-11-20
国家自然科学基金项目(31172348)
刘 芳(1988-),女,河南洛阳人,在读硕士研究生,研究方向:畜禽疫病快速检测与防制。 E-mail:346103828@qq.com
*通讯作者:李学伍(1964-)男,河南通许人,研究员,博士,主要从事动物病毒生物学研究。E-mail:lixuewu2002@126.com
S855.3
A
1004-3268(2016)04-0122-04
