三个抗寒紫花苜蓿品系的寒境生理适应性研究
2016-02-05魏双霞师尚礼康文娟谭谌淼
魏双霞,师尚礼,康文娟,谭谌淼
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
三个抗寒紫花苜蓿品系的寒境生理适应性研究
魏双霞,师尚礼,康文娟,谭谌淼
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
【目的】 以‘俄罗斯杂花苜蓿’‘金皇后紫花苜蓿’和‘阿尔冈金紫花苜蓿’为对照材料,以暖温半干旱气候区(甘肃农业大学兰州牧草试验站)为对照区,对育成的3个抗寒紫花苜蓿品系‘GNKH-1’‘GNKH-2’‘GNKH-3’在高寒湿润气候区(甘南州夏河县)进行抗寒生理适应性研究.【方法】 采用同期采样法,在2个气候区的最后一茬即10月初采集长势一致的6个参试苜蓿材料,测定叶部和根部的脯氨酸、可溶性糖、可溶性蛋白质、丙二醛含量.【结果】 低温胁迫下,高寒区供试苜蓿越冬率显著低于对照区,其中,高寒区抗寒苜蓿品系越冬率显著高于对照材料,‘GNKH-3’和‘GNKH-2’的越冬率最高,分别为76.66%、72.34%;苜蓿叶部和根部的脯氨酸含量和可溶性糖均上升,‘GNKH-3’脯氨酸增加的程度最大,分别增加了783.15、881.04 μg/g,‘GNKH-2’可溶性糖增加较大,分别增加了6.64%、13.99%;高寒区‘GNKH-2’根部可溶性蛋白质含量最高,为2.84 mg/g;‘GNKH-1’‘GNKH-3’和‘阿尔冈金’叶部和根部MDA增加的幅度最小,且含量较低.【结论】 以抗寒生理指标为基础采用隶属函数法综合分析供试苜蓿材料在高寒湿润气候区的生理适应性,其顺序为:‘俄罗斯杂花’>‘GNKH-2’>‘GNKH-3’>‘GNKH-1’>‘金皇后’>‘阿尔冈金’.
苜蓿;新品系;越冬率;生理;抗寒性
苜蓿(MedicagosativaL.)是一种重要的豆科饲料作物,因其具有产草量高、富含蛋白质、适口性好、适应性强的特点而被誉为“牧草之王”,在世界范围内广泛种植.苜蓿是我国目前栽培面积最大的牧草,已达到190万hm2,居世界苜蓿栽培面积第6位[1-2].由于引种和管理利用方式不当引起的苜蓿越冬率低,甚至大面积死亡,造成了严重的经济损失.解决这一问题,需要了解紫花苜蓿对寒冷适应的规律和特点[3-4].因此,深入研究苜蓿的抗寒性,对于克服该地区寒冷等自然条件对苜蓿栽培的制约,扩大其种植范围,提高生产力,具有举足轻重的意义.
截止目前,我国先后从全世界5大洲37个国家引进苜蓿品种410多个,多数为紫花苜蓿[5-7].国内学者李兴福等[8-13]对于抗寒苜蓿引种做了一些研究,探讨了苜蓿在低温胁迫下的生理变化,并筛选出了一些抗寒苜蓿材料,比如‘俄罗斯杂花’‘金皇后’和‘阿尔冈金’等,郝凤等[14]于2010年成功克隆抗冻基因AFP和CBF2导入苜蓿基因组中,提高苜蓿抗寒性.本研究以3个抗寒苜蓿育成品系为供试材料,以抗寒性较强的‘俄罗斯杂花’‘金皇后’和‘阿尔冈金’为对照,以暖温半干旱气候区为参考区,从越冬率和生理角度探讨高寒湿润气候区低温胁迫对3个苜蓿育成品系的影响和抗寒性生理差异,为抗寒性苜蓿的选育提供理论依据和技术支持.
1 材料与方法
1.1 试验地概况
试验于2013年4月~2014年5月在高寒湿润气候区的甘南州夏河县牧草试验站和对照区(暖温半干旱气候区的甘肃农业大学兰州牧草试验站)进行.夏河县位于甘南藏族自治州西北部,地理坐标E 101°54′~103°25′,N 34°32′~35°34′,地处青藏高原东北边缘,地势由西北向东南倾斜,海拔3 000~3 800 m, 属于寒凉湿润性气候,高原大陆性气候比较明显,地势平坦,土壤类型为高寒草甸土,有明显的腐殖质积聚,腐殖质层厚10 cm,呈灰棕至黑褐色粒状-扁核状结构,土壤有机质含量45.14 g/kg,pH 7.87,速效氮50.83 mg/kg,速效磷10.57 mg/kg,速效钾230.0 mg/kg,吸附水3.08%,土壤肥力均匀.
对照区甘肃农业大学兰州牧草试验站位于兰州市西北部,地处黄土高原西端,E 105°41′,N 34°05′,海拔1 525 m,属温带半干旱大陆性气候,区内地势平坦,肥力均匀,土壤类型为黄绵土,黄土层较薄,土壤有机质含量0.84%,pH 7.5,土壤含盐量0.25%,有效氮95.05 mg/kg,有效磷7.32 mg/kg,有效钾182.8 mg/kg,土壤肥力均匀.
1.2 试验材料
供试材料为‘GNKH-1’(MedicagosativaL.kanghanNo.1)‘GNKH-2 ’(M.sativaL.kanghanNo.2)和‘GNKH-3’(M.sativaL.kanghanNo.3)为甘肃农业大学草业学院育成品系,具有较强的抗寒能力和优质高产性能.对照材料为抗寒能力强的苜蓿品种‘俄罗斯杂花苜蓿’(M.sativaL.subsp,varia)(秋眠级:2)‘金皇后紫花苜蓿’(M.sativaL.GoldenEmpress)(秋眠级:2)和‘阿尔冈金紫花苜蓿’(M.sativacv.Algonguin)(秋眠级:3)均由甘肃农业大学草业生态系统教育部重点实验室提供.
1.3 试验设计
试验采用随机区组设计,小区面积3 m×5 m,3次重复,人工开沟条播,播深2 cm,行距30 cm,播量1.7 g/m2.适时对试验地进行锄草及病虫害防治等田间管理.采用同期采样法,在2个气候区的最后一茬即10月初采集长势一致的6个参试苜蓿材料5株,3次重复.冲洗干净叶片和根部后,用滤纸吸干多余水分,封入密封袋并迅速放入液氮中速冻,贮存于-80 ℃超低温冰箱,用于生理指标的测定;第二年返青期进行越冬率测定.
1.4 测定指标及方法
越冬率用返青株数与前一年样段内总株数相比所得;游离脯氨酸含量采用酸性茚三酮法测定,可溶性糖含量采用蒽酮法测定,可溶性蛋白质含量采用考马斯亮蓝G-250 染色法测定,丙二醛含量采用硫代巴比妥酸法测定[15].
1.5 数据处理
用SPSS 17.0统计软件和EXCEL进行数据处理及分析,应用 Fuzzy数学中隶属度函数法[16]对多个指标进行综合评判.对与抗寒性呈正相关参数的越冬率、游离脯氨酸(Pro)、可溶性糖(WSS)、可溶性蛋白质(SP)采用公式Fij=(Xij-Xjmin)/(Xjmax-Xjmin),对与抗寒性呈负相关参数的丙二醛(MDA),采用公式Fij=1-(Xij-Xjmin)/(Xjmax-Xjmin).式中,Fij为i品种的j性状测定的具体隶属值;Xij为i品种j性状标准化值;Xjmin为i品种j性状中标准后的最小值;Xjmax为i品种j性状中标准化后的最大值;然后分别对两个气候区,每一材料所有性状的具体隶属值进行累加,求平均值得到该材料的隶属度值.
2 结果与分析
2.1 越冬率
由图1可知,低温胁迫均促使供试苜蓿材料的越冬率降低,高寒区显著低于对照区(P<0.05),高寒区平均越冬率为60.32%,对照区平均越冬率97.57%.高寒区供试材料越冬率排序为:‘GNKH-3’>‘GNKH-2’>‘GNKH-1’>‘金皇后’>‘俄罗斯杂花’>‘阿尔冈金’,‘GNKH-3’‘GNKH-2’‘GNKH-1’越冬率高于对照苜蓿材料,分别为76.66%、72.34%、65.23%,分别是对照苜蓿材料‘俄罗斯杂花’的1.64、1.55、1.40倍,‘金皇后’的1.27、1.19、1.08倍,‘阿尔冈金’的1.90、1.79、1.62倍.
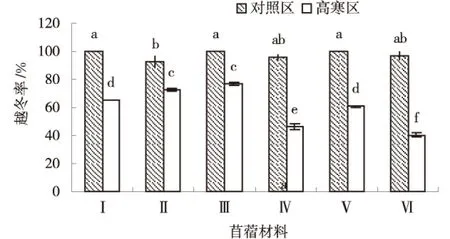
Ⅰ-GNKH-1,Ⅱ-GNKH-2,Ⅲ-GNKH-3,Ⅳ俄罗斯杂花苜蓿,Ⅴ-金皇后苜蓿,Ⅵ-阿尔冈金苜蓿,下同.图1 越冬率Fig.1 Wintering rate
2.2 抗寒生理指标
2.2.1 脯氨酸 由图2可知,除了GNKH-1,其他供试苜蓿材料的叶部游离脯氨酸含量均为高寒区显著高于对照区(P<0.05),高寒区各材料平均叶部脯氨酸含量为464.74 μg/g,对照区各材料平均叶部脯氨酸含量为1 007.02 μg/g.高寒区各材料叶部脯氨酸含量排序为:‘金皇后’>‘GNKH-3’>‘阿尔冈金’>‘俄罗斯杂花’>‘GNKH-2’>‘GNKH-1’‘金皇后’叶部脯氨酸含量最高,为1 271.44 μg/g,GNKH-1最低为714.58 μg/g.
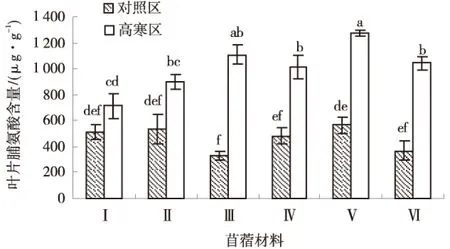
图2 叶片脯氨酸含量Fig.2 Content of leaf proline
表明经过低温锻炼,供试苜蓿材料叶部的游离脯氨酸含量均明显上升,‘GNKH-3’积累幅度最大,为783.15 μg/g,‘GNKH-3’比对照品种‘金皇后’‘阿尔冈金’和‘俄罗斯杂花’分别相对增加了75.31、109.78、255.61 μg/g,说明‘GNKH-3’在低温下叶部能够积累较多的游离脯氨酸来提高其抗寒性.
由图3可知,供试苜蓿材料根部游离脯氨酸含量均为高寒区高于对照区,且差异显著(P<0.05),高寒区材料平均根部脯氨酸含量为1 183.17 μg/g,对照区各材料平均根部脯氨酸含量为379.88 μg/g.高寒区各材料根部脯氨酸含量排序为:‘俄罗斯杂花’>‘金皇后’>‘阿尔冈金’>‘GNKH-1’>‘GNKH-3’>‘GNKH-2’,‘俄罗斯杂花’根部脯氨酸含量最高,为1 355.38 μg/g,显著高于抗寒苜蓿品系和‘阿尔冈金’,‘GNKH-2’根部脯氨酸含量最低为932.31 μg/g.
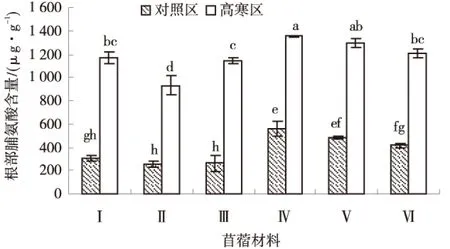
图3 根部脯氨酸含量Fig.3 Content of roots proline
表明经过低温锻炼,供试苜蓿材料根部的游离脯氨酸含量均明显上升,‘GNKH-3’积累幅度最大,为881.04 μg/g,‘GNKH-1’次之,为859.78 μg/g,‘GNKH-3’比对照品种‘金皇后’‘阿尔冈金’‘俄罗斯杂花’分别相对增加了67.43、87.72、88.17 μg/g,说明‘GNKH-3’和‘GNKH-1’在低温下根部也能够积累较多的游离脯氨酸来提高抗寒性.
2.2.2 可溶性糖 由图4可知,供试苜蓿材料叶部可溶性糖含量均为高寒区高于对照区,且差异显著(P<0.05),高寒区各材料平均叶部可溶性糖含量为11.63%,对照区各材料平均叶部可溶性糖含量为6.06%.高寒区各材料叶部可溶性糖含量排序为:‘GNKH-1’>‘阿尔冈金’>‘GNKH-2’>‘俄罗斯杂花’>‘GNKH-3’>‘金皇后’,‘GNKH-1’叶部可溶性糖含量最高,为13.30%,‘金皇后’最低为9.36%.
由供试苜蓿材料在高寒区和对照区叶部可溶性糖含量的变化情况可以看出,高寒区‘阿尔冈金’叶部可溶性糖积累最大,‘GNKH-2’次之,分别较对照区增加了7.94%和6.64%,高寒区‘阿尔冈金’比‘GNKH-2’相对增加了1.30%,‘GNKH-2’比对照品种‘俄罗斯杂花’和‘金皇后’分别相对增加了0.99%和3.68%,表明低温胁迫发生时,‘GNKH-2’和‘阿尔冈金’的叶片可溶性糖调节作用较强,有利于植株在低温环境中维持正常的水分生理作用.
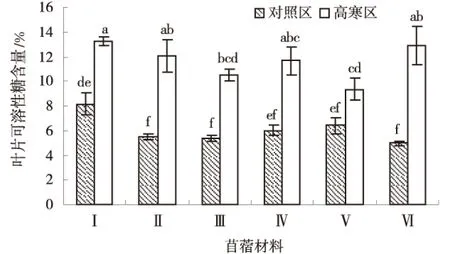
图4 叶片可溶性糖含量Fig.4 Content of leaf soluble sugar
由图5可知,除‘金皇后’紫花苜蓿外,其他供试苜蓿材料根部可溶性糖含量均为高寒区高于对照区,并且‘GNKH-1’和‘GNKH-2’根部可溶性糖含量高寒区显著高于对照区(P<0.05),高寒区各材料平均根部可溶性糖含量为23.90%,对照区各材料平均根部可溶性糖含量为20.50%.高寒区各材料根部可溶性糖含量排序为:‘GNKH-2’>‘俄罗斯杂花’>‘GNKH-3’>‘GNKH-1’>‘金皇后’>‘阿尔冈金’,‘GNKH-2’根部可溶性糖含量最高,为29.48%,显著高于其他供试苜蓿材料,‘阿尔冈金’最低为20.02%.
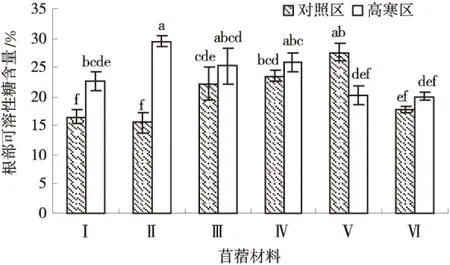
图5 根部可溶性糖含量Fig.5 Content of roots soluble sugar
由供试苜蓿材料在不同气候条件下根部可溶性糖含量的变化情况可以看出,经过低温锻炼,‘GNKH-2’积累幅度最大,‘GNKH-1’次之,‘GNKH-3’再次之,分别增加了13.99%、6.11%和3.10%;‘GNKH-2’比对照品种‘俄罗斯杂花’‘阿尔冈金’‘金皇后’分别相对增加了11.65%、 11.74%、 21.34%,表明低温胁迫发生时,抗寒苜蓿品系的根部可溶性糖调节作用较强,有利于植株在低温环境中维持正常的水分生理作用,其中‘GNKH-2’最强.
2.2.3 可溶性蛋白质 低温胁迫不利于可溶性蛋白质的合成.由图6可知,除‘俄罗斯杂花’外,其他供试苜蓿材料叶部可溶性蛋白质含量高寒区显著低于对照区(P<0.05),高寒区各材料平均叶部可溶性蛋白质含量为2.87 mg/g,对照区各材料平均叶部可溶性蛋白质含量为3.03 mg/g.高寒区各材料叶部可溶性蛋白质含量排序为:‘金皇后’>‘阿尔冈金’>‘俄罗斯杂花’>‘GNKH-3’>‘GNKH-1’>‘GNKH-2’,‘金皇后’叶部可溶性蛋白质含量最高,为2.99 mg/g,显著高于其他供试苜蓿材料,‘GNKH-2’叶部可溶性蛋白质含量最低为2.79 mg/g.
由供试苜蓿材料在不同气候条件下叶部可溶性蛋白质含量的变化情况可以看出,经过低温锻炼,‘GNKH-1’减少的幅度最大,‘GNKH-2’次之,‘GNKH-3’再次之,分别减少0.29、0.24和0.19 mg/g;‘俄罗斯杂花’减少幅度最小,‘金皇后’次之,分别减少了0.043和0.097 mg/g,‘俄罗斯杂花’比‘GNKH-3’‘GNKH-2’‘GNKH-1’相对增加了0.15、0.19、 0.25 mg/g.表明低温胁迫发生时,‘俄罗斯杂花’和‘金皇后’的叶部可溶性蛋白质调节作用较强,有利于植株在低温环境中维持正常的水分生理作用.
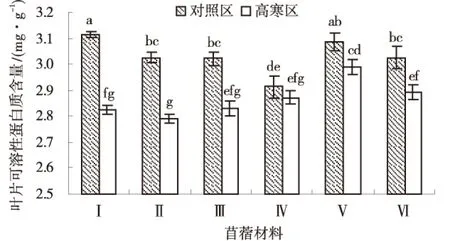
图6 叶片可溶性蛋白质含量Fig.6 Content of leaf soluble protein
由图7可知,供试苜蓿材料在高寒区和对照区根部可溶性蛋白质含量变化没有一定的规律,高寒区各材料平均根部可溶性蛋白质含量为2.71 mg/g,对照区各材料平均根部可溶性蛋白质含量为2.63 mg/g.高寒区各材料根部可溶性蛋白质含量排序为:‘GNKH-2’>‘俄罗斯杂花’>‘GNKH-3’>‘阿尔冈金’>‘GNKH-1’>‘金皇后’,‘GNKH-2’根部可溶性蛋白质含量最高,为2.84 mg/g,‘金皇后’最低为2.60 mg/g.
由供试苜蓿材料在不同气候条件下根部可溶性蛋白质含量的变化情况可以看出,经过低温锻炼,‘俄罗斯杂花’增加的幅度最大,‘阿尔冈金’次之,‘GNKH-3’再次之,分别增加了0.25、0.23、0.10 mg/g;‘金皇后’减少幅度最小,‘GNKH-1’次之,分别减少了0.032和0.085 mg/g,‘俄罗斯杂花’比‘GNKH-3’‘GNKH-2’‘GNKH-1’相对增加了0.15、0.24、 0.34 mg/g.表明低温胁迫发生时,‘俄罗斯杂花’和‘GNKH-2’的根部可溶性蛋白质调节作用较强,有利于植株在低温环境中维持正常的水分生理作用.
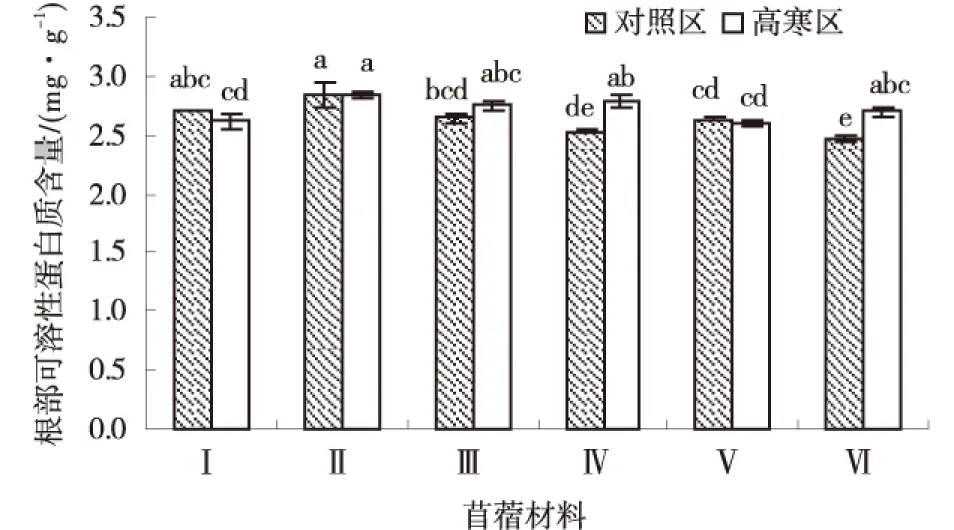
图7 根部可溶性蛋白质含量Fig.7 Content of roots soluble protein
2.2.4 丙二醛 由图8可知,‘GNKH-3’和‘阿尔冈金’叶部丙二醛含量高寒区显著低于对照区(P<0.05),高寒区各材料平均叶部丙二醛含量为14.15 μmol/g,对照区各材料平均叶部丙二醛含量为15.52 μmol/g.高寒区各材料叶部丙二醛含量排序为:‘金皇后’>‘GNKH-1’>‘俄罗斯杂花’>‘GNKH-2’>‘阿尔冈金’>‘GNKH-3’,‘金皇后’叶部丙二醛含量最高,为16.21 μmol/g,‘GNKH-3’最低为12.08 μmol/g.
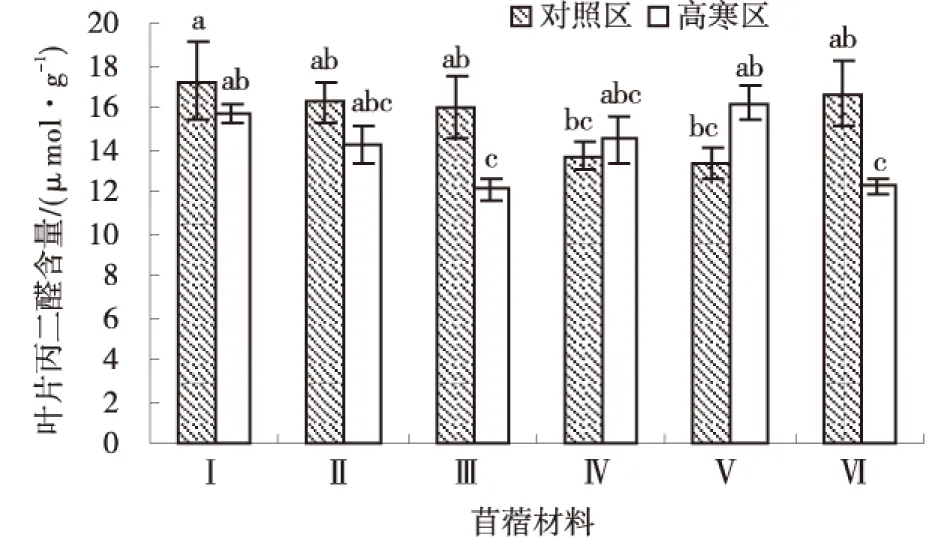
图8 叶片丙二醛含量Fig.8 Content of leaf MDA
由供试苜蓿材料在不同气候条件下叶部丙二醛含量的变化情况可以看出,经过低温锻炼,‘金皇后’和‘俄罗斯杂花’增加幅度较大,分别增加了2.88和0.80 μmol/g.抗寒苜蓿品系叶片丙二醛含量都降低;‘GNKH-3’和‘阿尔冈金’的变化幅度较小,分别减少了3.92和4.40 μmol/g,而且他们在低温条件下的含量较低,说明其膜脂过氧化程度较低,抗寒性较强.
由图9可知,‘GNKH-1’根部丙二醛含量高寒区显著低于对照区(P<0.05),高寒区各材料平均根部丙二醛含量为16.06 μmol/g,对照区各材料平均根部丙二醛含量为15.13 μmol/g.高寒区各材料根部丙二醛含量排序为:‘GNKH-3’>‘GNKH-2’>‘金皇后’>‘阿尔冈金’>‘GNKH-1’>‘俄罗斯杂花’,‘GNKH-3’根部丙二醛含量最高,为25.32 μmol/g,显著高于其他供试苜蓿材料,‘俄罗斯杂花’最低为11.79 μmol/g.
由供试苜蓿材料在不同气候条件下根部丙二醛含量的变化情况可以看出,经过低温锻炼,‘GNKH-3’和‘俄罗斯杂花’增加幅度较大,分别增加了6.50、3.58 μmol/g;‘GNKH-1’和‘阿尔冈金’的呈下降,分别减少了7.93和1.36 μmol/g,而且他们在低温条件下的含量较低,说明其膜脂过氧化程度较低,抗寒性较强.
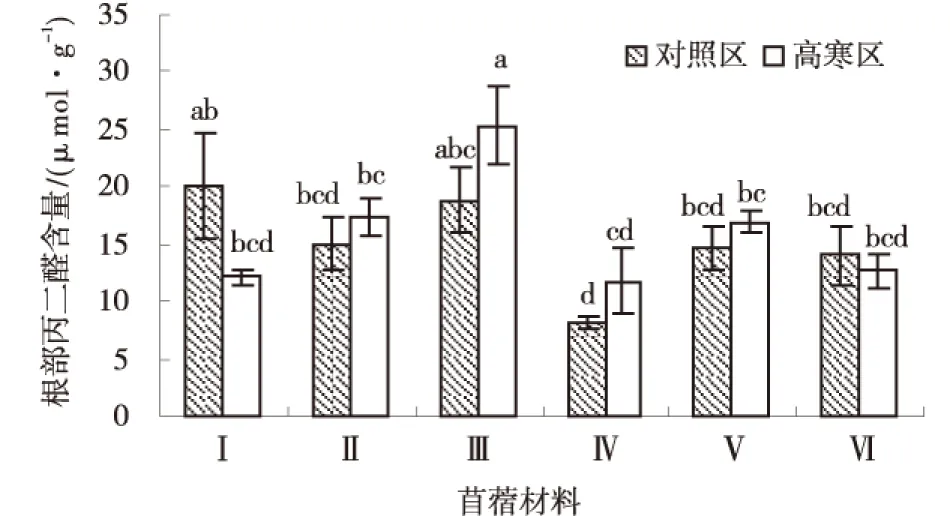
图9 根部丙二醛含量Fig.9 Content of roots MDA
2.3 高寒湿润气候区供试材料隶属度分析
植物受到逆境胁迫后,受到多种因素的影响,生理变化是错综复杂的[17],孤立的用某一个生理指标反映这个复杂的生理变化,很难较准确地反映植物的抗寒性本质[18],必须用多个指标进行隶属度分析.这里采用隶属函数法,项目包括越冬率;根部脯氨酸、可溶性糖、可溶性蛋白质、丙二醛.将供试材料各项生理指标的隶属函数值加起来求其平均值得其隶属度值.隶属度值越大,抗寒性越强,反之则弱.
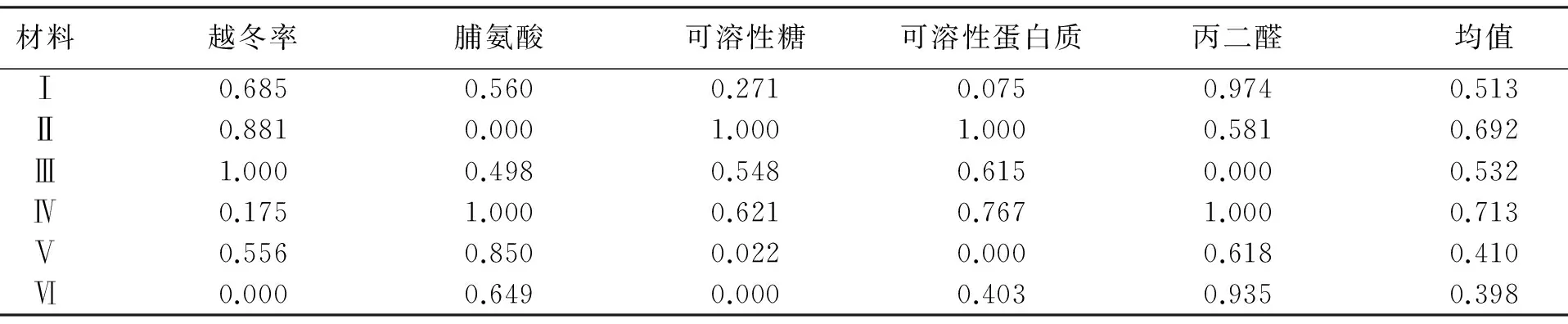
表1 在高寒湿润气候区供试苜蓿材料隶属度值Tab.1 Membership value of different alfalfa varieties in cold moist climate region
表1为高寒湿润气候区供试苜蓿材料5项抗寒参数的隶属度值,抗寒性强弱顺序为:‘俄罗斯杂花’(0.713)>‘GNKH-2’(0.692)>‘GNKH-3’(0.532)>‘GNKH-1’(0.513)>‘金皇后’(0.410)>‘阿尔冈金’(0.398).
对照区供试苜蓿材料5项抗寒参数的隶属度值分别为:‘金皇后’0.729、‘俄罗斯杂花’0.653、‘GNKH-3’0.434、‘GNKH-1’0.384、‘阿尔冈金’0.350、‘GNKH-2’0.286.
供试材料隶属度值高寒区均高于对照区,高寒区‘俄罗斯杂花’和抗寒苜蓿品系隶属度值均高于对照‘金皇后’和‘阿尔冈金’材料.
3 讨论
本研究从越冬率和脯氨酸含量、可溶性糖含量、可溶性蛋白质含量和丙二醛含量4个生理指标分析了苜蓿的抗寒性.结果表明,供试苜蓿材料越冬率高寒区显著低于对照区(P<0.05),高寒区‘GNKH-3’和‘GNKH-2’的越冬率排在所有供试材料的前两位,分别为76.66%、72.34%,显著高于‘GNKH-1’和对照材料;对照材料‘金皇后’和‘阿尔冈金’在高寒区的越冬率分别为60.58%、40.39%,低于丁培俊等[19]2001年所测定的‘金皇后紫花苜蓿’和‘阿尔冈金紫花苜蓿’在甘南的越冬率(74%、43%),造成的原因可能与当年的降水、田间管理措施不同等因素有关.越冬率是最重要的一个抗寒性指标,其大小可以反映品种的抗寒越冬能力大小.在低温胁迫下,植株的越冬率越高,表明抗寒性越强,越能适应高寒气候,因此,可以看出‘GNKH-3’和‘GNKH-2’抗寒性好.
苜蓿各项生理指标在生长期的低温反应下可导致产量和体内生理代谢发生变化.脯氨酸和可溶性糖作为重要的渗透调节物质,低温胁迫下迅速增加,能降低渗透势,维持膨压,是植物适应低温胁迫的一种重要机制[20].在植株处于低温胁迫状态时,其体内的游离脯氨酸具有一定的保护作用,它可以防止失水,增强蛋白质的水合作用,保护生物大分子结构和功能的稳定,能维持细胞结构、细胞运输和调节渗透压等,使植株表现出抗性.WSS是一种低温保护物质,可提高细胞液的浓度,增加细胞持水能力,从而降低细胞质的冰点,还可缓解细胞质过度脱水,保护细胞质胶体不致遇冷凝固,从而提高植物抗寒性[21].罗新义等[22]研究表明,低温胁迫下,‘肇东苜蓿’叶片中游离脯氨酸的含量逐渐累积,随低温处理时间的延长,含量逐渐增加.本研究结果与罗新义的研究结果一致,苜蓿叶片和根部脯氨酸和可溶性糖含量在低温胁迫下都有升高趋势,其中,‘GNKH-3’脯氨酸含量增加幅度最大,‘GNKH-2’可溶性糖含量增加幅度最大.可溶性蛋白质在低温下的浓度会增加,高浓度的氮可束缚自由水,提高抗寒性.低温胁迫导致苜蓿植株体内可溶性蛋白质增多,其中某些蛋白质既能起到酶的作用又有防冷冻的功能.Krasnuk等发现,低温胁迫导致苜蓿植株体内可溶性蛋白质增多,其中某些蛋白质既能起到酶的作用又有防冷冻的功能.本研究表明,在低温胁迫下,各个品种苜蓿叶片可溶性蛋白质都减少,‘GNKH-1’叶片可溶性蛋白质减少幅度最大,‘GNKH-2’次之,俄罗斯杂花减少幅度最小;‘GNKH-2’根部可溶性蛋白质积累最多.寒冷不利于苜蓿叶片积累可溶性蛋白质,但是根部可溶性蛋白质积累较多,表明‘GNKH-2’根部可以积累较多的可溶性蛋白质来提高其抗寒性.MDA是植物遭受低温胁迫时,生物膜系统膜脂过氧化的最终产物,它的积累对植物具有毒害作用,其浓度的高低代表膜脂过氧化程度及膜系统的伤害程度,直接关系到植物抗寒能力的强弱[23],与苜蓿抗寒性呈负相关,低温胁迫下,苜蓿体内丙二醛含量有升高的趋势.本试验表明,‘GNKH-3’‘GNKH-1’和‘阿尔冈金’在低温胁迫下的积累较少,而且他们在低温下的含量较低,表明,低温胁迫对他们的伤害最小.
本研究得出高寒湿润气候区供试苜蓿材料抗寒生理适应性最好的为‘俄罗斯杂花苜蓿’,然而‘GNKH-3’‘GNKH-2’抗寒生理适应性稍次于‘俄罗斯杂花苜蓿’,但实际越冬率比‘俄罗斯杂花’高,也反映出苜蓿在寒境环境中的生理适应性是个复杂的问题,这里研究的少量几个指标仅能反映该材料的部分抗寒潜在能力,没有完全反映出该材料的抗寒能力,故还需作进一步的研究探讨.
4 结论
1) 高寒湿润气候区供试苜蓿材料抗寒生理适应性强弱顺序为:‘俄罗斯杂花’>‘GNKH-2’>‘GNKH-3’>‘GNKH-1’>‘金皇后’>‘阿尔冈金’.
2) 高寒区供试苜蓿材料越冬率大小排序为:‘GNKH-3’>‘GNKH-2’>‘GNKH-1’>‘金皇后’>‘俄罗斯杂花’>‘阿尔冈金’.
3) 苜蓿在寒境环境中的生理适应性是个复杂的问题,少量几个生理指标仅能反映苜蓿材料的部分抗寒潜在能力,不能完全反映苜蓿材料的抗寒能力.
[1] 罗志成.北方旱地农业研究的进展与思考[J].干旱地区农业研究,1994,12(1):4-13
[2] 于林青.3种苜蓿形态特征及变异分析[J].草原与草坪,2008,128(3):29-33
[3] 邓雪柯,乔代蓉,李良,等.低温胁迫对紫花苜蓿生理特性影响的研究[J].四川大学学报:自然科学版,2005(1):190-194
[4] Brumme C E,Moore K J,Bjork C N.Agronomic consequences of dormant-non dormant alfalfa mixtures[J].Agronomy Journal,2002,94(4):782-789
[5] 桂枝,高建明.我国苜蓿育种的研究进展[J].天津农学院学报,2003,10(1):37-41
[6] 全国牧草品种审定委员会.中国牧草登记品种集[M].北京:中国农业大学出版社,1999
[7] 全国草品种的审定委员会.中国审定登记草品种集(1996-2000)[M].北京:中国农业出版社,2008
[8] 李兴福.紫花苜蓿引种品比试验[J].草业科学,2003,20(5):11-13
[9] 张耀生,赵新全,徐世晓,等.祁连山北坡河谷农业区苜蓿引种试验[J].草业科学,2004,21(2):22-24
[10] 石杰,高宇,阴玉华,等.高寒盐碱地区紫花苜蓿引种研究[J].安徽农业科学,2012,40(17):9274-9275
[11] 李发明,刘世增,郭春秀,等.民勤荒漠区优良苜蓿引种栽培试验研究[J].草业学报,2009,18(6):248-253
[12] 张瑞珍,张新跃,何光武,等.四川高寒牧区紫花苜蓿引进品种的筛选[J].草业科学,2006,23(4):43-46
[13] 孙广玉,李威,蔡敦江,等.高寒区苜蓿越冬的生理适应性[J].东北林业大学学报,2005,33(6):49-51
[14] 郝凤.AFP、CBF2基因表达载体的构建及其对苜蓿遗传转化的研究[D].兰州:甘肃农业大学,2010
[15] 邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000
[16] 龚明.作物抗旱性鉴定方法与指标及其综合评价[J].云南农业大学学报,1989,4(1):73-81
[17] 杨敏生,裴华,程志鹏.白杨杂种无性系抗寒性生理指标动态分析[J].生态学报,1997,21(4):367-375
[18] 沈漫,王明庥.黄敏仁.植物抗寒机理研究进展[J].植物学通报,1997,14(2):1-8
[19] 丁培俊,索燕萍,李冬红,等.甘南高寒牧区紫花苜蓿引种试验初报[J].中国草地学报,2006(5):113-117
[20] 王赞,李源,高洪文,等.鸭茅苗期抗旱性综合评价[J].干旱地区农业研究,2007,25(6):31-36
[21] Shonosuke sagisake,Yusuke Matsuda,Tohru Okuda,et al.Comparative studies of the changes in enzymatic activities in hardy and less hardy cultivars of winter wheat in late fall and in winter under snow soil[J].Sci Plant Nutr,1991,37(3):543-550
[22] 罗新义,冯昌军,李红.低温胁迫下肇东苜蓿SOD脯氨酸活性变化初报[J].中国草地,2004,26(4):79-81
[23] 赵世杰.植物组织中丙二醛测定方法的改进[J].植物生理学通讯,1994,30(3):207-210
(责任编辑 赵晓倩)
Physiological adaptability of three cold-resistant strains ofMedicagosativain cold zone
WEI Shuang-xia,SHI Shang-li,KANG Wen-juan,TAN Shen-miao
(College of Pratacultural Science,Key Laboratory of Grassland Ecosystem of Ministry of Education,Sino-U.S.Center for Grazing Land Ecosystem Sustainability,Gansu Agricultural University,Lanzhou 730070,China)
【Objective】 Physiological adaptation to coldness of three cold-resistant strains of Medicage sativa (‘GNKH-1’‘GNKH-2’‘GNKH-3’) was researched in cold humid climate zone (Xiahe County,Gannan Prefecture) by choosing ‘M.varia’‘M.sativacv.Conden Empress’ and ‘M.sativacv.Algonguin’ as controls and warm semi-arid climate zone (Gansu Agricultural University,Lanzhou forage experimental station)as control plot.【Method】 Collecting 6 kinds of alfalfa materials of the last stubble in early October in the two climate zone to determine the content of proline,soluble sugar,soluble protein and malondiadehyde(MDA) of leaves and roots.【Result】 The results showed that wintering rate of alfalfa in cold humid zone was significantly lower than that in control plot under low temperature stress ,cold-resistant strains had significant higher wintering rate compared with controls,‘GNKH-3’ and ‘GNKH-2’ reaching the highest (76.66% and 72.34%,respectively).The content of proline and soluble sugar in leaves and roots increased,‘GNKH-3’ increased in the maximal degree reaching 783.15 μg/g and 881.04 μg/g,respectively and the content of soluble sugar of ‘GNKH-2’ increased by 6.64% and 13.99%.The content of soluble protein of ‘GNKH-2’ in cold humid plot was the highest (2.84 mg/g).MDA content of ‘GNKH-1’‘GNKH-3’ and ‘M.sativacv.Algonguin’ increased in the minimum degree.【Conclusion】 Comprehensive priority of the physiological adaptability of tested ‘M.sativa’ materials in cold humid climate zone took in the order from high to low of ‘M.varia’‘GNKH-2’‘GNKH-3’‘GNKH-1’‘M.sativacv.Colden Empress’ and ‘M.sativacv.Algonguin’ using subordinate function analysis based on the cold-resistant physiological indexes.
alfalfa;new strains;wintering rate;physiology:cold resistance
魏双霞(1988-),女,硕士研究生,研究方向为草种质资源及育种.E-mail:2383298375@qq.com
师尚礼,男,教授,博士生导师,主要从事草种质资源及育种方面的教学与研究工作.E-mail:shishl@gsau.edu.cn
农业部公益性行业(农业)科研专项“青藏高原社区特色生态畜牧业关键技术集成与示范(甘南社区)”(201203010).
2015-11-09;
2016-11-25
S 541+.1
A
1003-4315(2016)06-0095-07
