超富营养水平下初始碱度对铜绿微囊藻生长影响的研究
2016-01-16王思莹张君枝马文林
王思莹,张君枝,马文林
(北京应对气候变化研究和人才培养基地,城市雨水与水环境教育部重点实验室,北京建筑大学 环境科学与工程系,北京 100044)
超富营养水平下初始碱度对铜绿微囊藻生长影响的研究
王思莹,张君枝,马文林
(北京应对气候变化研究和人才培养基地,城市雨水与水环境教育部重点实验室,北京建筑大学 环境科学与工程系,北京 100044)
摘要:以城市河湖天然水为试验用水,利用碳酸氢钠作为碱度调节剂,设置了不同碱度水平,研究了氮磷营养处于超富营养水平条件下,铜绿微囊藻各相关指标受碱度的影响。结果表明:碱度条件对藻类生长有显著影响,存在最适宜的碱度条件约为2.46mmol/L;水体中碱度的改变,会影响铜绿微囊藻对N、P的利用;碱度过高,藻类将吸收更多的营养物质来维持其生长。
关键词:碱度;铜绿微囊藻;生长
收稿日期:2014-12-18
作者简介:王思莹(1990—),女,北京人,北京建筑大学硕士研究生。
通讯作者:马文林(1968—),女,山西人,博士,教授,主要从事水环境生态修复技术、固体废弃物资源化处理与利用和适应气候变化等研究。
中图分类号:
文献标识码:A
文章编号::1674-9944(2015)02-0203-04
1引言
水体藻类生长离不开碳、氮、磷3种关键元素,而这3种元素也是引起水体富营养化的决定因素[1]。目前人们对水体富营养研究较多的是氮磷过量进入水体导致的影响[2],而对碳源的影响研究较少[3]。
在我国,每年都有大量有机物随着废水、雨水径流和垃圾等途径进入到地表水体中,它们中大部分在水微生物作用下被不断分解,向水中释放CO2。此外,大气中CO2浓度的升高,也导致从大气向水体转移的CO2量增加。水中CO2浓度的增加,促使水中碳酸盐平衡向碳酸盐增加的方向转变,引起水体发生无机碳源增强和碱度升高这样两个同步的水质效应。
藻类能够以水中无机碳作为其营养碳源。从营养源的角度看,碳酸盐浓度与藻类生长是正相关关系。但从碳酸盐离子强度对藻类细胞离子渗透压角度看,藻类生长有一个适宜的碳酸盐强度。也即,水中碳酸盐浓度增加对藻类生长的影响,是其作为碳源和碳酸盐离子强度两方面综合作用的结果。
微囊藻是一类全球性分布的蓝藻,在我国大部分富营养化水体中,铜绿微囊藻(Microcystisaeruginosa)在数量和发生频率上均占优势[4,5]。本论文以碱度为指标表征水中碳酸盐体系强度变化,利用小试实验研究初始碱度变化对铜绿微囊藻生长的影响,为揭示水中无机碳源对铜绿微囊藻生长影响进行理论基础。
2材料与方法
2.1 实验用水
取北京展览馆后湖湖水,回到实验室内,进行过滤、消毒处理,然后检测水中的碱度、总氮、总磷和pH值含量,结果为pH值8.2,TN 0.672mg/L,TP 0.0083mg/L,碱度1.12mmol/L。使实验体系的营养水平处于超富营养水平(TP、TN浓度分别≥0.2mg/L和≥2.0mg/L),通过适量添加K2HPO4和NaNO3来调节水体的总磷浓度和总氮浓度分别为2.85mg/L和0.19mg/L,然后储存备用。
2.2 实验方法
以湖水自然碱度(1.12mmol/L,T0组)为对照,通过向湖水中添加不同剂量的NaHCO3溶液,调整实验水的碱度分别为1.65mmol/L、2.46mmol/L、3.34mmol/L、4.01mmol/L、4.82mmol/L,简计为T1组、T2组、T3组、T4组和T5组,每个组各设2个平行样。为此,取500mL 广口锥形瓶12个,先向每个锥形瓶中各加入200mL经过预处理的天然湖水。留出1组2个锥形瓶作为对照组,不向其中添加NaHCO3溶液,其余5组10个锥形瓶作为处理组,分别向各组锥形瓶中添加不同量的NaHCO3溶液。将这些锥形瓶置于灭菌容器内121℃高温下灭菌20min,取出放凉备用。取处于对数生长期的铜绿微囊藻纯藻种,经离心、洗涤处理后,分别定量接入锥形瓶中,然后放入人工气候箱中进行培养。培养温度26℃,光照强度3000lux,光暗比12∶12。在光照周期内,每天摇动锥形瓶3~4次,促进营养循环,减少藻类下沉和器壁效应,并随机交换锥形瓶位置。
在实验过程中每天采集水样测定实验体系中的藻体生物量,每两天监测一次pH值、TN、TP和总碱度等浓度,得到在不同碱度下水质变化和藻类生长情况,并且对各指标的变化曲线进行分析,以研究碱度变化对藻类生长的影响规律。
2.3 监测指标与方法
本研究的测定项目包括pH值、碳酸盐碱度、重碳酸盐碱度、总碱度、总磷、总氮和浮游藻类生物量。其中,pH值采用酸度计测量;藻类生物量采用血球计数板方法测量;碳酸盐碱度、重碳酸盐碱度、总碱度采用自动电位滴定仪测量;总氮和总磷分别采用过硫酸钾氧化法—紫外分光光度法和钼锑抗分光光度法测量[6,7]。
2.4 藻类比增长速率计算
比增长速率采用公式(1)进行计算:
μ=ln(xn/xn-1)/(tn-tn-1)
(1)
式中:μ为比增长速率,xn、xn-1分别为第n天与第n-1天细胞计数值,tn、tn-1分别为对应于xn、xn-1的培养时间。
3结果与讨论
3.1 碱度对藻类生长的影响
3.1.1 藻类生长曲线
每日对6个碱度条件培养系统中铜绿微囊藻的藻密度值进行测定,绘制藻的生长曲线如图1所示。

图1 铜绿微囊藻的生长曲线(◆T0;■T1;▲T2;×T3;*T4;●T5)
从图1看出,各组培养体系中铜绿微囊藻的生长曲线有相似的变化趋势,都在第2天进入对数增长期,然后依次经历减速增长期和衰减期。所不同的是,各组藻类生长的速度不一样,培养体系中藻类生长从高向低的碱度顺序分别为2.46mmol/L、1.65mmol/L、3.34mmol/L、1.12mmol/L、4.01mmol/L、4.82mmol/L。2.46mmol/L碱度条件下藻的生长速度一直高于其他各组,在达到稳定期时具有最高的生物量;其次是1.65mmol/L组;3.34mmol/L组和1.12mmol/L组,无论是生长速度还是稳定期的最高生物量,都很接近;4.01mmol/L组和4.82mmol/L组藻的生长速度和最高藻密度都比较低,而且衰亡时间也相对提前,4.82mmol/L组的最低。
因此,2.46mmol/L的碱度条件,最适合于铜绿微囊藻生长,高于或低于这个浓度,藻类的生长情况都趋于下降,且总的来说,降低碱度比提高碱度更适合藻类生长。
杨威[9]等人以不含Na2CO3的BG11培养基为基础,,设计4组NaHCO3碱度水平培养组,培养条件与本实验基本相同,探讨碱度水平对铜绿微囊藻生长的影响。研究表明,铜绿微囊藻在不同碳酸氢盐处理中的总体生长趋势顺序是2.4 mmol/L>4.8mmol/L>0.2mmol/L>7.3mmol/L,在实验条件下最适宜的碳酸氢盐碱度水平约为2.4mmol/L。与本研究得出的碱度条件影响规律相一致。
3.1.2 最大比增长率
最大比增长率是一定条件下藻类潜在增长率的最高表现[10]。根据比增长率计算公式对各种碱度条件下的铜绿微囊藻的比增长率进行连续计算,分别求出对数生长期内的最大比增长率μmax以及在对数生长期和稳定期的平均比增长率μave,列于表1中。

表1 不同碱度对藻类生长速率的影响
从表1的数据看出,在单独培养过程中,6组藻所达到的最大比增长率,T2组的最高,T5组的最低,说明适量加入碳酸氢盐对藻类的生长有促进作用,但添加过量时会表现出抑制作用来。按照最大比增长率排列顺序依次为T2> T1> T3>T0> T4> T5组,这与稳定期时各组最高藻细胞密度是相一致的。
3.2 pH值和碱度的变化
实验过程中,对各培养体系的pH值、总碱度、重碳酸盐碱度以及碳酸盐碱度进行测定,这些指标随时间的变化情况如图2~图5所示。
图2表示的是实验过程中各个培养体系的pH值随时间的变化情况。实验初期,各组培养体系,由于添加NaHCO3的量不一样,表现出不同的pH值,添加NaHCO3越多的体系,其pH值越高。而随着实验的进行,最后各组pH值趋于接近的水平。

图2 pH随时间的变化(◆T0;■T1;▲T2;×T3;*T4;●T5)
溶解于水体中的无机碳共有4种存在形式:CO2、H2CO3、HCO3-和CO32-,它们之间存在着一定的动态平衡转化关系,其平衡方程为:CO2+H2O⟺H2CO3⟺H++HCO3-⟺2H++CO32-。现今越来越多的研究者发现CO2其实并不是水生植物获取无机碳的唯一途径,HCO3-同样能够作为藻类的重要无机碳源已经得到普遍认可[11,12]。影响实验体系pH值变化的因素包含2个作用相反的方面,分别是藻类光合作用吸收利用无机碳源(CO2和HCO-3)和藻类呼吸作用释放CO2。藻类生长过程中,利用水体中游离态CO2或HCO-3进行光合作用,平衡向左移动,H+浓度减少,故pH值增大[13]。而藻类呼吸过程释放二氧化碳,平衡将向右移动,pH值将下降。图2中不同实验体系的pH值变化情况,是这两个过程的综合作用结果。pH值升高的阶段,表明这个时期藻类生长旺盛,生长过程吸收利用二氧化碳和碳酸盐对水中pH值的影响作用强于呼吸过程释放二氧化碳的影响;pH值下降的阶段则是相反的作用机制。对比图2中pH值变化曲线和图1生长曲线,二者之间有较好的相关性。T2组生长量最高,实验结束时该组的pH值也最高。
图3~图5表示了各实验体系中总碱度、碳酸盐碱度和碳酸氢盐碱度的变化情况,总碱度和碳酸氢盐碱度有一致的变化趋势,与碳酸盐碱度变化趋势相反,说明实验体系的总碱度主要受碳酸氢盐碱度的控制。
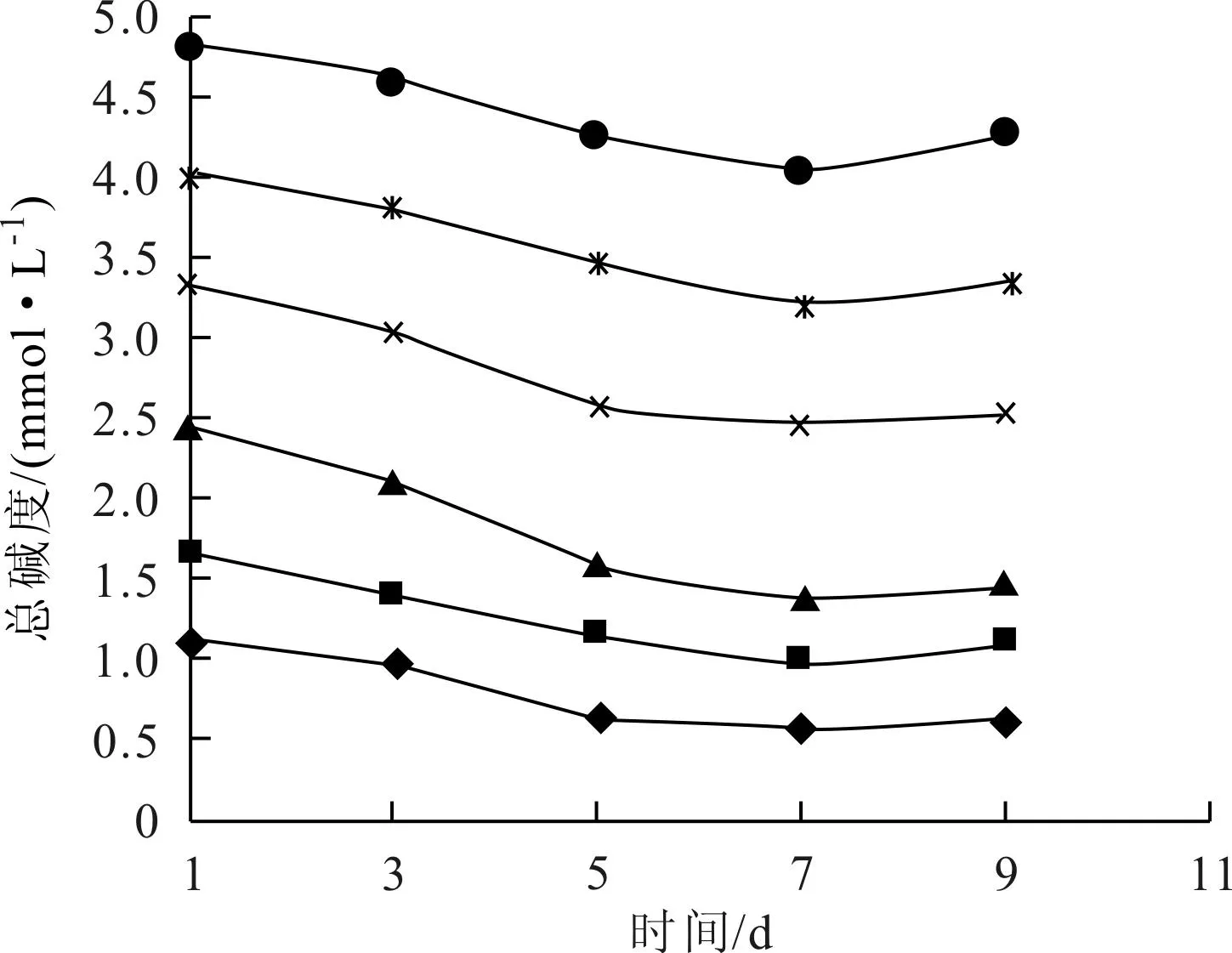
图3 总碱度随时间的变化(◆T0;■T1;▲T2;×T3;*T4;●T5)

图4 碳酸盐碱度随时间的变化(◆T0;■T1;▲T2;×T3;*T4;●T5)

图5 碳酸氢盐碱度随时间的变化(◆T0;■T1;▲T2;×T3;*T4;●T5)
藻类生长不能直接吸收碳酸盐,只能利用水体中游离态CO2或HCO-3进行光合作用[13]。Stumm和Morgan[14]指出,淡水水体的pH值在8 左右时,碳酸氢盐—碳占水体中总无机碳的90%左右;而pH值为9 左右时,游离CO2浓度接近于零。故在弱碱性条件下(pH值8~9) ,总无机碳中碳酸氢盐是主要的存在形态,也是藻类能够间接或直接利用的主要碳源形态[9]。在培养初期和中期,各体系中藻的生长量大,从水中大量吸收CO2和HCO-3,使得体系总碱度和碳酸氢盐碱度都降低,并因此使得pH值升高,碳酸盐平衡体系向右移动,水中碳酸盐浓度升高。实验后期,藻类开始先后出现衰亡迹象,生长和呼吸作用都减弱,对碳酸氢盐的利用减少,同时在一定的pH值条件下,水中碳酸盐体系达成新的平衡。从各图碱度曲线看,第5天和第7天的碱度变化比较大,对应于图1中,第3~7d铜绿微囊藻生长迅速,表明藻类生长对体系碱度变化确实存在显著影响。
3.3 TN和TP的变化情况
罗固源等人[15]研究表明:在TN/TP为12∶1时藻类的产生周期最短,产生量最多。为使实验用水的初始氮磷营养水平达到中富营养化程度,又能保证铜绿微囊藻的正常生长,故调节初始氮磷营养水平分别约为2.85mg/L和0.19mg/L,总氮与总磷的浓度比大约为15∶1。实验过程中水的TN和TP变化情况如图6和图7所示。

图6 TN随时间的变化及7天内TN的利用率(◆T0;■T1;▲T2;×T3;*T4;●T5;+利用率)

图7 TP随时间的变化及7天内TP的利用率(◆T0;■T1;▲T2;×T3;*T4;●T5;+利用率)
图6表明,随实验的进行,各体系TN浓度普遍呈下降趋势,在第7天趋于平衡。但不同碱度下,实验结束时各组的TN浓度稍有差异。T2组的TN浓度一直是6个组别中最低的,表明T2组具有最大的TN利用效率。T5组的TN浓度在实验初期处于6个组别的中等水平。
由图7可以看出,TP的变化趋势与TN的基本相同,但各组间的差异比TN的要显著。铜绿微囊藻的生长主要吸收利用水中的可溶性磷PO43-,但是由于碱度的影响,使得各组水藻的长势不同,所以对可溶性磷的利用率也不同的。实验结束时,各组实验体系中TP浓度由高到低的顺序依次为:T0、T1、T5、T4、T3和T2,相应地各组对磷的利用率分别为:38.1%、43.4%、51.3%、46.3%、46.3%、44.7%,该顺序与藻类生长曲线稍有不同,T4组和 T5组的TP的利用率相对较高,分析原因可能是过高的碱度环境,使得铜绿微囊藻在生长时消耗更多的可溶性磷。从曲线变化趋势可以看出,TP浓度在第3~7d下降程度较大,与藻类的生长趋势相对应。
比较藻对氮和磷的利用情况表明,铜绿微囊藻对TN的利用率比对TP的低。郑朔方[16]等人研究表明,铜绿微囊藻对总磷的半饱和常数KSP=0.019,对总氮的半饱和常数KSN=0.111,说明铜绿微囊藻对总磷的吸收能力远高于对总氮的吸收能力,这是该水藻能够高效利用磷的原因所在。
4结论
(1)铜绿微囊藻在含不同碱度水平的培养液中生长,能够间接或直接利用水中碳酸氢盐作为光合无机碳源。藻类的生长和碱度水平存在某些相关性。
(2)初始碱度的不同会对藻类的生长产生影响,本实验体系中最适宜的碱度条件约为2.46mmol/L,高于或低于这个浓度,藻类的生长情况都趋于下降,但总的来说,降低碱度比提高碱度更适合藻类生长。
(3)藻类生长会影响水体的酸碱水平,随着藻类的生长,pH值逐渐升高,而水中碱度的成分也会因此相互转换。
(4)水体中碱度的改变,会影响铜绿微囊藻对N、P的利用。各组实验体系中对氮的利用率分别为14.3%、14.7%、15.6%、14.9%、15.3%、15.1%;对磷的利用率分别为:38.1%、43.4%、51.3%、46.3%、46.3%、44.7%。碱度过高,藻类将吸收更多的营养物质来维持其生长,N、P利用量也会相应提高。
参考文献:
[1]郑焕春,周 青.微生物在富营养化水体生物修复中的作用[J].中国生态农业学报,2009,17(1):197~202.
[2]马经安,李红清.浅谈国内外江河湖库水体富营养化状况[J].长江流域资源与环境,2002,11(6):575~577.
[3]岳国峰,王金霞,朱明远,等. 藻类无机碳营养的研究进展(Ⅰ)——研究起源及研究方法[J].研究综述,2003,27(5):15~18.
[4]金相灿. 中国湖泊环境[M].北京:海洋出版社,1995.
[5]Lu Yuan, Wen Jianfan. Isolation,Pure Cultivation and Total DNA Extraction of Microcystis aeruginosa. Kütz in Dianchi Lake[J].Journal of Sciences, 2001,13(3):285~288.
[6]国家环境保护总局,水和废水监测分析方法编委会.水和废水监测分析方法[M].4版.北京:中国环境科学出版社,2002: 120~124.
[7]全国主要湖泊、水库富营养化研究课题组.湖泊富营养化调查规范[M].北京:中国环境科学出版社,1987.
[8]金相灿,章宗涉.湖泊富营养化调查规范[M].北京:中国环境科学出版社,1909:257~268.
[9]杨威,孙凌,袁有才.碱度水平对铜绿微囊藻和四尾栅藻生长和竞争的影响[J].农业环境科学学报,2007,26(4):1264~1268.
[10]方涛,敖鸿毅,刘剑彤.滇池水体理化环境状况时空分布格局研究[J].水生生物学报,2004,28(2):124~130.
[11]岳国峰,王金霞,朱明远.藻类无机碳营养的研究进展(Ⅱ)——藻类利用无机碳的机理及其调节[J].海洋科学,2003,27(6),31~34.
[12]Jones J I,Young J,Eaton J,et al.The influence of nutrient loading, dissolved in organic carbon and higher trophic levels on the interaction between submerged plants and periphyton[J].Journal of Ecology, 2002,90(1):12~24.
[13]孙凌,阚元卿,张冬梅.无机碳对浮游藻类生长和群落结构的影响[J].环境污染与防治,2007,29(5):352~356.
[14]Stumm W, Morgan J J .水化学—天然水体化学平衡导论[M].汤鸿霄,译.北京: 科学出版社, 1987:91~170.
[15]罗固源,康康,朱亮.水体中TN/TP与藻类产生周期及产生量的关系[J].重庆大学学报:自然科学版,2007,30(1):142~145.
[16]郑朔方,杨苏文,金相灿.铜绿微囊藻生长的营养动力学[J].环境科学,2005,26(2):152~156.
