川西北岷江冷杉种群的空间格局
2016-01-15康瑶瑶
川西北岷江冷杉种群的空间格局
康瑶瑶
(北京市林业勘察设计院,北京 100029)
摘要:以四川王朗国家自然保护区岷江冷杉林1hm`2样地的调查数据为基础,研究岷江冷杉的径级结构和重要值,运用Ripley’s K函数分析其空间格局和种内不同生长阶段的空间关联。结果表明:岷江冷杉在群落中占据绝对优势,中大径级林木较多。不同生长阶段的岷江冷杉主要呈聚集分布,可能与物种种子扩散方式有关;幼树和小树在所有空间尺度上均呈聚集分布,聚集强度随着尺度增大呈增大后减小的趋势,大树的空间格局随尺度增大,由随机(1~3m)趋于聚集分布(4~50m)。随着种群发育(幼树→小树→大树),岷江冷杉的聚集强度逐渐减小。幼树与小树在中小尺度上(1~25m)呈正向关联,不同生长阶段的岷江冷杉均与枯立木呈现出正向关联,随尺度变化而变化;岷江冷杉的聚集分布和种内的正向关联关系体现出了种群的生物学特性(耐荫)、干扰机制(林窗更新)和物种内部相互有利的生态关系。根据岷江冷杉的空间属性和更新机制,可以适当的进行人为干扰,以促进岷江冷杉林的发展。
关键词:岷江冷杉;空间格局;空间关联;点格局分析
中图分类号:S718.54
文献标志码:A
文章编号:2095-1914(2015)02-0030-07
Abstract:A studied diameter distribution and importance value of Abies faxoniana was made based on the field survey in a plot with an area of a 1 hm`2 of A. faxoniana forest in Wanglang Natural Reserve, Sichuan Province.In the study Ripley’s K function was used to analyze spatial pattern and spatial association of A. faxoniana at different growth stages. The result showed that: A. faxoniana was dominant species in the community with highest importance value, and there were many medium and big trees in the plot. A. faxoniana at different growth stages mainly showed aggregated distribution, indicating the way of seedling dispersal. Saplings and small trees were aggregated distributed at all scales, and aggregation intensity tended to increase first and then decreased as scale increased. While big trees tended to be clumped as scale increased.As age increase (sapling→small tree→big tree), aggregation intensity tended to reduce. Saplings and small trees showed positive association, while A. faxoniana at all growth stages were positively associated with standing dead trees, varied with spatial scale. The spatial pattern and association of A. faxoniana indicated the species′ shade-tolerance characteristic, positive relationship among age classes, and mechanism of gap disturbance affecting regeneration. Based on its spatial character and regeneration mechanism,it is suggested that some measures can be taken to facilitate the development of A. faxoniana.
Keywords:Abies faxoniana; spatial pattern; spatial association; point pattern analysis
收稿日期:2014-11-02
基金项目:广州市林业和园林局项目资助。
doi:10.11929/j.issn.2095-1914.2015.02.006
Spatial Distribution Pattern ofAbiesfaxonianain
Northwestern Sichuan
KANG Yao-yao
(Beijing Forestry Survey and Design Institute, Beijing 100029,China)
种群结构是种群的重要属性,不仅反应了种群个体在空间上的组配方式,也在一定程度上反应了种群的发展趋势[1-2]。空间分布格局是种内/种间相互关系及环境条件综合作用的结果[3-4],空间格局分析是研究种群结构特征、种群互作以及种群与环境间关系的重要研究方法,也是近年来生态学研究的热点之一[5-7]。由于空间格局与种群结构(特别是年龄结构)有着密切联系,因此,可以通过研究种群结构和空间格局来阐明森林生态系统的形成与维持、种群的生态特征以及群落的稳定性与演替规律,且在理论和实践中都具有重要意义[8-9]。
岷江冷杉(Abiesfaxoniana)为我国特有树种,耐阴性强,喜冷湿气候。岷江冷杉林是我国川西北亚高山地区的重要森林类型,主要分布于青藏高原东南缘以及四川盆地西缘山地[10],对于涵养水源、调控径流、减少水土流失等起着重要作用,是该地区重要的生态屏障[11-12]。目前,该地区已开展了岷江冷杉林小气候特征[13-14],林线种群特征[12,15]的研究,但关于优势种空间格局及空间关联的研究鲜见报道。本文分析了岷江冷杉林中优势树种岷江冷杉的空间格局及空间关联,探讨种群空间格局的发展动态和形成机制,以期为川西北亚高山森林生态系统的保育、恢复与可持续经营提供理论依据。
1研究区概况
研究区位于四川省平武县境内的王朗国家级自然保护区,地处东经103°55′~104°10′,北纬32°49′~33°02′,区内山高谷深,海拔2300~5000m,坡度多在30°以上。本区属于丹巴-松潘半湿润气候区,干湿季明显[13]。夏秋季暖湿,降水量大,日照少;冬春季日照强烈,降水少,空气干燥。年积温1056.5 ℃,年平均气温2.9 ℃,年降水量859.9mm。土壤以暗棕壤、棕壤和山地草甸土为主。山地植被垂直变化明显[16],由下至上为针阔混交林(2300~2600m)、方枝圆柏(Sabinasaltuaria)-紫果云杉(Piceapurpurea)林和岷江冷杉林(2600~3500m)、亚高山灌丛草甸(3500~4400m)和高山流石滩植被(4400~4900m)。
海拔2800~3200m的岷江冷杉林保存完好,乔木层的优势种为岷江冷杉,伴生有紫果云杉、方枝圆柏、槭树(Acerspp.)、红桦(Betulaalbosinensis)和糙皮桦(Betulautilis);灌木层主要有缺苞箭竹(Fargesiadenudata);草本层物种主要有紫花碎米芥(Cardaminetangutorum)、鹿药(Smilacinajaponica)和蕨类(Pteridophyta);苔藓层盖度大,主要为塔藓(Hylocomiumsplenden)和山羽藓(Abietinellaabietina)[14,16]。
2研究方法
2.1野外调查
研究样地设置在大窝凼,坡向东偏南32°,坡度为21°,海拔3020m。2009年8月,在林分保存完好的原始林内建立1 hm2(100m×100m)固定样地,为便于野外调查,将样地划分为100个10m×10m的小样方,记录每个小样方内所有胸径≥1cm林木的种类、胸径(cm)、树高(m)、冠幅(m2)和生长状态(枯立),并量取每个个体的坐标(x,y)来确定其空间位置。
2.2生长阶段划分与重要值计算
以往研究表明,采用胸径划分岷江冷杉个体的大小,具有良好的一致性[17]。为了研究不同径级个体的空间格局及相互间的空间关联,根据以往岷江冷杉生长阶段划分的经验(不同大小的个体在群落中所处的高度层次)[18-19],以胸径为依据将岷江冷杉种群划分为幼树(1~5cm)、小树(5~15cm)和大树(≥15cm)3个生长阶段。
树种在群落中的优势度通过重要值来判断,计算公式:
重要值=(相对多度+相对显著度+相对频度)/3
2.3空间格局分析
运用Ripley’sK点格局法进行空间格局分析。点格局分析法将每个个体视为二维空间的1个点,以全部个体组成的二维点图为基础进行格局分析[20]。近年来,Ripley’sK函数已经成为一种最流行的点格局分析方法[5,9,18,21],该函数通过描述不同空间尺度下种群的分布格局,更好地反映了种群的结构特点。Ripley’sK的基本公式为:
式中:t为距离(某一空间尺度);n为植物个体数;uij为点i和点j之间的距离,当uij≤t时,It(uij)=1,当uij>t时,It(uij)=0;Wij为以点i为圆心uij为半径的圆周长在面积A中的比例。

应用Ripley’sK12(t) 函数进行双变量空间关系分析,其计算及原理与Ripley’sK类似,该函数修正形式为:
当L12(t)=0时2个变量相互独立,当L12(t)>0时2个变量空间正关联,当L12(t)<0时2个变量负相关。通过Monte Carlo方法模拟了99%置信区间。本文Ripley’sK点格局分析通过ADE-4软件实现[9,23]。
3结果与分析
3.1乔木层的主要树种
由重要值的分析可知(表1),乔木层中岷江冷杉占据绝对优势,相对显著度、相对多度和相对频度值均为最大。伴生种主要有紫果云杉、方枝圆柏和槭树,较高的出现频率(相对频度)是其在群落中占有一席之地的重要原因。另外,紫果云杉的胸高断面积(相对显著度)和数量(相对多度)比其他2个伴生种高。

表1 乔木层4个主要树种的重要值
3.2岷江冷杉径级分布与空间分布图
野外调查中共测得岷江冷杉522株。其中幼树49株,小树51株,大树370株,枯立木52株;中小径级的个体(5~15cm)较为稀少,只占整个活立木的10.8%;中大径级的个体较多,15~50cm的个体占66.8%;枯立木的数量占整个种群的10%,其中小于20cm的枯立木占到了所有枯立木的57.7%,结果见图1。
点分布图可以直观的反映出个体在样地空间上的分布状况。岷江冷杉各生长阶段的密度差异较大,幼树和小树均呈现出团块状分布,而枯立木和大树的这种团块分布则相对较弱,受到空间尺度的影响(图2)。
3.3空间格局
由Ripley’sK点格局分析可知(图3),岷江冷杉整个种群的空间格局随尺度增加由聚集趋于随机分布,在1~47m的尺度上呈聚集分布,48~50m的尺度上呈随机分布;幼树和小树在所有空间尺度下均呈现出聚集分布,且聚集强度(L(t))随着尺度的增大呈现为先增大后减小,最大值分别出现在14m和30m处;大树在1~3m的尺度上呈随机分布,4~50m的尺度上呈聚集分布。不同生长阶段个体的聚集强度排序为:幼树>小树>大树(图3)。


3.4空间关联
不同生长阶段岷江冷杉个体的空间关联见图4,幼树和小树在1~25m的尺度上呈正相关,其余尺度下相互独立,说明幼树和小树在中小尺度下表现出互利关系;幼树和大树,小树和大树均在所有尺度下表现为空间独立。
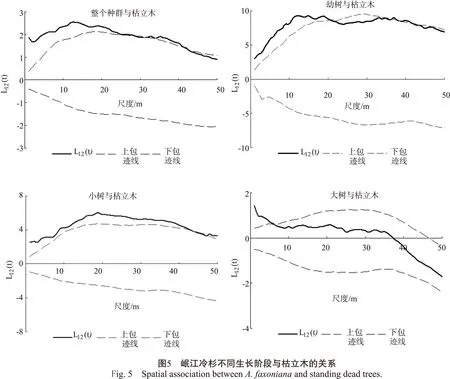
不同生长阶段的岷江冷杉均与枯立木表现出一定程度的正向关联,未出现负关联关系(图5),说明枯立木有利于岷江冷杉种群的发展。但这种正向关联的关系受空间尺度影响(图5),整个种群与枯立木在1~28m和32~42m尺度上表现为正相关;幼树与枯立木在1~20m、35~37m和40~41m尺度上呈正相关;小树在大部分尺度下(1~43m)与枯立木表现为正向关联;大树则与枯立木在1~6m尺度上呈正相关。

4结论与讨论
岷江冷杉的空间格局主要表现为聚集分布,该空间分布格局反映出岷江冷杉的更新方式和生物学特性,与前人研究结果一致[17-19]。幼树和小树在所有研究尺度下均呈聚集分布,且幼树的聚集强度最强,可能与种子的散布方式有关。岷江冷杉的种子多散布在母树周围[17,20],次年萌发后呈密集团状,另外岷江冷杉耐阴能力强,这种聚集分布状态可持续到幼树和小树阶段,且在幼树阶段表现出最大的聚集强度。幼年阶段的聚集分布有利于相互庇护,提高存活率[17,23]。但随着龄级的增大,个体对资源的需求增强,种内与种间竞争加剧,种群个体死亡率提高,聚集强度减弱或趋向于随机分布[24-25]。
岷江冷杉的幼树和小树在中小尺度上(25m以内)呈正关联,与种子扩散机制有关。岷江冷杉短距离散布种子的方式常形成聚集分布,由于其耐阴性使得聚集分布格局持续到小树阶段。在这一过程中,小树为幼树成长提供了良好的生存环境,起到了庇护作用[19]。这也说明幼树和小树的相互作用主要发生在20多m内的范围内,超出这个范围,相互作用将减弱。幼树和小树均与大树表现出空间独立,说明大龄级个体对中幼龄个体非对称作用的竞争不明显[26]。
枯立木对岷江冷杉有着积极作用,各生长阶段个体均与枯立木表现出一定的正相关关系,这和当地的干扰机制影响更新有关。以往研究表明,该地区林窗更新模式占主导作用,幼龄阶段岷江冷杉的更新数量和聚集程度与林窗大小有关[27]。这就解释了幼树在小、中、大尺度上(1~20、35~37m和40~41m)均与枯立木表现出正相关关系,无论是小尺度下的林隙或林窗,还是大尺度上林窗斑块的镶嵌分布,都有利于岷江冷杉的更新。另外,枯立木的出现有利于小树的生长(即由林下层进入主林层),故小树在大部分空间尺度上与枯立木呈正相关。但是最终能够进入到主林层的个体占少数,这就解释了大树只在小尺度上(1~6m)与枯立木呈正相关。
总体而言,岷江冷杉在不同生长阶段和多数空间尺度上的空间格局趋于聚集分布,种内空间关联(含枯立木)主要呈正相关关系。这种聚集分布和空间正关联体现了种群内部正向和相互有利的生态关系[25]。结合该地区的干扰机制,岷江冷杉的生物学特性、更新机制和空间属性,可对群落进行适度的人为干扰,以促进岷江冷杉林的进展演替。
1) 针对岷江冷杉种群聚集分布和种内(幼树和小树)正关联的特性以及物种的耐阴性,在幼龄阶段可以保持较高种群密度,便于整个种群的生长和繁衍。
2) 针对各生长阶段与枯立木正关联的特性,可适当进行主林层的透光抚育,创造良好的光照条件,以促进种群发展,尤其是幼树和小树的生长发育。
3) 针对该地区竹子对林窗更新有较强的抑制作用[28],可对林下层进行抚育,清除部分箭竹和其他灌木,为幼苗幼树创造适当的生存环境。另外,岷江冷杉的空间格局还可能受到其他因素的影响,比如种间竞争(与伴生种的空间关联以及林下灌草的影响),这些作用机制是下一步应该深入研究的方向。
[参考文献]
[1]Druckenbrod D L, Shugart H H, Davies I. Spatial pattern and process in forest stands within the Virginia piedmont[J]. Journal of Vegetation Science, 2005, 16(1): 37-48.
[2]惠刚盈,Klaus von Gadow,胡艳波,等.结构化森林经营[M].北京:中国林业出版社,2007.
[3]Wang X G,Wiegand T,Hao Z Q, et al. Species associations in an old-growth temperate forest in north-eastern China [J]. Journal of Ecology,2010,98:674-686.
[4]张春雨,赵秀海,夏富才.长白山次生林树种空间分布及环境解释[J].林业科学,2008,44(8):1-8.
[5]张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):57-62.
[6]肖文娅,周琦,董务闯,等.苏州太湖流域2种针阔混交林林分空间结构特征比较[J].西南林业大学学报,2014,34(4):8-13.
[7]严绍裕.天宝岩自然保护区长苞铁杉不同群落结构与空间格局比较[J].西南林业大学学报,2013,33(4):44-48.
[8]王巍,刘灿然,马克平,等.东灵山两个落叶阔叶林中辽东栎种群结构和动态[J].植物学报,1999,41(4):85-92.
[9]张赟,张春雨,赵秀海,等.长白山次生林乔木树种空间分布格局[J].生态学杂志,2008,27(10):1639-1646.
[10]管中天,陈尧,徐润青.峨眉冷杉林森林类型的研究[J].植物生态学与地植物学丛刊,1984,8(2):133-145.
[11]刘世荣,孙鹏森,王金锡,等.长江上游森林植被水文功能研究[J].自然资源学报,2001,16(5):451-456.
[12]程伟,吴宁,罗鹏.岷江上游林线附近岷江冷杉种群的生存分析[J].植物生态学报,2005,29(3):349-353.
[13]李德文,高跃,高志勇.川西亚高山岷江冷杉林小气候特征[J].四川林业科技,2006,27(6):30-34.
[14]张远彬,王开运,鲜骏仁.岷江冷杉林林窗小气候及其对不同龄级岷江冷杉幼苗生长的影响[J].植物生态学报,2006,30(6):941-946.
[15]程伟,罗鹏,吴宁.岷江上游林线附近岷江冷杉种群(AbiesfaxonianaRehd.et Wild)的生态学特点[J].应用与环境生物学报,2005,11(3):300-303.
[16]鲜骏仁,胡庭兴,张远彬,等.林窗对川西亚高山岷江冷杉幼苗生物量及其分配格局的影响[J].应用生态学报,2007,18(4):721-727.
[17]缪宁,史作民,冯秋红,等.川西亚高山岷江冷杉种群的空间格局分析[J].林业科学,2008,44(12):1-6.
[18]缪宁,刘世荣,史作民,等.青藏高原东缘林线杜鹃-岷江冷杉原始林的空间格局[J].生态学报,2011,31(1):1-9.
[19]缪宁,刘世荣,史作民,等.川西亚高山红桦-岷江冷杉林优势种群的空间格局分析[J].应用生态学报,2009,20(6):1263-1270.
[20]Ripley B D. Modelling spatial pattern[J].Journal of the Royal Statistical Society.Series B,1977,39(2):217-212.
[21]赵常明,陈庆恒,乔永康,等.青藏高原东缘岷江冷杉天然群落的种群结构和空间分布格局[J].植物生态学报,2004,28(3):341-350.
[22]Besag J E. Comments on Ripley’s paper[J]. Journal of the Royal Statistical Society,1977,39(2):193-195.
[23]张赟,张春雨,赵秀海.内蒙古黑里河天然油松林主要树种的空间分布格局[J].西北植物学报,2009,29(1):167-173.
[24]Hutchings M J. Standing crop and pattern in pure stands of Mercurialis perennis andRubusfruticosusin mixed deciduous woodland[J]. Okios,1978,31(3):151-357.
[25]Kenkel N C. Pattern of self-thinning in jack-pine-testing the random mortality hypothesis[J]. Ecology, 1988, 69(4): 1017-1024.
[26]Hou J H,Mi X C,Liu C R,et al. Spatial patterns and associations in aQuercus-Betulaforest in northern China[J]. Journal of Vegetat Ion Science,2004,15(3):407-414.
[27]申国珍,李俊清,任艳林,等.大熊猫栖息地亚高山针叶林结构和干扰关系研究[J].北京林业大学学报,2002,24(5):115-119.
[28]申国珍,李俊清,蒋仕伟.大熊猫栖息地亚高山针叶林结构和动态特征[J].生态学报,2004,24(6):1294-1299.
(责任编辑赵粉侠)
第1作者:梁璇(1989—),女,硕士生。研究方向:森林经理。Email:sharonleung311@163.com。
通信作者:刘萍(1964—),女,博士,教授。研究方向:森林经理和林业系统工程。Email:bengtiaoliu@126.com。

