童年压力及死亡威胁启动对择偶要求的影响*
2016-01-10汪佳瑛陈斌斌
汪佳瑛 陈斌斌
(1 Teachers College,Columbia University,New York,NY 10027 U.S.A.)(2复旦大学心理学系,上海 200433)
1 引言
一直以来,择偶都是进化心理学的一项核心议题。每个个体都通过择偶来获取繁衍的成功,将基因传递给后代,并发展出对所处环境的适应力(Buss,2007)。而择偶要求则是择偶活动顺利进行的一个基础。没有人会对周围所有人都产生相等的择偶欲望。每个个体都有其择偶的要求和偏好,这些要求和偏好会影响个体所采用的择偶策略,以保证择偶及最终繁衍的成功。
已有众多学者研究了与择偶要求有关的变量,其中包括择偶要求的具体内容(好基因、资源以及好父母) (Li,Bailey,Kenrick,&Linsenmeier,2002;Lu,Zhu,&Chang,2015;Shackelford,Schmitt,&Buss,2005)、男女在择偶要求上的性别差异以及对不同关系性质的配偶(长期伴侣或短期伴侣)择偶要求的差异(Buss,2007)。从进化心理学的角度而言,这些择偶偏好都为个体的成功繁衍及基因延续提供了帮助。
然而在众多有关择偶要求的研究中,目前国内外鲜有研究涉及择偶要求的变化以及环境对择偶要求的影响(陈斌斌,2016)。事实上,一个人的择偶要求并非一成不变。个体需要采用适应于环境的择偶策略以使自己的繁衍得以成功(Chen &Chang,2015)。由于个体所处的环境对形成及改变个体的择偶要求有重要作用,因此环境对择偶要求的影响需要更多的研究予以探索。而生命史理论则从进化的角度为环境(特别是童年环境)对个体行为倾向和决策的影响提供了研究及解释的途径(Chen &Chang,2016;Del Giudice,Gangestad,&Kaplan,2015;Ellis,Figueredo,Brumbach &Schlomer,2009)。本研究试图在生命史理论的框架下,研究童年期的环境压力如何与当前环境中的压力线索相互作用并影响个体对异性择偶要求的变化。
2 生命史理论
2.1 生命史理论的提出及主要内容
由于每个生物体都只拥有有限的时间、精力等资源,因此生物如何在众多与生存繁衍有关的活动中分配这些资源,以使资源利用最大化成为生物学中重要的问题。进化生物学中的生命史理论就试图回答这一问题(Mittal &Griskevicius,2014)。生命史理论认为,因为每个生物体所能获得的生物能量及物质资源都是有限的,所以个体需要权衡其在不同的、甚至互相冲突的生命任务中的资源分配,其中包括身体投资以及繁衍投资。前者意味着个体将资源用于维持个体的生存,包括维持身体健康以及成长;而后者则意味着个体将资源用于繁衍后代以使其基因得以延续(Ellis et al.,2009;Figueredo et al.,2006)。由于资源的有限性,生物体在以上各方面的资源投入不可能全部最大化,因此只能从中进行取舍。例如,生物体投入于身体成长的资源会使其生长并拥有较大的体型,但同时其性成熟时间会相应延迟,即较长的生长期是以推迟繁衍时间为代价的;相反,如果资源被用于当前的繁衍任务,则个体就无法同时兼顾身体的成长。生命史理论认为,为了更好的适应环境条件,人们采取不同的策略将有限的时间和资源分配给不同活动、分配给现在及未来(陈斌斌,2016;Chen &Chang,2016)。因为分配到某一方,另一方就得不到分配,所以分配是一个权衡的过程(Kaplan &Gangestad,2005;Stearns,1992)。例如,个体把有限的能量分配到促进免疫系统功能的话,就不能分配到促进吸引伴侣的功能上去了。何时以及如何分配时间和资源就建构了生命史策略,它体现在快和慢的连续维度上(Ellis et al.,2009;Figueredo,Vásquez,Brumbach,&Schneider,2004;Nettle,2010)。慢策略认为身体投资优先于繁衍投资,因此采用慢策略的个体性成熟时间较晚,会在较晚的年龄开始性行为及生育;与此相反,快策略则认为繁衍投资优先于身体投资,因此他们性成熟的时间较早,并且会在较早的年龄就开始性行为及生育(Belsky,Steinberg,&Draper,1991;Copping,Campbell,&Muncer,2013;Ellis,2004;Griskevicius,Delton,Robertson,&Tybur,2011;Nettle,2010;Simpson,Griskevicius,Kuo,Sung,&Collins,2012)。除此之外,快、慢策略在其他众多方面也具有较大的行为表现差异(陈斌斌,2016)。采用慢策略的个体倾向于拥有较少的性伴侣(Belsky,Schlomer,&Ellis,2012)和较少的子女并且在每个子女上投入的资源较多,注重子女的质量并因此在择偶中有更高的标准(Dillon,Adair,Wang,&Johnson,2013);他们会规避风险,倾向于将资源集中投入一个方面,并能够延迟满足(Griskevicius,Tybur,Delton,&Robertson,2011;Griskevicius et al.,2013;White,Li,Griskevicius,Neuberg,&Kenrick,2013);同时有较少的攻击性行为、不良行为及犯罪行为(Copping et al.,2013;Simpson et al.,2012)。而快策略则倾向于拥有更多的性伴侣、想要更多的子女且在每个子女身上投入的资源较少;他们偏好风险和多样化的资源分配方式,不喜欢延迟满足;并且会有更多的攻击性行为、不良行为及犯罪行为。两种策略所表现的不同行为倾向并不意味它们具有好坏之分。这两种策略应看作是一个连续体的两端(Figueredo et al.,2006),影响个体在这一连续体中的选择的是个体所处的环境。而不同的生命史策略应被解读为是个体为了适应不同生活环境而产生的行为及心理上的适应结果(陈斌斌,2016;Chen &Chang,2016;Ellis et al.,2009)。
2.2 童年环境压力对生命史策略的影响
生命史理论的研究显示,人们在童年期经历的环境压力是促使不同生命史策略产生的主导根源(Belsky et al.,1991,2012)。当个体在童年期处于压力较大的环境(如资源缺乏或死亡率高等)中,个体会发展出相应的策略以适应该环境。Ellis等人(2009)通过对生物学、生态学及心理学的前人研究进行整合和分析,从童年生活环境中提取出了对影响生命史策略的进化及发展最为基础的两个环境因素,即环境的严峻性和不可预测性。其中,严峻性是指在这一环境里,某一特定年龄层的人群中由外因所导致的致病率和死亡率,而不可预测性则是指环境严峻性随时间空间变化的频率(Ellis et al.,2009)。
当人们童年期所处的环境较为严峻(如死亡率高、资源缺乏、环境不稳定等)时,个体的预期寿命会较短(Copping et al.,2013;Simpson et al.,2012),因此为了保证在个体死亡前能够顺利地繁衍,将自己的基因传递给后代,个体会采用较快的生命史策略,即将资源更多地投入于繁衍而非身体成长。同时为了保证其后代有较高概率能够存活到成年繁衍期,个体会选择生育更多的子女,而不注重子女养育的质量。相反,当个体童年期所处的环境较好(如死亡率低、资源丰富、环境稳定等)时,个体的预期寿命较长,个体倾向于将资源更多地投入自身身体的维持及成长而非繁衍中,即采取较慢的生命史策略。
在此类研究中,社会经济地位(Socioeconomic status;SES)这一指标通常用于测量个体童年期所处环境的严峻性和不可预测性(Griskevicius et al.,2013;Griskevicius,Delton,et al.,2011;Griskevicius,Tybur,et al.,2011;Simpson et al.,2012;White et al.,2013)。研究显示,社会经济地位的高低与死亡率具有线性关系,在SES较高的环境中致病率及死亡率更低,而在SES较低的环境中这些比率则更高(Ellis et al.,2009)。同时,社会经济地位较低的儿童所处的环境也更不稳定(Evans,Gonnella,Marcynyszyn,Gentile,&Salpekar,2005)。同时,值得注意的是,Griskevicius等人(Griskevicius,Delton,et al.,2011;Griskevicius,Tybur,et al.,2011)的研究结果显示,相比于被试当前的SES,其童年期的SES对其生命史策略的影响更大,这也正好符合生命史理论的核心思想,早期的环境会让个体按照某种特定的发展轨迹采取相应的行为策略(Belsky et al.,1991)。
2.3 当前环境压力对生命史策略的影响
除了童年环境压力外,当前环境压力也会对个体所采用的生命史策略产生影响(参见陈斌斌,2016对该问题的详细论述)。研究显示,预期寿命较短(高死亡率的指标之一)、收入较低的女性会在更早的年龄进行生育(Nettle,2011),并且预期寿命与高中毕业率、大学毕业率和求学的时长均有显著的正相关,即预期寿命越长,毕业率越高,求学时间越长(Krupp,2012)。Promislow和Harvey (1990)对哺乳动物的研究显示,少年期的死亡率与成年期死亡率呈显著正相关,而在控制了少年期死亡率后,成年期死亡率仍与个体成熟年龄显著相关。Chisholm (1993,1996)提出当前环境的死亡率是进化史中的一个重要生态线索,它会影响自然选择的压力,并由此使人们产生相应的心理机制以适应这一环境,即采用较快或较慢的生命史策略。
童年期和当前的环境压力会共同影响个体生命史策略的选择。研究发现,在通过死亡威胁启动使被试更强烈地感受到当前环境中的较高严峻性和不可预测性后,来自不同童年经济地位的被试原先无差异的生命史策略表现会呈现出显著的差异性(Griskevicius,Delton,et al.,2011;Griskevicius,Tybur,et al.,2011;Griskevicius et al.,2013;Hill,Rodeheffer,DelPriore,&Butterfield,2013;Laran &Salerno,2012;White et al.,2013)。Griskevicius等人(Griskevicius,Delton,et al.,2011;Griskevicius,Tybur,et al.,2011)最先通过实验对此进行了研究,他们让一部分被试阅读一篇描述近期暴力事件导致死亡率上升或描述近期经济衰退形势严峻的新闻文章作为当前环境压力的启动,另一部分被试阅读一篇描述一个人花了很长时间寻找遗失钱包的文章作为控制组。结果发现在控制组中,来自不同童年经济地位的被试的生命史策略的表现并没有显著差异;而在实验组中,童年社会经济地位较低的被试会显著表现出快策略的特征,如变得更为冲动、冒险、想要更早且更多地进行生育,童年社会经济地位较高的被试则显著表现出慢策略的特征,如变得更不冲动、更规避风险、想要延迟生育。之后的一些研究者借用这一范式也得到了相似的结果(Griskevicius et al.,2013;Hill et al.,2013;Laran&Salerno,2012;White et al.,2013)。因此,当人们面对当前环境中的压力线索时,每个人都会根据其童年所经历的压力表现出与其生命史策略相关的不同的反应。
3 择偶要求及其变化
通过前人大量的研究,研究者们已经提取出了择偶要求的3个主要维度,即身体吸引力、资源以及好父母(陈斌斌,2016;Lu et al.,2015)。具有身体吸引力通常代表着个体拥有较好的基因(Barber,1995),而这些基因有一定概率能够传递给后代,以影响后代的质量。研究显示,虽然男女两性在择偶过程中都偏好身体更具吸引力的异性,但男性相比女性而言更加注重这一指标(Buss,2007)。且采用不同生命史策略的个体对异性身体吸引力的择偶价值也存在差异,其中采用慢策略的人对自己及配偶身体吸引力的择偶价值的评分更高,这与采用慢策略的人更注重子女质量相符(Dillon et al.,2013)。同时,虽然两性在择偶中都看重对方能为自己提供资源,但资源的内容并不相同(Sefcek,Brumbach,Vasquez,&Miller,2006),且女性更加注重对方获取金钱和资源的能力(Buss,2007)。同时,男女两性在择偶中都会重视对方成为好父母的潜质,因为人类幼儿的成长期较长,且在一夫一妻制度下父母是子女最直接的照料者,后代的生存和成长离不开父母所提供的食物、保护以及教育(Sefcek et al.,2006)。从进化心理学视角看,这些择偶要求及其中的差异等都是个体为保证其择偶和繁衍的成功而形成的(Buss,2007;Eagly &Wood,1999)。
在有关择偶要求的研究中,已有大量文献基于性选择的理论来探讨男女在择偶标准上的性别差异(陈斌斌,2016)。相比之下,从其他进化观点或者角度研究择偶要求高低差异以及择偶要求变化的研究只占了少数。这其中的多数研究都聚焦于在被试的择偶动机被唤起后,被试为提升自身吸引力而做出的行为改变(Hill,Rodeheffer,Griskevicius,Durante,&White,2012;Janssens et al.,2011),或对资源等择偶过程中重要因素的偏好改变(Roney,2003)。而且,值得注意的是,只有少数文献研究了个体对异性择偶要求的变化或高低差异。
Yong和Li (2012)的研究显示,当男性被试被给予了大额金钱后,相比于被给予小额金钱和没有被给予金钱的被试,他们对异性身体吸引力的最低择偶要求显著升高,但女性被试没有显著区别。这一结果说明,当男性拥有较多资源时他们处于择偶竞争中的优势地位,不用过于担心或急于择偶,因此只有当异性质量较高时他们才想与之建立配偶关系。Spielmann等人(2013)使用自己开发的工具,测量个体对单身恐惧。他们通过问卷法显示对单身更为恐惧的个体更倾向于维持低质量的恋爱关系,作为对择偶要求妥协的一种表现,并且在恋爱关系质量较低的情况下更少主动提出分手。该研究同时通过实验让单身被试对实验者虚构的异性网上相亲档案进行兴趣的评价,发现对于档案中表现得更不负责和更不具魅力的对象,对单身较恐惧的被试比对单身不恐惧的被试表现出更大的兴趣,即他们的择偶要求更低。而在真实的快速约会中进行的问卷调查也发现对单身恐惧较大的被试对潜在对象有兴趣的数量更多,对约会对象更不挑剔,且男性对潜在对象有兴趣的数量多于女性。研究者认为这是因为降低择偶要求能够增大潜在的约会对象的范围,从而降低单身的风险。然而同时,该研究通过让被试自我报告其对异性的择偶要求,发现对单身较恐惧的个体在潜在对象做好父母的择偶要求上却更高。研究者认为这一结果说明对单身的恐惧部分是出于对没有子女的恐惧,他们对拥有子女和家庭更为重视,因此会对对方做好父母的要求更高。上述这些研究尽管没有基于生命史理论来探讨,但是可以显示个体自身的特点以及环境都会影响择偶要求的变化。
综上所述,由于目前对择偶要求变化的研究数量仍较少,且缺少较为系统的理论予以解释,因此本研究着眼于个体择偶要求随环境因素而发生的变化并试图从生命史理论的角度对择偶要求的变化进行解释。如前文所述,在个体感受到当前环境的压力时,其采用的生命史策略的特征会表现得更为明显。其中,童年压力较小的个体在当前环境压力下会表现出慢策略的特征。他们会推迟生育繁衍时间不急于择偶繁衍,将资源更多地投入于自身身体的成长和发展,并注重子女的质量。因此他们会相应升高对异性身体吸引力、资源和好父母的择偶要求,以获得质量更高的异性伴侣。相反,童年压力较大的个体会表现出快策略的特点,倾向于尽早繁衍并寻求更多的繁衍机会。而这种对择偶繁衍的动机以及其对子女质量的不重视会驱使个体降低其对异性身体吸引力和资源的择偶要求,以扩大择偶对象的选择范围,增加繁殖的机会和成功率。同时根据 Spielmann等人(2013)研究中有关做好父母的结果,本研究假设童年压力较大的个体在当前环境压力下对异性做好父母的择偶要求也会升高。
4 实验设计
4.1 被试
本实验被试为70名本科生(11名男生,59名女生),年龄分布为 17~23 岁(M
=19.37,SD
=1.16)。其中18人正处于恋爱关系中,52人处于单身状态中。所有被试中有一人为双性恋,其余均为异性恋。所有被试都通过参加实验换取相应课程分数。4.2 实验过程
本研究有两个被试间的启动条件:死亡威胁启动及控制条件。被试被随机分配到死亡威胁启动组或控制条件组。被试先阅读两篇新闻文章中的一篇。第一篇文章用于启动死亡威胁,第二篇文章用于控制条件。在阅读了新闻文章后,被试分别对理想择偶要求进行评分,并报告他们的童年压力。除阅读文章外,其余的实验步骤都在 Inquisit程序上完成。
为了减少被试对实验目的产生可能的猜疑以及欺骗行为,本研究采用单盲实验。被试被告知这是一个由多个测试组成的实验,第一部分是有关记忆的实验。被试首先需要阅读一篇新闻文章,并且他们需要在之后的实验部分中对这篇文章的内容进行回忆测试。而由于在回忆测试前需要有足够的时间用于记忆消退,他们需要完成一份有关择偶要求的评分问卷。通过实验结束后的询问,没有被试对所告知的实验目的产生怀疑。
4.3 实验材料
4.3.1 死亡威胁启动
根据Griskevicius等人(Griskevicius,Delton,et al.,2011)的实验程序,被试首先阅读一篇虚构的新闻文章。启动死亡威胁的新闻文章的标题为《暴力事件频发,治安形势严峻》,内容是关于近期国内随机暴力犯罪及死亡率有上升趋势。文章着重强调了随机的死亡变得越来越常见以及环境的不可预测。被试被告知这篇新闻文章是新华网的一篇在线新闻,而选取这篇文章是因为其长度大约 1000字,是记忆测试最理想的长度。文章被设计成与真实的新华网在线新闻的格式相同,包括网站的标志和字体,以追求新闻的真实感。
控制条件所使用的新闻文章同样是虚构的,并且格式和长度都与死亡威胁条件所使用的文章相同,并且将其设计为能达到相同水平的一般情绪唤起。控制组文章的内容是一个人在某个周二下午花了很长时间寻找遗失的钱包的事件。
在确保两篇文章能唤起相同水平的一般情绪的基础上,死亡威胁组的文章还能唤起被试对周围环境的不安全感和不确定感。为此,研究者在正式实验前进行了预实验。预实验的被试均是大学本科生,共有16人(5男,11女),其中一半阅读死亡威胁条件的文章,另一半阅读控制组的文章。预实验中的指导语与正式实验相同。在预实验的最后,被试需要对5个问题进行回答。前两个问题评估被试阅读之后的安全感:1)这篇文章使你感觉环境变得更加危险的程度是多少?2)这篇文章使你感觉环境变得不安全的程度是多少?第三、四个问题评估被试阅读后的不可预测感和不确定感:3)这篇文章使你感觉环境变得更加不可预测的程度是多少?4)这篇文章使你感觉环境变得更加不确定的程度是多少?最后一个问题评估被试的一般情绪唤起:5)这篇文章使你唤起情绪的程度是多少?所有问题都使用1(一点也不)到7(非常强烈)的7点量表进行评分。
对预实验的数据进行独立样本 t检验,结果显示与控制条件的文章相比,死亡威胁条件所使用的文章使被试感到环境更加危险、更不安全、更不可预测、更不确定,但两篇文章所唤起的一般情绪水平相同。因此实验所使用的两篇文章能够达到实验所要求的启动效果。具体t检验结果见表1。
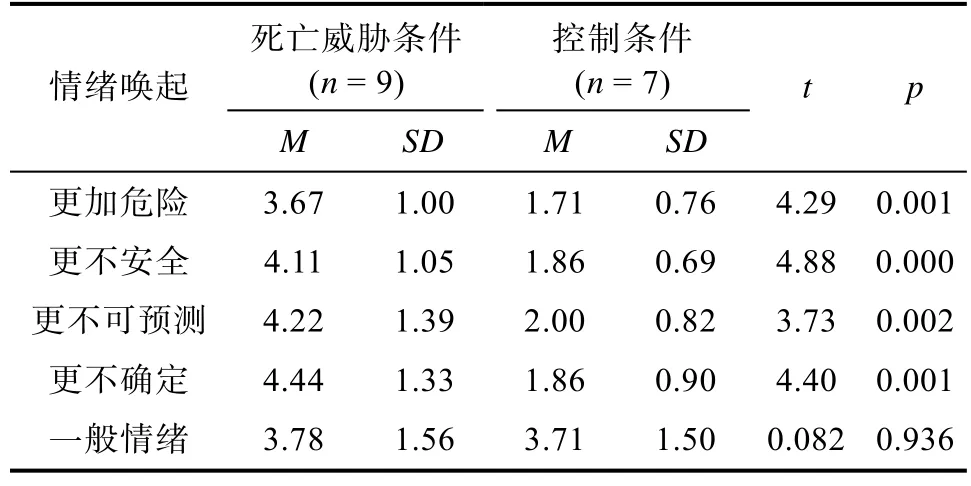
表1 启动材料预实验独立样本t检验结果
4.3.2 择偶要求
理想择偶要求的测量采用Spielmann等人(2013)使用的量表。被试根据其对伴侣的理想要求,分别对9项特质进行评分。评分采用百分数,被试回答其理想异性的某一特质的水平需要处于整个异性群体中的百分之几以上。百分数范围为0%~100%。
9项择偶要求由3个维度组成,分别包括3项有关身体吸引力的标准(具有身体吸引力、外表好看、性感;α=0.75),3项有关资源的标准(收入高、有较好的赚钱能力、有较好经济前景;α=0.94),3项有关好父母的标准(渴望家庭和孩子、注重孩子的抚养、喜欢孩子;α=0.92)。
4.3.3 童年压力
被试的童年期压力采用 Griskevicius等人(Griskevicius,Delton,et al.,2011;Griskevicius et al.,2013)所使用的量表进行测量,其项目包括:(1)我成长于一个相对富有的社区;(2)与学校中的其他孩子相比,我觉得我相对而言更加富有;(3)在我成长过程中,我通常有足够的资源来做我想做的事;(4)在我成长过程中,我的家庭通常有足够的钱来购买所需物品。量表为7点量表,评分从1 (非常不符合)到7(非常符合),分数越小则童年压力越大,α=0.77。
5 结果
5.1 描述性统计及各变量相关系数
本研究首先进行了各变量的描述性统计及变量间的相关分析,具体结果见表2。
5.2 性别、恋爱状态的组间差异
通过独立样本 t检验,本文首先检验在理想择偶要求的3个维度上的性别差异。结果显示,在对于资源的择偶要求上男女有显著地性别差异,t
(68)=5.78,p
<0.001,Cohen’sd
=1.93,女性对异性资源的要求显著高于男性。其次,在对于资源的择偶要求上,单身和处于恋爱关系中的被试存在显著差异,t
(68)=2.74,p
=0.008,Cohen’sd
=0.76,处于恋爱关系中的被试对对方资源的要求显著低于单身的被试。具体结果见表3。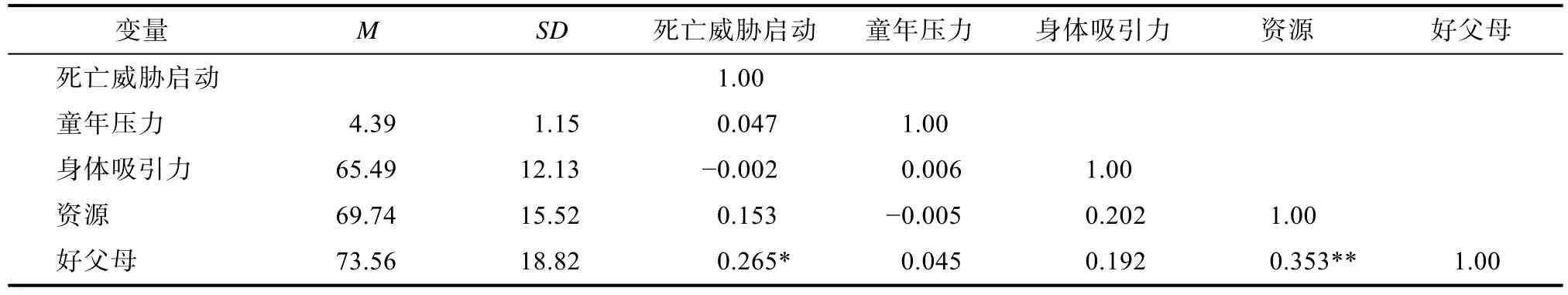
表2 各变量的描述性统计及各变量间相关分析结果
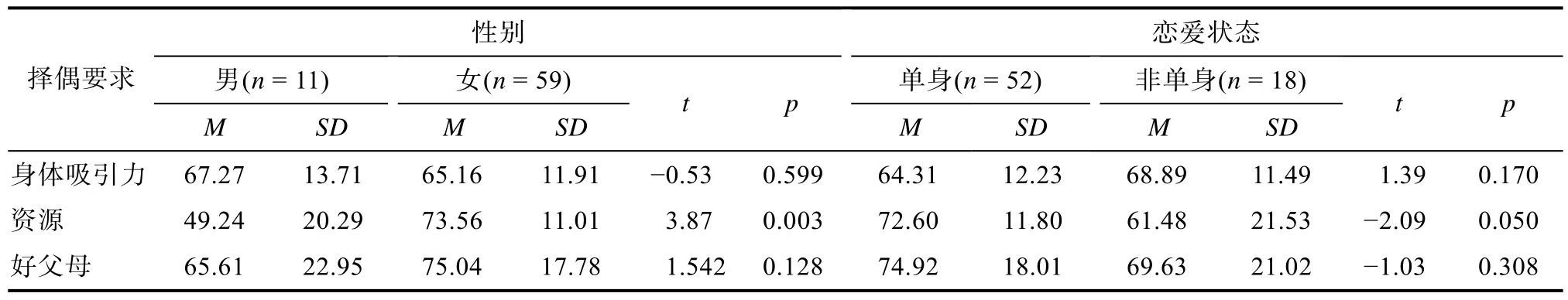
表3 性别及恋爱状态在择偶要求的3个维度上的独立样本t检验结果
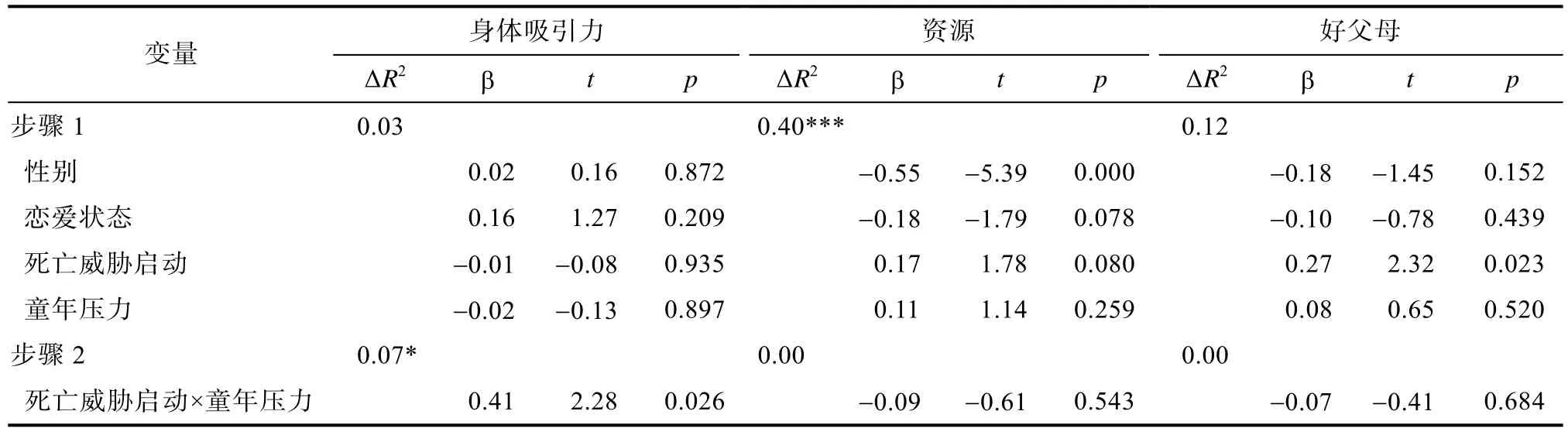
表4 童年压力及死亡威胁启动在对异性身体吸引力、资源及好父母择偶要求上的回归分析
5.3 死亡威胁启动与童年压力在择偶要求上的交互作用
通过调节回归分析,在控制了性别及恋爱状态的情况下,对童年压力与死亡威胁启动在理想择偶要求的3个维度上的交互作用进行分析。结果显示,童年压力与死亡威胁启动在异性身体吸引力的理想要求上没有主效应,但两者存在显著的交互作用。随后通过一般线形模型(GLM)进行简单效应的检验,结果发现,在低童年压力组,死亡威胁启动组对异性身体吸引力的理想要求显著高于控制条件组,F
(1,66)=5.40,p
=0.02;在高童年压力组,死亡威胁启动组对异性身体吸引力的理想要求显著低于控制条件组,F
(1,66)=5.01,p
=0.03。取童年压力均值1个标准差以下及以上作为横轴作图得到图1。在好父母的理想要求上,启动条件存在显著的主效应,死亡威胁启动下的被试对异性做好父母的择偶要求高于控制条件的被试,但童年压力不存在主效应,且两者不存在交互作用。在对资源的择偶要求上,既不存在童年压力和死亡威胁启动的主效应,也不存在交互作用。具体回归分析结果见表4。
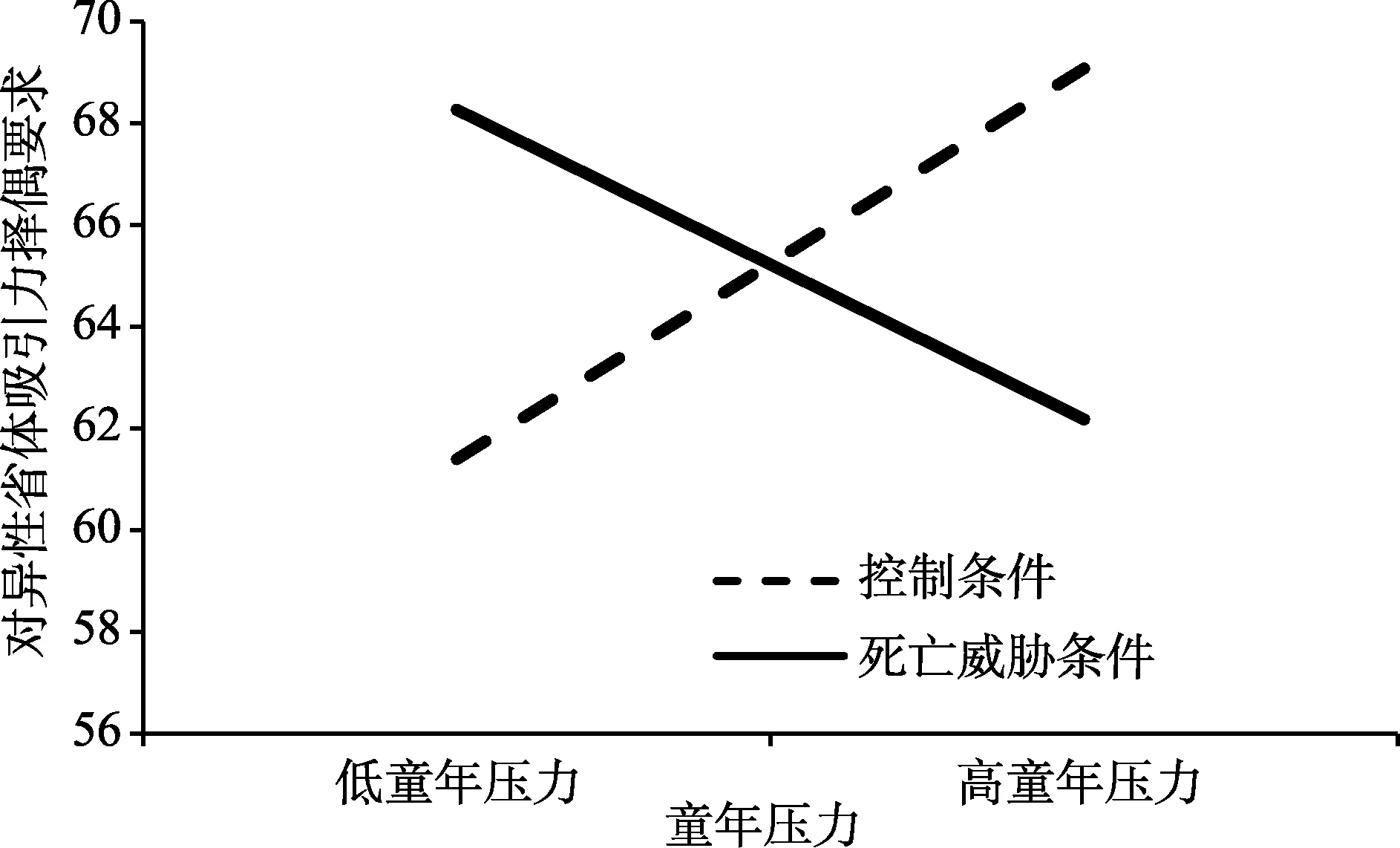
图1 童年压力与死亡威胁启动在对异性身体吸引力的择偶要求上的交互作用
6 讨论
6.1 对实验结果的分析与讨论
本研究从生命史理论的角度出发,研究了个体童年期所处的环境压力与其在当前环境中感受到的环境压力如何通过交互作用影响个体对异性的择偶要求。研究结果显示,童年压力与启动条件仅在身体吸引力的理想要求上有显著的交互作用。其中,低童年压力组在死亡威胁启动下对异性身体吸引力的理想要求显著高于控制条件,而高童年压力组在死亡威胁启动下对异性身体吸引力的理想要求则显著低于控制条件。此外,启动条件在好父母的理想要求上有显著的主效应,死亡威胁启动下的被试对异性做好父母的择偶要求高于控制条件的被试。而在对资源的择偶要求上则既不存在童年压力和启动条件的主效应,也不存在交互作用。
童年压力与死亡威胁启动在异性身体吸引力的理想要求上的显著交互作用与本研究假设相符。根据Griskevicius等人(Griskevicius,Delton,et al.,2011;Griskevicius,Tybur,et al.,2011)提出的敏感性模型,生命早期的生活状态已经形成独特的生命史策略,会影响个体在成年期遇到不同的生活境况时作出相应的不同反应模式。经历了不同童年环境的成人在良好的当前环境中,由于当前环境的宜人性而可能表现出相似的行为模式,但在当前环境中遇到困境时则会表现出不同的行为模式予以应对。因此当童年压力较大的被试通过死亡威胁启动感受到较大当前环境压力后,其采用较快的生命史策略使其获得更多的繁衍机会。而在繁衍的动机被唤起后,他们对异性身体吸引力的要求降低,意味着他们不再关心择偶异性身体吸引力的优劣。而在进化心理学中,身体吸引力通常与好基因有关(Barber,1995;Lu et al.,2015),代表着异性的健康等方面的质量(Nedelec &Beaver,2014),且这一基因质量会遗传给后代。因为采用快策略的个体急于繁衍,并且他们将自身资源更多地投入于繁衍而非对后代的抚养,注重孩子的数量而非质量,所以童年压力较大的个体对异性可能遗传给后代的基因质量并不重视,导致其对异性身体吸引力的要求更低,以扩大其择偶范围,增加其繁衍的机会和成功率。相反,童年压力较小的个体在感受到当前环境的压力后,会表现出慢策略的特征。相比于子女的数量,他们更注重子女的质量及对子女的抚养,因此他们对异性的基因更为重视,导致其对异性身体吸引力的要求升高。
而在好父母的理想要求上,本研究仅发现启动条件存在主效应,即死亡威胁启动下的被试对好父母的择偶要求高于控制条件的被试;但交互作用不显著。这一结果与研究假设相符。根据亲本投资理论,在一夫一妻制下,父母的亲本投资不仅影响后代的发育发展,更重要的是直接关系到后代的生存率和死亡率(Geary,2000)。在相对恶劣的环境中,后代需要父母的养育才能保证他们的存活及其基因的延续。因此不管自己的童年经历是好的还是坏的,一旦在当前的死亡威胁启动下,个体所感知到的当前环境的恶劣性,让人们更看重伴侣能够为养育后代提供更好的合作与支持,都希望异性能够具有做好父母的潜质,以代替或帮助自己抚养后代,这样有利于后代的存活。
此外,本研究结果还显示,在对异性资源的择偶要求上既不存在童年压力和启动条件的主效应,也不存在交互作用,与研究假设不符。对这一结果可能的解释是:在择偶中,对异性资源方面的要求是为了让自己可以在择偶中从对方那里获取额外的资源(Buss,1989),以提高自己及子女的生活质量。但对于童年压力较大的个体而言,在死亡威胁下,作为较快生命史策略的特征,他们关注的是繁衍的数量,而不是自身的生活质量或其子女的抚养和生活质量,因此他们在择偶过程中并不考虑异性资源的多少,导致其对资源的要求在启动条件下没有显著变化。而对于童年压力较小的个体而言,在死亡威胁下,他们所采用的慢策略使他们将自己的资源投入于对后代的教养以保证后代的质量,因此在这一基础上,异性能够遗传给后代的基因的质量比异性提供的资源对后代质量的影响作用更大,所以慢策略个体对异性的资源的要求也不会显著提高。
综上所述,根据实验结果,本研究认为,在死亡威胁下,童年压力较大的个体因为其采取的较快生命史策略注重繁衍的数量而不注重繁衍后代的质量,所以相比于控制条件,这些个体会降低对代表异性身体健康及基因质量的身体吸引力这一标准的要求,以扩大择偶范围增加繁衍的机会和成功率。但同时为了确保后代在严厉环境下的存活率,他们会提高对异性做好父母的标准要求,以希望在自己不投资于后代抚养的情况下,异性能够为后代提供抚养。而因为童年压力较小的个体采用较慢的生命史策略,所以在死亡威胁下,他们会更多地投资于孩子的教养,重视后代的质量,因此他们会提高对异性的身体吸引力及做好父母的标准的要求。由于在择偶过程中对异性资源的要求更多的是为了获取对方的资源以提高自己及后代的生活质量,因而对于并不注重子女质量的快策略个体和已经将自己资源投入于子女教养的慢策略个体而言,在死亡威胁下异性的资源不是他们考虑的重点,所以对于异性资源的要求没有任何显著的差异和变化。
6.2 生命史理论与相关理论的比较
本研究的假设及对结果的分析均基于生命史理论中环境压力对人们行为及心理活动的影响所提出的。然而,在其它理论中也有就死亡线索对个体择偶及其他行为影响的研究,其中包括恐惧管理理论(Terror Management Theory;Greenberg,Pyszczynski,&Solomon,1986)。恐惧管理理论主要研究人们受到最终死亡的提醒对其各种行为产生的影响。虽然两种理论探讨的都是死亡对人们的影响,但其中死亡的含义并不相同。恐惧管理理论的实验通常让被试自己想象并书写一段有关自己的死亡的文字做为对最终死亡的启动(Rosenblatt,Greenberg,Solomon,Pyszczynski,&Lyon,1989),而生命史理论中的死亡威胁则是来自于环境中的外在的高死亡率(Chisholm,1993)。然而也有学者指出,这两种死亡启动可能产生了相似的心理机制(Griskevicius,Delton,et al.,2011)。但即便如此,恐惧管理理论仍不能完全解释本研究中所得到的当前环境中的死亡威胁对个体异性择偶标准的影响,并且进化心理学的研究者并不认为恐惧管理理论属于进化观的理论(Buss,1997)。同时,恐惧管理理论主要聚焦于最终死亡的提醒对自尊的影响(Greenberg et al.,1986),而生命史理论则更关注于个体如何进行资源的分配以更适应于环境(Mittal &Griskevicius,2014)。但这也意味着,在之后的研究中,研究者们可以尝试探索两种死亡启动所产生的心理机制的异同,尝试对两种理论进行整合并互相完善。
6.3 本研究的局限及未来研究展望
由于本实验中男女被试数量差异较大,男性被试数量较少,因此虽然在有关资源的 t检验中得到了显著的性别差异,但男女被试数量的不均衡可能会影响到本实验的结果,且使得本研究无法进一步分析童年压力和当前环境压力线索在男女两性对异性的择偶要求上的影响作用是否相同。例如,前人研究显示虽然男女两性都偏好有资源的异性(Sefcek et al.,2006),但女性比男性更看重异性所拥有的金钱或资源获取途径(Buss,2007),因此可以预测同是童年压力较大的被试中,男性在受到当前环境压力线索时对异性资源的择偶要求比女性下降得更多。同样,在恋爱状态上,本研究的单身与非单身的被试数量也差距较大,非单身的被试数量较少。因此未来的研究应尝试均衡男女被试的数量及单身和非单身被试的数量,以便对这一问题进行进一步的分析。并且,本研究被试基本都是刚步入大学的大一学生,对择偶等问题尚没有较为成熟的考虑。因此在未来研究中,也可以从社会上选取年龄及童年背景更为宽泛的被试,以获得更具代表性或更为显著的结果。
其次,有学者提出,童年环境中的严峻性和不可预测性是通过影响家庭环境及亲子关系对孩子生命史策略的发展产生影响的,即家庭环境及亲子关系是其中的中介变量(Belsky et al.,1991,2012;Chen &Chang,2012)。环境的严峻性,包括低收入等,会削弱父母对孩子的温暖和敏感性,而这种疏离会进一步影响孩子的健康以及繁衍策略(Worthman&Kuzara,2005)。而环境的不可预测性,包括父母婚姻、工作及住所等的变动,也与低质量的父母教养和孩子的问题行为等相关(Belsky et al.,2012)。因此,未来的研究中也可将父母的教养方式或者亲子关系纳入对个体童年压力的测量,以使童年压力的测量维度更为全面。
此外,本研究选用了描述近期随机暴力犯罪事件和死亡率上升的新闻文章作为当前环境压力的线索,来启动死亡威胁。在以往有关生命史策略的研究中,也有一些研究者采用过描述经济衰退的新闻文章来启动当前环境中资源的稀缺(Griskevicius et al.,2013;Hill et al.,2012;White et al.,2013);且有研究显示,在死亡威胁和资源稀缺的启动下,女性对异性的不同择偶要求的偏重有所不同,在死亡威胁下,女性认为与好基因有关的特征优先于与好父母有关的特征,而在资源稀缺的启动下,女性的偏重则恰好相反(Lee &Zietsch,2011)。因此,未来的研究也可尝试采用其他的当前环境压力的启动材料(如启动资源稀缺),以检验不同的当前环境压力与童年压力在异性择偶要求上的交互作用是否不同。
Barber,N.(1995).The evolutionary psychology of physical attractiveness:Sexual selection and human morphology.Ethology and Sociobiology,16
,395−424.Belsky,J.,Schlomer,G.L.,&Ellis,B.J.(2012).Beyond cumulative risk:Distinguishing harshness and unpredictability as determinants of parenting and early life history strategy.Developmental Psychology,48
,662−673.Belsky,J.,Steinberg,L.,&Draper,P.(1991).Childhood experience,interpersonal development,and reproductive strategy:An evolutionary theory of socialization.Child
Development,62
,647−670.Buss,D.M.(1989).Sex differences in human mate preferences:Evolutionary hypotheses tested in 37 cultures.Behavioral and Brain Sciences,12
,1−14.Buss,D.M.(1997).Human social motivation in evolutionary perspective: Grounding terror management theory.Psychological Inquiry,8
,22−26.Buss,D.M.(2007).The evolution of human mating.Acta Psychologica Sinica,39
,502−512.Chen,B.B.(2016).Research on three intimate relationships:Kinship,friendship,and mateship in an evolutionary perspective
.Shanghai,China:Fudan University Press.[陈斌斌.(2016).三情研究论稿:进化心理学视角下的亲情、友情和爱情
.上海:复旦大学出版社.]Chen,B.B.,&Chang,L.(2012).Adaptive insecure attachment and resource control strategies during middle childhood.International Journal of Behavioral Development,36
,389−397.Chen,B.B.,&Chang,L.(2015).Creativity and aggression as ornament and armament:Intersexual and intrasexual selection on men’s mating behaviors.Evolutionary Psychology,13
,266−282.Chen,B.B.,&Chang,L.(2016).Procrastination as a fast life history strategy.Evolutionary Psychology,14
,1−5.Chisholm,J.S.(1993).Death,hope,and sex:Life-history theory and the development of reproductive strategies.Current Anthropology,34
,1−24.Chisholm,J.S.(1996).The evolutionary ecology of attachment organization.Human Nature,7
,1−37.Copping,L.T.,Campbell,A.,Muncer,S.(2013).Violence,teenage pregnancy,and life history.Human Nature,24
,137−157.Del Giudice,M.,Gangestad,S.W.,&Kaplan,H.S.(2015).Life history theory and evolutionary psychology.In D.M.Buss (Ed.),The handbook of evolutionary psychology
(2nd ed.,Vol.1:Foundations,pp.88−114).New York:Wiley.Dillon,H.M.,Adair,L.E.,Wang,Z.,&Johnson,Z.(2013).Slow and steady win the race:Life history,mate value,and mate settling.Personality and Individual Differences,55
,612−618.Eagly,A.H.,&Wood,W.(1999).The origins of sex differences in human behavior.American Psychologist,54
,408−423.Ellis,B.J.(2004).Timing of pubertal maturation in girls:An integrated life history approach.Psychological Bulletin,130
,920−958.Ellis,B.J.,Figueredo,A.J.,Brumbach,B.H.,&Schlomer,G.L.(2009).Fundamental dimensions of environmental risk:The impact of harsh versus unpredictable environments on the evolution and development of life history strategies.Human Nature,20
,204−268.Evans,G.W.,Gonnella,C.,Marcynyszyn,L.A.,Gentile,L.,&Salpekar,N.(2005).The role of chaos in poverty and children’s socioemotional adjustment.Psychological Science,16
,560−565.Figueredo,A.J.,Vásquez,G.,Brumbach,B.H.,&Schneider,S.M.R.(2004).The heritability of life history strategy:The K-factor,covitality,and personality.Biodemography and Social Biology,51
,121−143.Figueredo,A.J.,Vásquez,G.,Brumbach,B.H.,Schneider,S.M.R.,Sefcek,J.A.,Tal,I.R.,… Jacobs,W.J.(2006).Consilience and life history theory:From gene to brain to reproductive strategy.Developmental Review,26
,243−275.Geary,D.C.(2000).Evolution and proximate expression of human paternal investment.Psychological Bulletin,126
,55−77.Greenberg,J.,Pyszczynski,T.,&Solomon,S.(1986).The causes and consequences of a need for self-esteem:A terror management theory.InPublic self and private self
(pp.189−212).Springer New York.Griskevicius,V.,Ackerman,J.M.,Cantú,S.M.,Delton,A.W.,Robertson,T.E.,Simpson,J.A.,… Tybur,J.M.(2013).When the economy falters,do people spend or save?Responses to resource scarcity depend on childhood environment.Psychological Science,24
,197−205.Griskevicius,V.,Delton,A.W.,Robertson,T.E.,&Tybur,J.M.(2011).Environmental contingency in life history strategies:The influence of mortality and socioeconomic status on reproductive timing.Journal of Personality and Social Psychology,100
,241−254.Griskevicius,V.,Tybur,J.M.,Delton,A.W.,&Robertson,T.E.(2011).The influence of mortality and socioeconomic status on risk and delayed rewards:A life history theory approach.Journal of Personality and Social Psychology,100
,1015−1026.Hill,S.E.,Rodeheffer,C.D.,DelPriore,D.J.,&Butterfield,M.E.(2013).Ecological contingencies in women’s calorie regulation psychology:A life history approach.Journal of Experimental Social Psychology,49
,888−897.Hill,S.E.,Rodeheffer,C.D.,Griskevicius,V.,Durante,K.,&White,A.E.(2012).Boosting beauty in an economic decline:Mating,spending,and the lipstick effect.Journal of Personality and Social Psychology,103
,275−291.Janssens,K.,Pandelaere,M.,van den Bergh,B.,Millet,K.,Lens,I.,&Roe,K.(2011).Can buy me love:Mate attraction goals lead to perceptual readiness for status products.Journal of Experimental Psychology,47
,254−258.Kaplan,H.S.,&Gangestad,S.W.(2005).Life history theory and evolutionary psychology.In D.M.Buss (Ed.),The handbook of evolutionary psychology
(pp.68−95).New York:Wiley.Krupp,D.B.(2012).Marital,reproductive,and educational behaviors covary with life expectancy.Archives of Sexual Behavior,41
,1409−1414.Laran,J.,&Salerno,A.(2012).Life-history strategy,food choice,and caloric consumption.Psychological Science,24
,167−173.Lee,A.J.,&Zietsch,B.P.(2011).Experimental evidence that women’s mate preferences are directly influenced by cues of pathogen prevalence and resource scarcity.Biology Letters,7
,892−895.Li,N.P.,Bailey,J.M.,Kenrick,D.T.,&Linsenmeier,J.A.W.(2002).The necessities and luxuries of mate preferences:Testing the tradeoffs.Journal of Personality and Social Psychology,82
,947−955.Lu,H.J.,Zhu,X.Q.,&Chang,L.(2015).Good genes,good providers,and good fathers:Economic development involved in how women select a mate.Evolutionary Behavioral Sciences,9
,215−228.
Mittal,C.,&Griskevicius,V.(2014).Sense of control under uncertainty depends on people’s childhood environment:A life history theory approach.Journal of Personality and Social Psychology,107
,621−637.Nedelec,J.L.,&Beaver,K.M.(2014).Physical attractiveness as a phenotypic marker of health:An assessment using a nationally representative sample of American adults.Evolution and Human Behavior,35
,456−463.Nettle,D.(2010).Dying young and living fast:Variation in life history across English neighborhoods.Behavioral Ecology,21
,387−395.Nettle,D.(2011).Flexibility in reproductive timing in human females:Integrating ultimate and proximate explanations.Philosophical Transactions of the Royal Society B:Biological Science,366
,357−365.Promislow,D.E.L.,&Harvey,P.H.(1990).Living fast and dying young:A comparative analysis of life-history variation among mammals.Journal of Zoology,220
,417−437.Roney,J.R.(2003).Effects of visual exposure to the opposite sex:Cognitive aspects of mate attraction in human males.Personality and Social Psychology Bulletin,29
,393−404.Rosenblatt,A.,Greenberg,J.,Solomon,S.,Pyszczynski,T.,&Lyon,D.(1989).Evidence for terror management theory:I.The effects of mortality salience on reactions to those who violate or uphold cultural values.Journal of Personality and Social Psychology,57
,681−690.Sefcek,J.A.,Brumbach,B.H.,Vasquez,G.,&Miller,G.F.(2006).The evolutionary psychology of human mate choice:How ecology,genes,fertility,and fashion influence mating strategy.Journal of Psychology and Human Sexuality,18
,125−182.Shackelford,T.K.,Schmitt,D.P.,&Buss,D.M.(2005).Universal dimensions of human mate preferences.Personality and Individual Differences,39
,447−458.Simpson,J.A.,Griskevicius,V.,Kuo,S.I-C.,Sung,S.,&Collins,W.A.(2012).Evolution,stress,and sensitive periods:The influence of unpredictability in early versus late childhood on sex and risky behavior.Developmental Psychology,48
,674−686.Spielmann,S.S.,MacDonald,G.,Maxwell,J.A.,Joel,S.Peragine,D.,Muise,A.,&Impett,E.A.(2013).Settling for less out of fear of being single.Journal of Personality and Social Psychology,105
,1049−1073.Stearns,S.C.(1992).The evolution of life histories
.Oxford:Oxford University Press.White,A.E.,Li,Y.J.,Griskevicius,V.,Neuberg,S.L.,&Kenrick,D.T.(2013).Putting all your eggs in one basket:Life-history strategies,bet hedging,and diversification.Psychological Science,24
,715−722.Worthman,C.M.,&Kuzara,J.(2005).Life history and the early origins of health differentials.American Journal of Human Biology,17
,95−112.Yong,J.C.,&Li,N.P.(2012).Cash in hand,want better looking mate:Significant resource cues raise men’s mating standards.Personality and Individual Differences,53
,55−58.