广西火桐自然种群和迁地保护种群的遗传多样性比较
2015-12-26骆文华代文娟胡兴华李祥军黄仕训
骆文华,代文娟,刘 建 ,胡兴华,李祥军,3,黄仕训
( 1. 中国科学院 广西植物研究所,广西 桂林 541006;2. 广西壮族自治区林业厅,广西 南宁 530028;3. 广西师范大学 生命科学学院,广西 桂林 541004 )
广西火桐自然种群和迁地保护种群的遗传多样性比较
骆文华1,代文娟1,刘 建2,胡兴华1,李祥军1,3,黄仕训1
( 1. 中国科学院 广西植物研究所,广西 桂林 541006;2. 广西壮族自治区林业厅,广西 南宁 530028;3. 广西师范大学 生命科学学院,广西 桂林 541004 )
采用 ISSR 标记方法,对广西火桐Erythropsis kwangsiensis 10个自然种群和1个迁地保护种群的遗传多样性和遗传结构进行了分析和比较。结果表明:用10个引物从广西火桐总DNA 中共扩增出60条带,其中多态性条带51 条。自然种群中以维新种群的遗传多样性最高,而三叠岭瀑布种群最低。自然种群的总遗传变异Ht为0.281 5,种群内遗传变异HS为0.076 4,种群间遗传变异为0.205 1,基因流为0.183 2。迁地保护种群的HEt和HES均为0.118 8。自然种群间存在一定程度的遗传分化(Gst= 0.731 9),种群间变异占主导地位。合并后的自然种群的多态性条带百分率(RPPB)、Nei’s多样性指数(H)和Shannon信息指数(I)均高于迁地保护种群,分别为85.00%和31.67%、0.284 1和0.118 8、0.428 5和0.175 7。研究结果表明:广西火桐迁地保护种群的遗传多样性较低,未能涵盖整个种群。建议从多个自然种群中收集种子,尽可能多的采集样本,分区育苗种植,以增加遗传多样性水平。
广西火桐;野生种群;迁地保护种群;ISSR;遗传多样性;遗传结构
广西火桐Erythropsis kwangsiensis为梧桐科火桐属植物,为优良工艺用材和珍贵的园林观赏树种,仅分布于广西中部至南部石灰岩地区的来宾、大新、靖西、那坡、田阳等县。由于广西火桐自身生物学特性和自然、人为因素的影响,在其分布区一般呈零星分布,个体数量少,稀有大的种群分布,已处于濒危状态,被列为国家二级重点保护野生植物[1]。目前对广西火桐的研究主要集中在其资源分布状况[2]、群落特征[3])、光合特性[4]和育苗技术[5-7]等方面,而遗传多样性方面的研究尚未见报道。
为保护濒危物种广西火桐的种质资源,广西植物研究所的科研人员从2002年开始开展广西火桐的迁地保护工作,繁育定植了100多株广西火桐于桂林植物园,现部分植株已开花结实,但结实量极少且幼果早落而未能产生成熟种子。迁地保护种群涵盖其野生种群的遗传多样性水平是评价迁地保护有效性之关键[8],为此,本项研究以广西火桐10个自然种群和1个迁地保护种群为研究对象,采用简单序列重复区间(inter-simple sequence repeat, ISSR)分子标记技术来揭示广西火桐自然种群及迁地保护种群的遗传多样性和遗传结构状况,以期为制定该物种的有效保护措施提供科学依据。
1 材料与方法
1.1 材 料
于2012年8月对分布于广西来宾、大新、靖西、那坡、田阳等县的10个广西火桐自然种群及桂林植物园1个迁地保护种群选取实验材料,各种群的概况见表1。共取样株95株,其中自然种群77株,迁地保护种群18株。于每个植株上取当年生叶片,硅胶中保存备用。
用于ISSR-PCR反应的Buffer、MgCl2、dNTPs、TaqDNA聚合酶等均购于上海生物工程有限公司,ISSR引物也由上海生物工程有限公司合成。
1.2 方 法
1.2.1 DNA的提取
采用植物基因组DNA提取试剂盒(离心柱型)(上海捷瑞生物工程有限公司)进行DNA的提取,操作步骤参照说明。
1.2.2 反应体系和扩增程序
采用优化扩增反应体系:25 μL体系中1×PCR buffer,2.5 mmol/L MgCl2,0.15 mmol/L dNTPs,0.06 U/μL Taq DNA 聚合酶,3 ng/μLDNA 模板,0.2 μmol/L引物。最佳扩增程序为:94 ℃预变性 5 min;94 ℃变性 45 s,52 ℃退火 45 s,72 ℃延伸1.5 min,共30 个循环;72 ℃最后延伸7 min。
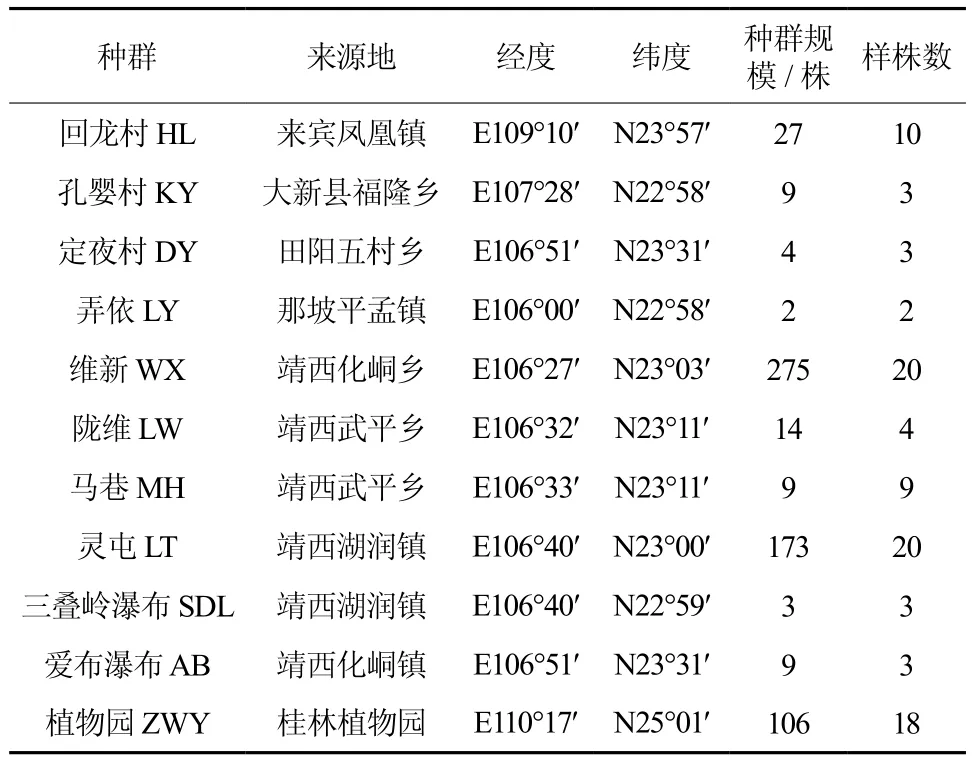
表1 供试11个广西火桐种群概况Table 1 General situation of eleven populations of E.kwangsiensis
1.2.3 扩增产物检测
PCR 扩增在Biometra TProfessional PCR仪(华粤企业集团有限公司)上进行。反应结束后,10 μL上样液含1 μL GelGreen核酸染料在1.5 %琼脂糖凝胶中电泳60 min,电压设定为120 V,于生物电泳图像分析系统(美国UVP)成像分析系统拍照记录。
1.2.4 统计分析
对扩增产物稳定的10 个引物的清晰条带进行统计分析。按条带有或无分别赋值,有带记为1,无带记为0,形成1、0数据矩阵。利用 Popgene 32 软件分析各个居群的等位基因数(Na)、有效等位基因数(Ne)、多态位点百分率。在物种水平上整个种群的遗传变异(Ht)、种群内遗传多样性(Hs)、遗传分化系数(Gst),以及Shannon多样性指数(I)和Nei’s遗传一致度(H)。采用NTSYS-pc 2.1软件对种群间遗传距离进行UPGMA(unweighedpair-group method with arithmetic-means)聚类分析,绘制聚类图。
2 结 果
2.1 种群遗传多样性
从100 条ISSR引物中筛选出电泳谱带清晰且多态性较好的10 条引物(表2),并对95个个体进行ISSR分析,共扩出60 条带,范围在400~2 000 bp(图1),其中多态性条带51条,平均每个引物扩增出的条带数为6条,平均每个引物扩增出的多态性条带数为5.1条,多态条带百分比为85.00%。
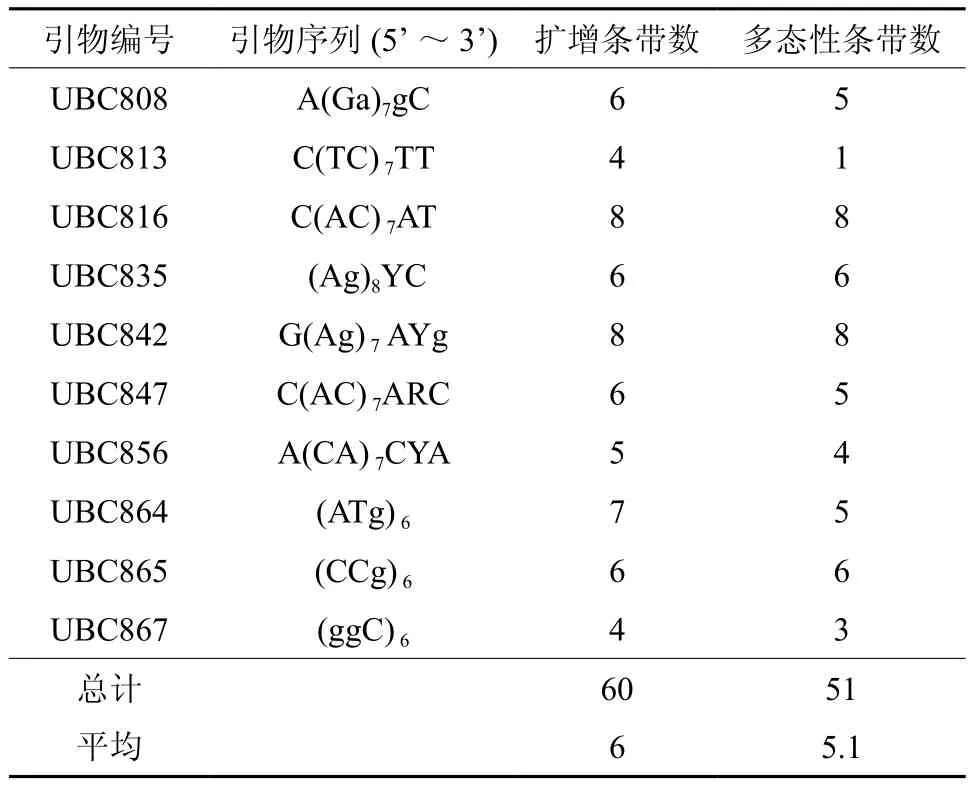
表2 10条ISSR引物序列及扩增结果Table 2 Ten ISSR primer sequences and amplification results
用Popgen32软件对11个种群的遗传多样性进行分析(表3):各种群的等位基因数(Na)范围为1.033 3~1.416 7,平均值1.207 6;有效等位基因数(Ne)从1.014 3~1.286 6,平均值1.140 9;多态性条带百分率(PPB)、Nei’s多样性指数(H)和Shannon信息指数(I)范围分别在3.33%~41.67%、0.010 0~0.157 3和0.015 9~0.231 8之间,平均值分别为20.76%、0.080 3和0.118 0。在物种水平上,广西火桐的多态性条带百分率(PPB)、Nei’s多样性指数(H)和Shannon信息指数(I)分别分别为85.00%、0.273 2和0.415 5。
图1 UBC842扩增DNA样品的电泳Fig.1 Electrophoresis of DNA samples with primer UBC842

表3 广西火桐13个种群的遗传多样性比较Table 3 Genetic diversity comparisons among 13 populations of E. kwangsiensis
自然种群中,遗传多样性较高的有维新(WX)和灵屯(LT)种群。其中维新(WX)种群的最高,多态性条带百分率(PPB)、Nei’s 遗传多样性指数(H) 和 Shannon信息指数(I)值分别为41.67%、0.156 9和0.228 5;回龙(HL)、陇维(LW)和马巷(MH)种群的PPB、H和I值处于中间;孔婴村(KY)、爱布瀑布(AB)、定夜村(DY)、弄依(LY)4个种群的遗传多样较低;三叠岭瀑布(SDL)最低,其PPB、H和I分别为3.33%、0.010 0和0.015 9。
迁地保护种群(ZWY)的等位基因数(Na)、有效等位基因数(Ne)、Nei’s多样性指数(H)和Shannon信息指数分别为1.316 7、1.206 8、0.118 8和0.175 7,多态性条带百分率(PPB)为31.67%,介于自然种群中的灵屯(LT)与回龙(HL)之间。
2.2 合并后的种群遗传多样性和遗传结构
将自然种群(定夜村DY、维新WX、弄依LY、陇维LW、马巷MH、灵屯 LT、三叠岭瀑布SDL、爱布瀑布AB、回龙村HL和孔婴村KY)合并作为 1 个组( 77个单株),桂林植物园ZWY种群(18个单株)单独作为一个组。合并后的广西火桐自然种群、迁地保护种群的遗传多样性分析结果和遗传分化结构见表4。
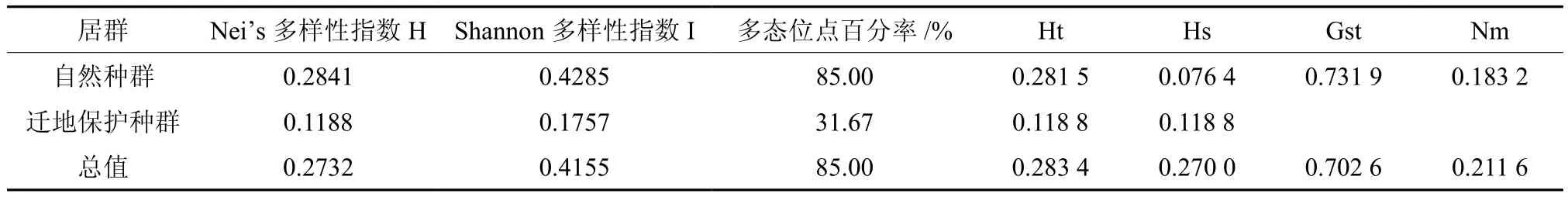
表4 广西火桐2个组的遗传多样性及结构分析Table 4 Genetic diversity and genetic differentiation parameters between 2 groups of E. kwangsiensis
表4显示:合并后,自然种群的PPB、H和I分别为85.00%、0.284 1和0.428 5;迁地保护种群的PPB、H和I分别为31.67%、0.118 8和0.175 7;自然种群的遗传多样性高于迁地保护种群。
自然种群的总遗传变异Ht为0.281 5,种群内遗传变异Hs为 0.076 4,种群间遗传变异为0.205 1,种群间遗传分化系数为0.731 9。在总的遗传多样性中,73.19%的遗传变异存在于种群间,26.81%的遗传变异存在于种群内。基因流为0.183 2。迁地保护种群的Ht和Hs均为0.118 8。
合并后,2个组的总遗传变异Ht为 0.283 4,种群内遗传变异Hs为 0.270 0,种群间遗传变异为0.013 4,种群间遗传分化系数Gst为0.702 6。在总的遗传多样性中,70.26%的遗传变异存在于种群间,29.74%的遗传变异存在于种群内,遗传变异主要存在于种群间。基因流为0.211 6。
2.3 种群遗传聚类
广西火桐11个种群的遗传一致度为0.622 0~0.940 3,遗传距离D为0.061 6~0.474 9(表5)。其中弄依(LY)和陇维(LW)之间的遗传距离最大,为0.474 9,表明两者之间的遗传差异性最大,且其相似度也最低,为0.622 0。回龙(HL)和植物园(ZWY)之间的遗传距离最近,为0.061 6,同时相似度也是最高的,为0.940 3,说明其亲缘关系最近。
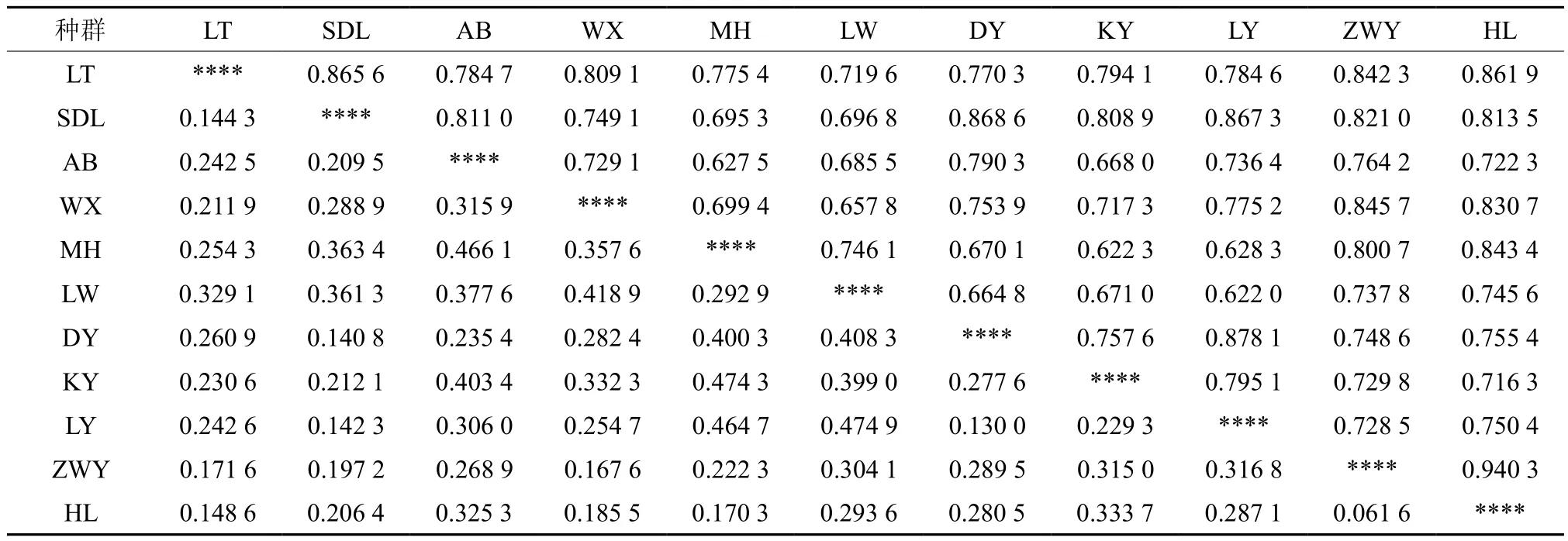
表5 广西火桐11个种群遗传一致度(对角线以上)和遗传距离D(对角线以下)Table 5 Genetic similarity coefficients (above diagonal) and genetic distance D(below diagonal) of E. kwangsiensis
基于11个种群间遗传距离的UPGMA聚类结果显示(图2),10个自然种群中靖西的灵屯(LT)先和来宾的回龙(HL)聚在一起,再与靖西的维新(WX)、马巷(MH)聚为一组;田阳的定夜村(DY)和那坡的弄依(LY)先聚在一起,再和靖西的三叠岭瀑布(SDL)、大新的孔婴村(KY)、靖西的爱布瀑布(AB)聚为一组;靖西的陇维(LW)单独聚为一组。可见,10个自然种群的亲缘关系与其地理距离之间的相关性不强。而植物园迁地保护种群(ZWY)是先与回龙(HL)聚在一起,再与灵屯(LT)相聚,表明迁地保护种群与这两个种群的亲缘关系较密切。
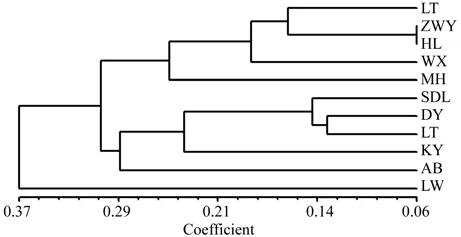
图2 广西火桐13个居群的UPGMA聚类Fig.2 UPGMA dendrogram of 13 pupulations of Erythropsis kwangsiensis
3 讨 论
3.1 广西火桐种群的遗传多样性
物种水平上,广西火桐自然种群维持较高的遗传多样性(PPB、Nei’s多样性指数H和Shannon信息指数I分别为85.00%、0.273 2和0.415 5)。种群水平的遗传多样性则较低(PPB、Nei’s多样性指数H和Shannon信息指数I分别为20.76%、0.080 3和0.118 0)。其中,自然种群的维新、灵屯和回龙种群的遗传多样性较高,而其他种群的遗传多样较低。由于种群间的遗传多样性不平衡,部分种群较低的遗传多样性使得整个种群的遗传多样性水平降低。根据野外调查,维新、灵屯和回龙种群的个体数量较多,而其他种群的个体数量较少,这表明种群遗传多样性水平与种群大小相关,较大的种群趋于具有更高的遗传多样性水平,这与刘国彬对锥栗的研究结果一致[9]。
广西火桐迁地保护种群的遗传多样性(PPB为31.67%、H为0.118 8、I为0.175 7)低于自然种群的物种水平,遗传聚类分析表明其与来宾的回龙(HL)和灵屯(LT)聚为一组。这主要与迁地保护种群的种源有关,由于当时引种时,尚未以遗传学理论为指导,且自然种群中的结实母株数量极少,当年进行种子采集时仅灵屯和回龙两个种群有母株结实,故迁地保护种群的种苗均来自于这两个种群,从其遗传多样性研究结果来看亦只基本涵盖了这两个自然种群。
3.2 广西火桐种群的遗传结构与分化
植物居群的遗传结构反映了种的长期进化史、分布区转移、生境化片断和居群特化、突变、遗传漂变、交配系统、基因流和选择等不同过程的相互作用[10]。广西火桐自然种群间的遗传分化系数较高(Gst=0.731 9),表明广西火桐自然种群间存在相当程度的遗传分化,主要遗传变异存在于种群间。自然种群的基因流较低(Nm=0.183 2),意味着种群间基因流动程度较低,种群间存在一定程度的遗传漂变。Hamrick等[11]认为,当Nm<1时,遗传漂变可以导致种群间明显的遗传分化。广西火桐种群间极低的基因流可能是导致较高遗传分化的主要原因:残存的广西火桐种群在广西石灰岩地区呈岛屿状零星分布,人为干扰严重,生境极度破碎化。此外,广西火桐自然种群中的成龄结实母树数量极少,结实量小,野外幼苗成活率低,导致有效种群的不断减少,从而限制了种群间的基因交流,增加了种群间的遗传分化。
3.3 广西火桐迁地保护建议
植物园是迁地保护稀有濒危植物最主要的场所,据统计,约有30%的受威胁植物已保存在全球1 800 个主要的植物园。我国植物园约有200个,占世界植物园的1/10,引种保存了中国植物区系的植物约20 000种,其中第一批国家重点保护植物已有90%(约350种)被引种保存[8]。迁地保护的核心问题是物种遗传完整性、种群生存力及迁地种群遗传管理[12]。就某一个种来说,其遗传变异水平关系到种群的适应能力乃至决定着整个种的进化与生存[13-14]。因此,对于广西火桐这类分布区狭窄、野生数量稀少的濒危物种,从进化与保育的角度考虑更应该充分重视其遗传多样性保护。本研究中,广西火桐迁地保护种群的遗传多样性低于自然种群,未能涵盖整个种群,与引种材料来源于少量自然种群有关。因此,为更好地保护这一珍稀濒危植物,还应进一步加强该物种的种质资源收集工作,尽可能多地从尚未进行过收集的各自然种群中采集繁殖材料,尤其是遗传多样性最高的维新(WX)种群。采集时要应用“多基因库采样法”(multiplegenepool sampling)[15],并分区育苗种植,以增加其遗传多样性水平。
[1] 国家林业总局,农业部.国家重点保护野生植物名录(第一批)[J].植物杂志, 1999(5): 4-11.
[2] 刘 演,宁世江.广西重点保护野生植物资源的现状与评价[J].广西科学, 2002, 9(2): 124-132.
[3] 骆文华,毛世忠,丁 莉,等. 濒危植物广西火桐群落特征研究[J].福建林业科技,2010,37(4):6-10.
[4] 毛世忠,唐文秀,骆文华,等. 濒危植物广西火桐净光合速率及其影响因子研究[J].广西农业科学,2010,41(11):6-10.
[5] 黄汉精,陆仕念,韦兰英.广西火桐采种与育苗技术[J]广西林业, 2008(1): 39-401.
[6] 骆文华,毛世忠,丁 莉,等. 濒危植物广西火桐种子繁殖技术及幼苗生长节律[J]福建林学院学报,2011,31(1):48-51.
[7] 付传明,黄宁珍,骆文华,等. 濒危植物广西火桐的组织培养和植株再生[J] 植物生理学通讯,2010,46(12):1253-1254.
[8] 许再富,黄加元,胡华斌,等.我国近30年来植物迁地保护及其研究的综述[J]广西植物,2008,28(6):764-774.
[9] 刘国彬.锥栗自然居群及农家品种遗传多样性的ISSR分析.硕士学位论文,华中农业大学,2009.
[10] Sehaal B A, Hayworth D A,olsenKM, Rauseher J T, Smith W A.Phylogeographic studies in Plants : Problems and prospects[J].Mol Ecol,1998,7:465-474.
[11] Hamrick J L ,Godt M J W , Sherman-Broyes S L. Gene flow among plant population:evidence from genetic markers. In:Hoch D C, Stephnon A G . Experimental and Molecular Approaches to Plant Biosystematics[J]. Missouri Botanical Garden, 1995,215-232
[12] 黄宏文,张 征.中国植物引种栽培及迁地保护的现状与展望[J].生物多样性, 2012,20:559-571.
[13] Frankham R,Ballon J D,Briscoe D A.(2002) Introduction to Conservation Genetics.Cambridge University Press
[14] Stefenon V M, Gailing O, Finkeldey R . Genetic structure of plantations and the conservation of genetic resources of Brazilian pine (Araucaria angustifolia)[J]. Forest Ecology and Management, 2008, 255:2718-2725.
[15] 许再富.稀有濒危植物迁地保护的原理与方法[M].昆明:云南科技出版社,1998.
Comparison of genetic diversity of natural and ex-situ conservation populations of Erythropsis kwangsiensis
LUO Wen-hua1, DAI Wen-juan1, LIU Jian2, HU Xing-hua1, LI Xiang-jun1,3, HUANG Shi-xun1
(1. Guangxi Institute of Botany, China Academy of Sciences, Guilin 541006, Guangxi, China; 2.Forestry Dept. of Guangxi, Nanning 530028, Guangxi, China; 3. College of Life Sciences, Guangxi Normal University; Guilin 541004, Guangxi, China)
Genetic diversity and structure of ten natural populations and one ex-situ conservation population of Erythropsis kwangsiensis were analyzed and compared by ISSR marker method. The results show that Sixty bands were amplif i ed from total genomic DNA with 10 primers,in which there were 51 polymorphic bands; Of the ten natural populations, the percentages of polymorphic band (PPB),Nei’s diversity index (h) and Shannon information index (I) of Weixin population were the highest, and those of Sandieling waterfall were the lowest; The total genetic differentiation (Ht) of natural populations was 0.281 5, with HS=0.076 4 in populations, 0.205 1 among populations, and the gene fl ow was 0.183 2; However, the genetic differentiation values (HEtand HES) both were 0.118 8 in ex-situ conservation population; There existed a genetic differentiation (Gst= 0.731 9) among natural populations but the differentiation between populations occupied an leading position. The genetic diversity values of the natural population after merged, such as the percentage of polymorphic bands (RPPB), Nei’s diversity index (H) and Shannon information index (I) were all higher than that of ex-situ conservation population(REPPB), (HE), (IE) their were 85.00% (RPPB) and 31.67%(REPPB), 0.284 1 (H) and 0.118 8 (HE), 0.428 5 (I) and 0.175 7 (IE)m respectively. It was concluded that the ex-situ conservation population failed to cover the entire genetic diversity of E. kwangsiensis, so the suggestions are that seeds collecting should be selected from several wild populations, collected samples as many as possible and made seedling and planting with partition method, in order to increase genetic diversity of E. kwangsiensis.
Erythropsis kwangsiensis; wild population; ex-situ conservation population; ISSR; genetic diversity; genetic structure
2014-04-25
广西自然科学基金项目(2012GXNSFAA053069);财政部科技支撑计划项目(KSCX2-YW-Z-1009)
骆文华,副研究员;E-mail:luowenhua2004@163.com
骆文华, 代文娟, 刘 建, 等. 广西火桐自然种群和迁地保护种群的遗传多样性比较[J].中南林业科技大学学报, 2015,35(2): 66-71.
S718
A
1673-923X(2015)02-0066-06
10.14067/j.cnki.1673-923x.2015.02.013
http: //qks.csuft.edu.cn
[本文编校:吴 彬]
