短梗大参的光合特性研究
2015-12-26梁文斌刘卫东聂东伶吴思政柏文富
梁文斌,刘卫东,聂东伶,吴思政,柏文富
(1.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;2.中南林业科技大学 林学院,湖南 长沙410004;3.湖南省森林植物园,湖南 长沙 410116)
短梗大参的光合特性研究
梁文斌1,刘卫东2,聂东伶3,吴思政3,柏文富3
(1.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;2.中南林业科技大学 林学院,湖南 长沙410004;3.湖南省森林植物园,湖南 长沙 410116)
以引种栽培的短梗大参为研究对象,采用LI-6400 便携式光合仪测定了短梗大参叶片的光合参数,探讨了其光合生理特性,为短梗大参引种及在园林上推广应用提供栽培基础。结果表明:短梗大参叶片净光合速率(Pn)、光合有效辐射(PAR)、大气温度(Ta)、蒸腾速率(Tr)和气孔导度(Gs)的日变化曲线均呈单峰曲线,不存在“午休”现象。相关性分析表明影响短梗大参叶片净光合速率的主要生态因子为光合有效辐射和大气温度,生理因子为气孔导度和蒸腾速率。短梗大参叶片净光合速率的年周期变化曲线呈单峰曲线,在夏季6~7月维持在较高水平,春季和秋季较低,冬季最低。短梗大参净光合速率的光响应曲线没有强光抑制现象,采用修正的直角双曲线模型拟合,估算出最大净光合速率(Pmax)为6.902 μmol·m-2s-1,光饱和点(LSP)为1 013.163 μmol·m-2s-1,光补偿点为(LCP)4.794 μmol·m-2s-1,暗呼吸速率(Rd)为 0.290 μmol·m-2s-1,表明短梗大参耐荫性强,具有较强的弱光适应能力,适合在遮阴环境下栽培。
短梗大参;光合特性;日变化;年变化;光响应曲线
短梗大参Macropanax rosthornii为五加科大参属常绿灌木或小乔木,多生长于林下湿润的荫蔽环境中,在四川、贵州、湖南、湖北、江西、福建等省均有分布[1],但分布范围狭窄,资源量小。由于短梗大参枝叶繁茂,叶形独特,树型美观,花大,花期长,因而具有较高的观赏价值,是一种可开发的优质野生观赏植物资源,可作为优良的园林观赏新树种,应用于园林绿化、盆景制作及室内观赏等。为了开发这种野生观赏植物资源,现已在湖南省长沙市开展了短梗大参引种栽培试验。目前有关短梗大参的研究主要集中于种子萌发特性[2]、扦插繁殖试验[3]及幼苗生长[4],光合作用研究仅发现有王萍等[5]关于盆栽短梗大参光合速率和叶绿素荧光参数日变化的报道。光合作用是影响植物物质积累与代谢的重要生理过程,也是分析环境因素影响植物生长和发育的重要手段[6]。短梗大参作为一种阴性植物,生长发育易受光照的影响,因而在引种栽培中短梗大参光合生理特性可作为评价其适应性的重要指标。本文以在长沙市引种栽培的短梗大参为试验材料,测定和分析其光合作用的日变化、光合作用的年变化和净光合速率的光响应,掌握短梗大参光合生理特性,为短梗大参的栽培生理及在园林上的应用提供理论依据。
1 试验地概况
试验地位于湖南省长沙市中南林业科技大学校园内,地处东经 112º59′34″,北纬 28º08′12″,属亚热带季风性湿润气候,季节变化明显,冬季寒冷,夏季炎热。雨量充沛,光照充足,年日照1 677 h。冬寒期短,无霜期年达281 d,年平均气温17.2℃,年均降水量1 361.6 mm。土壤为红壤。
2 材料与方法
2.1 试验材料
选定中南林业科技大学校园林下栽培5年的短梗大参(从湖南省永顺县引种),以树体外围中部的成熟叶片为试验材料。
2.2 研究方法
2.2.1 光合作用日变化的测定
在7月下旬采用LI-6400便携式光合分析仪。选择3株短梗大参,每株5枚叶片。光合日变化的测定选择在晴天进行,从6:00~18:00每隔2 h测定1次,共测3 d。同时测定气孔导度(Gs)、光合有效辐射(PAR)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、大气温度(AT)、空气相对湿度(RH)。
2.2.2 光合作用年变化的测定
从3月开始至12月,每月中、下旬选择3 d上午9:00~10:00测定短梗大参光合作用,共测3株,每株3枚叶片。
2.2.3 光响应曲线的测定
用LI-6400便携式光合仪于在6月中旬上午8:00~11:30之间测定叶片的净光合速率(Pn)对光强的响应曲线。测定时光强由强到弱,依次设定光合有效辐射(PAR)为1 600、1 300、1 000、700、400、100、80、60、40、20、0 µmol·m-2s-1,每个光照强度梯度下平衡3 min,测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等光合生理参数指标。在短梗大参植株中部枝条测定5枚叶片,取平均值。以PAR为横轴,Pn为纵轴绘制光合作用光响应曲线。
2.3 数据分析
2.3.1 光合参数计算
采用修正的直角双曲线模型[7]拟合Pn-PAR光响应曲线。
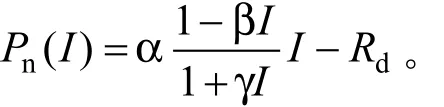
式中:Pn为净光合速率,I为光合有效辐射,α为光响应曲线的初始斜率,Rd为暗呼吸速率,β和γ为系数。通过拟合Pn-PAR 的曲线方程求解表观光量子效率(AQE)、最大净光合速率(Pmax),暗呼吸速率(Rd),光饱和点(LSP)和光补偿点(LCP)。
2.3.2 数据处理与分析
测定所获得的参数采用Microsoft Excel 2003进行数据处理,用SPSS 13.0软件进行统计分析。
3 结果与分析
3.1 短梗大参叶片净光合速率的日变化
从图1可以看出,短梗大参叶片净光合速率(Pn)日变化曲线呈单峰型,从早晨6:00开始,随着光照强度增加和气温升高,Pn逐渐升高,峰值出现在 14:00,其值为 6.62 μmol·m-2s-1,然后随着光照强度降低和温度下降,Pn逐渐下降。由于本试验中的短梗大参生长在林下,夏季高温和强光对其光合作用的影响明显减弱,因此,短梗大参Pn日变化不存在因高温和强光引起的“午休”现象。
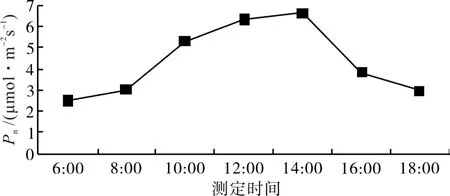
图1 短梗大参净光合速率的日变化Fig.1 Diurnal variations of net photosynthetic rate of M. rosthornii
3.2 短梗大参叶片净光合速率主要影响因子的日变化
从图2可知,短梗大参大气温度(AT)日变化为单峰型,与净光合速率(Pn)日变化趋势一致,表现为先上升后下降的趋势,早晨大气温度相对较低,随着一天中时间前移,温度逐渐上升,至中午12:00时达最高,午后开始缓慢下降;空气相对湿度(RH)的日变化与AT的日变化趋势相反,呈先下降后上升的变化趋势,在早晨6:00时最高,然后逐渐降低,至中午12:00时降至最低值,在下午随着光照强度的逐渐减弱,气温不断下降,RH逐渐回升。从图3可知,光合有效辐射(PAR)与蒸腾速率(Tr)的日变化均为单峰型,与Pn日变化趋势一致,但变化幅度大,中午的PAR和Tr分别为早晨的32.0倍和29.6倍。从图4可知,气孔导度(Gs)的日变化也是单峰型,与Pn日变化趋势一致,为先上升后下降的变化趋势,在14:00时达到峰值。胞间CO2浓度(Ci)日变化较复杂,但变化幅度不大,随着Tr的增加,Ci也随之升高,由于Pn逐渐增大,越来越多的CO2参与到光合作用中,至10:00时,Ci开始降低,在午间12:00~14:00之间降至最低,此时Pn则最高,在下午16:00后,由于Pn明显下降,Ci稍上升,但随即略下降。

图2 AT和RH的日变化Fig.2 Diurnal variations of temperature and humidity
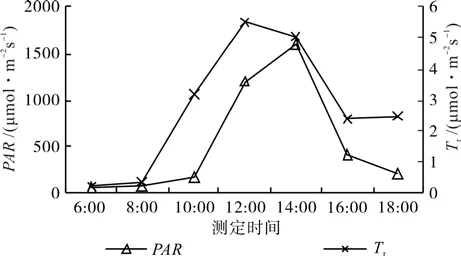
图3 PAR、Tr的日变化Fig.3 Diurnal variations of photo-synthetically active radiation and transpiration rate
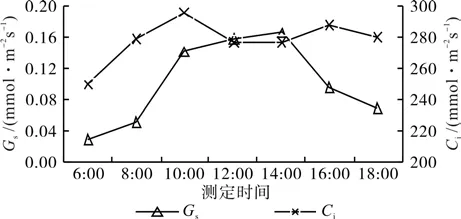
图4 Gs、Ci的日变化Fig.4 Diurnal variations of stomatal conductance and intercellular CO2 concentration
从以上分析可知,从早晨至傍晚的日进程中,随着PAR逐渐升高,AT上升,RH下降, 气孔逐渐打开,Gs也相应增加,随之Pn和Tr增加。短梗大参因处于林下环境中,因而其光合作用受高温强光的抑制弱。
3.3 短梗大参叶片净光合速率主要影响因子的相关性分析
由于短梗大参叶片净光合速率受各生理生态的综合影响,因此对所测定的结果应用相关性分析探讨了净光合速率与各影响因子的关系,结果如表1。从表1可知,RH与Pn显著负相关(P<0.05),也与其他各因子负相关;Gs、Tr与Pn极显著正相关(P<0.01),也与AT显著正相关(P<0.01);AT、PAR与Pn显著正相关(P<0.05);Ci虽然与Pn正相关,但相关度很低,只有0.379。一般来说,生态因子直接影响植物的生理活动,因而AT、PAR和RH影响Gs和Tr,进而影响短梗大参Pn。从相关性分析来看,Gs和Tr对Pn影响最大,而AT、RH对Gs、Tr的影响最大,PAR则对Gs、Tr的影响次之,也就是说在一定范围内AT和PAR越高,RH越低,则Gs和Tr越高,因而Pn就越高。从表1相关系数大小比较可知,各因子对净光合速率的综合影响程度由大到小(绝对值)的顺序为:Gs、Tr、PAR、AT、RH、Ci。由此可知,影响短梗大参叶片Pn最主要的生态因子PAR和AT,生理因子是Gs和Tr。
3.4 短梗大参叶片净光合速率年变化
树木季节性变化特征反映了其遗传特性和对环境的适应能力,研究树木光合能力的季节变化特征,有助于了解树木的生态学特性[8-9]。从图5可以看出,短梗大参叶片Pn年周期变化呈单峰曲线,季节的变化通过影响AT和PAR进而影响Pn。在3月前由于AT低和PAR弱,因而Pn低,随着春天AT上升和PAR增强,Pn逐渐升高,在夏季6~7月达最高,在秋季由于AT下降和PAR降低,Pn则逐渐下降,在冬季短梗大参仍有弱的光合能力,至12月Pn降至3月水平。
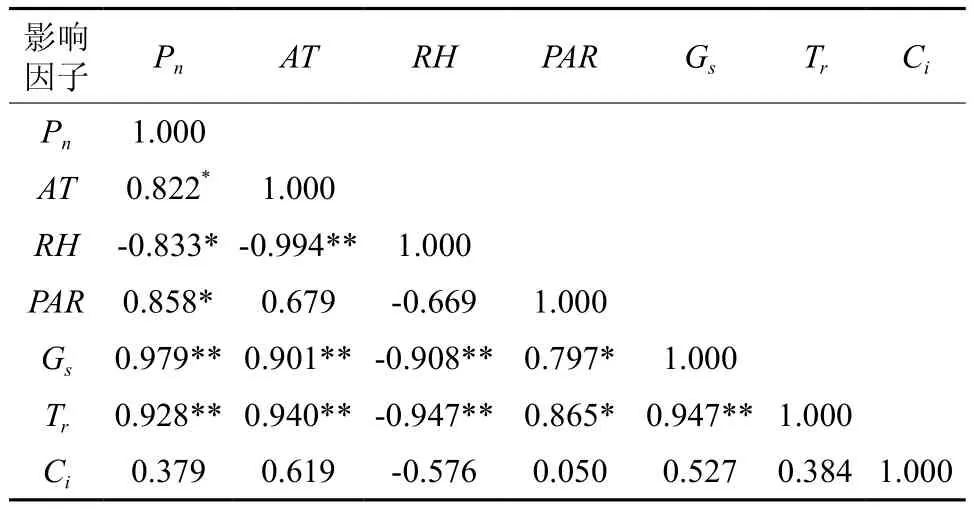
表1 短梗大参叶片净光合速率与主要生理生态因子的相关系数†Table 1 Relative correlations among photosynthetic rate and physio-ecological factors in leaves of M. rosthornii
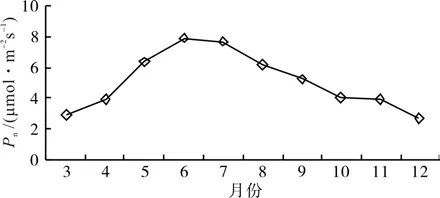
图5 短梗大参叶片净光合速率年变化Fig.5 Yearly variations of net photosynthetic rate in leaves of M. rosthornii
3.5 短梗大参叶片净光合速率的光响应
由图6可知,短梗大参光合作用的光响应曲线属于饱和趋近型[10],即在光照强度上升达到某个值后,其光合速率随光照强度进一步上升呈略微增加逐渐趋于饱和。光照强度对植物的光合作用有显著的影响,不同生态型植物对光照的响应不尽相同,其光合速率变化在强光和弱光下均有差异[11]。短梗大参光合作用的光响应过程为:当 PAR 在 0 ~ 100 μmol·m-2s-1左右时, Pn呈线性上升,且上升速度快;随着PAR继续上升,Pn的上升速度减缓;当 PAR 在400 μmol·m-2s-1后,Pn的上升处于极缓慢状态(图6),在PAR在1 000 μmol·m-2s-1后,Pn不再增加,趋于平稳,没有强光抑制现象。运用光响应曲线修正的直角双曲线模型对光响应曲线进行拟合,得到高精度拟合值,决定系数(R2)达0.998,并估算出短梗大参叶片光合速率-光响应曲线的参数值(表2)。从表2可知,短梗大参叶片最大净光合速率(Pmax)为 6.902 μmol·m-2s-1, 光 饱 和 点(LSP)1013.163 μmol·m-2s-1,光补偿点(LCP)4.794 μmol·m-2s-1,暗呼吸速率(Rd)0.290 μmol·m-2s-1。
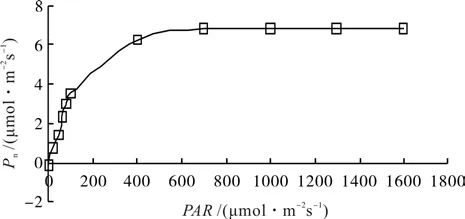
图6 短梗大参叶片净光合速率对光照强度的响应Fig.6 Responses of net photosynthetic rate to light intensity in leaves of M. rosthornii

表2 短梗大参光合速率-响应曲线的参数值Table 2 Parameters of photosynthesis-response curve of M. rosthornii
4 结论与讨论
根据目前研究,光合作用日变化主要有单峰型[12-14]、双峰型[15-18]和三峰型[19-21]三种类型。本研究中短梗大参由于栽培于林下,郁闭度较高,明显减弱了夏季高温强光的影响,叶表面接受的光合有效辐射少,往往达不到光饱和点水平,不存在光抑制的现象,因而光合速率日变化曲线呈单峰型,净光合速率的峰值出现在14:00,在其他阴性植物的光合生理特性研究中也有类似现象[22-23]。以往的研究[5]表明暴露于夏季高温强光下的盆栽短梗大参,其光合作用易受到明显的抑制,光合作用日变化类型为“双峰”型,存在明显的“午休”现象。对于同一种植物因光照环境差异导致光合作用日变化类型不同的现象在其他植物如东北铁线莲[24]、红叶桃[25]、草莓[26]也有发现,它们在全光照下均为“双峰”型,而在遮荫条件净光合速率变化趋势均为单峰曲线。
光合作用为一个复杂的生物物理和生物化学过程,主要受温度、湿度、光合有效辐射等外部生态因子和蒸腾速率、气孔导度和胞间CO2浓度等内部生理因子的综合影响,而且各种因子的关系错综复杂,在较短时期内,由于内部生理因子相对稳定,外部生态环境因子就成为主要影响因素[27],如净光合速率为双峰型的油茶“午休”发生在12:00,而不是光合有效辐射最强的14:00,表明光合作用受到温度和空气湿度的影响[28],孟丙南等[29]研究发现光照和温度是核桃光合作用的主要影响因素。影响短梗大参叶片净光合速率的生态因子有光合有效辐射、大气温度和空气相对湿度,生理因子有气孔导度、蒸腾速率和胞间CO2浓度。从各因子的相关分析来看,光合有效辐射和大气温度为影响短梗大参光合作用最主要的生态因子,而气孔导度和蒸腾速率为最主要的生理因子。光合有效辐射作为光合作用的直接能量来源,也是影响其他环境因子的最主要因素,因此光合有效辐射起主导作用,通过对大气温度和湿度的影响间接作用于净光合速率[30]。空气相对湿度是影响光合速率的一个重要生态因子,不同植物或不同环境空气相对湿度对光合速率的影响会有差异。不少研究[31-33]发现空气相对湿度高有利于提高净光合速率,但本研究中空气相对湿度与净光合速率呈负相关关系,这是因为短梗大参处于阴蔽环境中,当空气相对湿度增大时,空气蒸汽压随之增大,叶内外的蒸汽压差就会变小,蒸腾速率变慢,净光合速率也随之下降,反之则加快。王兴龙等[27]在研究阴蔽环境中生长的花榈木光合作用时也发现空气相对湿度与净光合速率呈负相关。
短梗大参叶片净光合速率在一年中不同月份中表现出明显的差异,夏季6~7月净光合速率维持在较高水平,春季和秋季较低,冬季最低。温度和光合有效辐射是影响不同季节净光合速率的主要因素,林金科[34]在茶树光合作用的研究中也证实了这个结论。
大体上阴性植物的光补偿点小于 20 μmol·m-2s-1,光饱和点为 500 ~ 1 000 μmol·m-2s-1或更低[35]。在本研究中短梗大参最大净光合速率为6.902 μmol·m-2s-1,光饱和点为 1 013.163 μmol·m-2s-1,光补偿点 4.794 μmol·m-2s-1,暗呼吸速率 0.290 μmol·m-2s-1。可以看出,短梗大参净光合速率和光饱和点均不高,光补偿点和暗呼吸速率均低,表明短梗大参是一种阴性植物,生长缓慢。另外,植物的光补偿点越低,表观光量子效率越大,则耐阴性越强[36]。本研究中短梗大参表观光量子效率为0.063,略高于自然条件下一般植物的表观光量子效率的范围(0.03~0.05)[37],说明短梗大参在较弱的光照环境中能充分利用光能,即具有较强的适应弱光的能力,这与笔者在实地调查发现野生短梗大参在林下荫蔽环境中生长良好的情况相吻合。因此,在短梗大参引种栽培中应考虑适度的荫蔽环境。
[1] 中科院中国植物志编委会.中国植物志 (第五十四卷)[M].北京:科学出版社,1978.
[2] 沈素贞,梁文斌,宋 雷.野生观赏植物短梗大参种子萌发特性的研究[J].中南林业科技大学学报,2013,33(8):57-62.
[3] 柏文富,聂东伶,吴思政,等.短梗大参扦插繁殖试验[J].湖南林业科技,2012,39(6):24-27.
[4] 陈白冰,梁文斌,聂东伶,等.不同栽培基质对短梗大参幼苗生长的影响[J].经济林研究,2011,29(3):41-44.
[5] 王 萍,聂东伶,柏文富,等.短梗大参叶片光合速率和叶绿素荧光参数日变化[J].经济林研究,2009,27(1):69-71.
[6] 贾彩凤,李艾莲.药用植物金荞麦的光合特性研究[J].中国中药杂志,2008,33(2):130-132.
[7] Z.-P.YE. A new model for relationaship between light irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica,2007,45(4):637-640.
[8] 张玉洁.香椿幼树光合作用及其影响因子研究[J].林业科学研究,2002,15(4):432-436.
[9] 宋丽丽,郭延平,徐 凯,等.温州蜜柑叶片光合作用光抑制的保护机理[J].应用生态学报,2003,14(1):47-50.
[10] 段爱国,张 建.光合作用光响应曲线模型选择及低光强属性界定[J].林业科学研究,2009,22(6):765-771.
[11] 朱万泽,王金锡,等.台湾桤木引种的光合生理特性研究[J].西北植物学报,2004,24(12):2012-2019.
[12] 张津林,张志强,查同刚,等.沙地杨树人工林生理生态特性[J].生态学报,2006,26(5):1523-1532.
[13] 李清河,刘建锋,张景波,等.乌兰布和沙漠东北部8种沙生灌木生长季末期的光合生理特性[J].西北植物学报,2006,26(11): 2318-2323.
[14] 孟德悦,吴海勇,刘光斌,等.不同种源无患子光合与蒸腾速率差异分析[J].经济林研究,2013,31(2):48-53.
[15] 彭少兵,郭军战.不同树莓和黑莓品种的光合特性研究[J].西北农林科技大学学报:自然科学版,2007,35(3):116-120.
[16] 周小玲,田大伦,许忠坤,等.中亚热带四川桤木与台湾桤木幼林的光合生态特性[J].中南林业科技大学学报,2007,27(1):40-49.
[17] 张毓涛,王文栋,李吉玫.新疆乌拉泊库区沙枣与胡杨光合特性比较[J].西北植物学报,2011,31(2):0377-0384.
[18] 郑文君,范崇辉,韩明玉.不同天气对苹果叶片光合特性的影响[J].西北农业学报,2007,16(6):124-127.
[19] 王志强,何 方,牛 良. CO2施肥对大棚油桃光合作用及产量品质的影响[J].果树学报,2001,18(2):75-79.
[20] 张 波,刘怀锋.葡萄光合作用年周期变化规律的研究[J].石河子农学院学报,1993, (4):5-8.
[21] 常美花,毕树光,张素英.加富CO2和铺设反光膜对温室桃光合特性的影响[J].西北农业学报 2007,16(4):157-160.
[22] 邓 涛,邹玲俐,王燕,等.越南抱茎茶林下光合特性研究[J].江苏农业科学,2013,41(5):143-145.
[23] 韦 霄,王满莲,蒋运生,等.显脉金花茶的光合生理特性研究[J].植物研究,2007,27(4):434-438.
[24] 王云贺,韩忠明,韩 梅,等.遮荫处理对东北铁线莲生长发育和光合特性的影响[J].生态学报,2010,30(24):6762-6770.
[25] 张斌斌,姜卫兵,翁忙玲,等.遮荫对红叶桃叶片光合生理的影响园[J].园艺学报,2010,37(8):1287-1294.
[26] 曾祥国,冯小明,向发云,等.遮阴对草莓光合特性的影响[J].湖北农业科学,2010,49(11):2811-2814.
[27] 肖笃宁,布仁仓,李秀珍.生态空间理论与景观异质性[J].生态学报,1997,17(5):453-461.
[28] 马锦林,张日清,叶 航,等.6 个油茶物种的光合特性[J].经济林研究,2012,30(4):73-76.
[29] 孟丙南,张俊佩,裴 东,等.不同砧木对核桃光合特性的影响[J].经济林研究,2013,31(2):32-37.
[30] 王兴龙,金则新,李建辉,等.花榈木光合作用日进程及其与环境因子的相关性[J].江苏农业科学,2012,40(3):143-46.
[31] 张 宇,宋敏丽,李利平.亚高温下不同空气湿度对番茄光合作用和物质积累的影响[J].生态学杂志,2012,31(2):342-347.
[32] 郭连生,田有亮.4种针叶幼树的光合生理特性与大气湿度关系的研究[J].生态学报,1994,14(2):136-141.
[33] 薛义霞,李亚灵,温祥珍.空气湿度对高温下番茄光合作用及坐果率的影响[J].园艺学报,2010,37(3):397-404.
[34] 林金科.茶树光合作用的年变化[J].福建农业大学学报,1999,28(1): 38-42.
[35] 蒋高明.植物生理生态学[M].北京:高等教育出版社,2004.
[36] 王旭军,吴际友,廖德志,等.长沙市8种常见绿化树种的耐荫性[J].东北林业大学学报,2010,38(3):14-15.
[37] 邱国雄.植物光合作用的效率//余淑文.植物生理学和分子生物学[C].北京:科学出版社:1992.236-243.
Study on photosynthetic characteristics of Macropanax rosthornii
LIANG Wen-bin1, LIU Wei-dong2, NIE Dong-ling3, WU Si-zheng3, BAI Wen-fu3
(1. School of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, Hunan, China;2. School of Forestry, Central South University of Forestry and Technology, Changsha 410004, Hunan, China;3. Hunan Forest Botanical Garden, Changsha 410116, Hunan, China)
The leaf photosynthetic parameters of the introduced Macropanax rosthornii were measured by LI-6400 portable photosynthesis system and its photosynthetic and physiological characteristics were investigated so that the results can provide a scientific guidance for its introduction and landscape application. The results showed that the diurnal course curves of the net photosynthetic rate, photosynthetically active radiation, air temperature, transpiration rate and stomatal conductance had peaks without“midday depression”; The relativity analysis indicated that the key ecological factors of photosynthetically active radiation, air temperature, the physiological factors of stomatal conductance and transpiration rate played key roles in the net photosynthetic rate, the yearly course curve of the net photosynthetic rate had a single peak, in which the net photosynthetic rate was in the peak in June and July of summer season, and in the middle in either spring or autumn and at the bottom in winter; No strong light suppression was found on the photosynthetic light response curve of M. rosthornii; The maximum net photosynthetic rate, light saturation point, light compensation point, and dark respiration rate simulated by modif i ed rectangular hyperbola were 6.902 μmol·m-2s-1,1013.163 μmol·m-2s-1,4.794 μmol·m-2s-1and 0.290 μmol·m-2s-1respectively, and thereby the fi ndings suggested that M. rosthornii had a good shade-tolerance and adaptation to weak light, so it was advisable for growing under shading conditions.
Macropanax rosthornii; photosynthetic characteristics; diurnal change; annual change; light response curve
S727.3;S687
A
1673-923X(2015)02-0006-06
10.14067/j.cnki.1673-923x.2015.02.002
http: //qks.csuft.edu.cn
2014-01-16
湖南省教育厅科学研究重点项目“野生观赏植物短梗大参引种驯化与繁育技术研究”(11A130)
梁文斌,硕士,副教授;E-mail:topeasie@163.com
梁文斌,刘卫东,聂东伶,等. 短梗大参的光合特性研究[J].中南林业科技大学学报, 2015,35(2):6-11.
[本文编校:吴 彬]
