羔羊瘤胃发育及其影响因素研究进展
2015-12-23祁敏丽刁其玉张乃锋
祁敏丽,刁其玉,张乃锋
(中国农业科学院饲料研究所,农业部饲料生物技术重点开放实验室,北京100081)
羔羊瘤胃发育及其影响因素研究进展
祁敏丽,刁其玉,张乃锋*
(中国农业科学院饲料研究所,农业部饲料生物技术重点开放实验室,北京100081)
瘤胃是反刍动物特有的消化器官,羔羊瘤胃的发育程度直接影响到其成年后的采食、消化能力及生产性能的发挥。本文根据瘤胃的发育进程,分别从非反刍阶段、过渡阶段和反刍阶段对瘤胃组织形态和生理代谢发育的研究进展做了综述,并对瘤胃发育的影响因素进行了阐述。
羔羊;瘤胃;组织形态;代谢功能
瘤胃是反刍动物特有的消化器官,成年羊的瘤胃相对质量占全胃的60%左右。瘤胃上皮是瘤胃执行吸收、代谢功能的重要组织,由瘤胃微生物产生的挥发性脂肪酸(VFA)的85%由瘤胃上皮直接吸收并可为宿主提供60%~80%的所需代谢能[1]。羔羊出生时瘤胃非常小,约占总胃重的17%[2]。羔羊从出生到以采食固体饲料为主,其瘤胃经历了由非反刍向反刍的生理功能转变。非反刍阶段瘤胃未发育完全,不具有代谢功能[3],随着饲料进入瘤胃后,瘤胃的生理代谢功能逐渐形成。羔羊瘤胃的发育程度直接影响到其成年后的采食和消化能力及生产性能的发挥。因此,本文对羔羊瘤胃形态和功能的发育及影响因素进行了综述。
1 羔羊瘤胃的发育
Wardrop等[4]和Poe等[5]根据瘤胃发育特点,将羔羊瘤胃发育分为3个阶段,分别是初生至3周龄的非反刍阶段;3~8周龄的过渡阶段和8周龄以后的反刍阶段。
1.1 非反刍阶段此阶段母乳营养充足羔羊机体发育迅速,瘤胃组织结构快速发育(如表1)。研究表明,到20日龄羔羊(波尔山羊)瘤胃重41 g[6],瘤胃相对质量在7~21日龄增速较大,瘤胃相对质量由约占全胃比例的20%增长到43%;同时瘤胃容积占全胃的比例由15%扩增到占46%[2,7]。新出生的羔羊瘤胃乳头长度为0.205 mm,宽度为0.091 mm,到15日龄瘤胃乳头长度为0.368 mm,宽度为0.129 mm,乳头变长变宽[8]。但是此阶段羔羊瘤胃乳头表面较光滑,上皮细胞相对细小扁平[9]。这主要是由于羔羊从出生到3周龄由于食管沟闭合,母乳或液体饲料直接进入真胃,对瘤胃上皮细胞没有直接刺激作用。
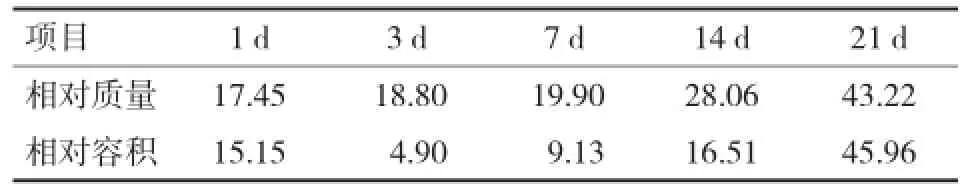
表1 放牧条件下瘤胃占全胃的相对质量和相对容积比例[2]%
早期研究发现,羔羊出生后2 d瘤胃内已有严格的厌氧微生物,数量与成年动物相当[10]。这表明瘤胃微生物区系的建立不依赖固体饲料的采食。羔羊出生后8~10日龄时其瘤胃中可出现厌氧真菌[10]。而瘤胃微生物是瘤胃功能发挥的基础,群体饲养的羔羊纤维素分解菌和产甲烷菌在出生后34日龄出现,1周后接近成年羊的水平;与母羊共同饲养的羔羊在15~20日龄可以在瘤胃内检测出原虫。20日龄的羔羊瘤胃内已经出现了瘤胃普雷沃氏菌(Prevotella ruminicola)、瘤胃壁细菌门(Firmicutes)及拟杆菌门(Bacteroidetes)的细菌[11]。瘤胃内的微生物出现的时间不一致是否与饲养模式或饲料有关有待进一步研究。羔羊在出生时瘤胃内已经检测到蛋白酶和淀粉酶,且不随日龄变化;14日龄的羔羊瘤胃内均可检出纤维素酶,随后其酶活力随日龄逐渐增加[4]。普遍认为瘤胃内的消化酶的变化由微生物产生,但是目前对瘤胃微生物优势菌群与消化酶的相关性研究的报道较少。
反映瘤胃内环境的指标主要有VFA浓度、氨态氮浓度和瘤胃pH。出生后羔羊瘤胃内的VFA浓度从无到有(图1)且存在个体差异。郭江鹏等研究发现1日龄羔羊瘤胃内没有挥发性脂肪酸(VFA),部分7日龄的羔羊瘤胃内出现VFA,21日龄所有羔羊瘤胃内均有VFA,总VFA浓度为25 mmol/L[12]。出生后3周内瘤胃内氨态氮浓度较高,14日龄时氨态氮浓度可达到25 mmol/L,此时瘤胃有较高的pH,pH接近6.8,随后降低[4]。但是,Baldwin等[13]体外研究发现,羔羊出生时瘤胃上皮氧化丁酸和葡萄糖的速度相同,随后发现瘤胃上皮氧化丁酸的能力随着年龄逐渐增加。此阶段活体瘤胃内VFA浓度低,因此丁酸供能少,瘤胃上皮可能主要利用葡萄糖氧化功能。
1.2 过渡阶段随着年龄增长羔羊采食固体饲料增多,瘤胃组织形态进一步发育,同时各项功能开始逐渐增强。此阶段羔羊瘤胃相对质量和容积进一步增加。到56日龄羔羊瘤胃占总胃重的比例达到60%[4],占总胃容积的比例达到78%,接近成年羔羊瘤胃相对质量和容积。30日龄的羔羊瘤胃乳头长度达到较高水平为1.709 mm,然后降低,到45日龄羔羊瘤胃乳头长度为0.707 mm,随后继续增长,到60日龄达到2.006 mm。瘤胃乳头宽度一直增加,从30日龄的0.276 mm增长到60日龄的0.503 mm[8]。随着日龄的增加瘤胃乳头表面角质化程度不断提高,到6~10周龄瘤胃乳头表面明显变粗糙[14]。因此,此阶段瘤胃组织形态发育主要是瘤胃基层以及瘤胃乳头的发育,其中瘤胃乳头生长是与非反刍阶段相比的最大变化。
21日龄的羔羊瘤胃内的微生物已经可以消化大部分成年羊消化利用的饲料[15]。50日龄羔羊瘤胃内优势菌群出现纤维分解菌[10]。兼性厌氧菌快速繁殖后,逐渐被厌氧微生物取代,在6-8周龄趋于稳定[16]。在2月龄内羔羊瘤胃内原虫数量一直持续增加,2个月时达(5.7±3.6)×105个/mL[10],70日龄优势菌群中出现原虫。受采食量的变化,此阶段瘤胃优势菌群不稳定,但是杆菌门和壁厚菌门一直是此阶段的优势菌[11]。羔羊瘤胃内的消化酶活力在此阶段变化不大,随日龄间差异不显著[2,12],部分日龄消化酶活力的变化可能与日粮的变化导致微生物种类与数量的变化有关。
21日龄后瘤胃内VFA浓度快速升高(如图1)。不同饲养管理条件下56日龄羔羊瘤胃内总VFA浓度在60~130 mmol/L,与成年羊的瘤胃VFA浓度相当[3]。瘤胃内氨态氮的浓度在21日龄后迅速降低,到5周龄后稳定在25 mmol/L,与成年羊瘤胃接近。瘤胃pH稳定在6.0~6.7[2],不随日龄变化[2,12]。但是瘤胃内的VFA浓度和氨态氮受瘤胃微生物产生速度和瘤胃上皮吸收速度的影响,这就表明瘤胃内环境的变化与瘤胃乳头生长和饲料的种类有关。

图1 放牧羔羊瘤胃内VFA浓度随日龄的变化[2]
新生绵羊的瘤胃上皮细胞利用葡萄糖的能力随着日龄的增长不断增加,一直持续到42日龄。随后葡萄糖的利用迅速降低,而丁酸的利用却在逐渐增加[17]。42日龄时瘤胃上皮出现生酮作用特征性的、显著的增加,42日龄以后其产生BHBA的速率和成年羊瘤胃产生的速率一致,且不随日龄变化。Lane等[18]研究发现,瘤胃上皮的3羟基3甲基辅酶A合成酶(HMGCoA合成酶)和乙酰乙酰辅酶A硫解酶(AcetoacetyCoA)的mRNA水平随日龄增加而改变,但并不随VFA的出现改变。这与Lane等[18]通过给羔羊灌注VFA,血液中β羟丁酸的浓度与VFA浓度的影响的结果一致。但是羔羊瘤胃上皮生酮基因的表达量反应羔羊瘤胃上皮生酮能力,这就有待深入研究此阶段影响羔羊瘤胃生酮能力的因素。
1.3 反刍阶段到56日龄羔羊瘤胃发育基本趋于成熟,瘤胃进入反刍阶段。此阶段全胃占总消化道的相对比例在不断增加,到112 d占全消化道的39%,成年后占49%,但是瘤胃占全胃的相对质量稳定在60%,这就表明此阶段瘤胃的发育与其他3个胃的发育速度相当。瘤胃重量随日龄逐渐增加,200日龄的小尾寒羊到可达到450 g,滩羊达到300 g[19]。瘤胃液的体积到100日龄增加趋于稳定,到150日龄增加到4.84升[8]。瘤胃乳头长度和宽度随日龄增加,但是单位面积上瘤胃乳头数量却减少,由2月龄的385个/cm减少到132.58个/cm[6],此阶段瘤胃角质化明显。瘤胃内的微生物主要包含原虫、真菌、细菌。瘤胃微生物中细菌的数量最多为1010~1011CFU/mL,其次是原虫105~106CFU/mL,真菌数量最少为103~104CFU/mL。羔羊瘤胃细菌总数量随日龄持续增加到120~135日龄后趋于稳定,瘤胃液中纤毛虫数量在75~90日龄增加迅速,在120日龄趋于稳定[8]。瘤胃内的消化酶活力在反刍阶段变化不明显,纤维素酶的酶活力较稳定,但是在9、11周龄和15周龄浓度较高[9]。α-淀粉酶的活力呈曲线变化,蛋白酶的总活性在80~200日龄间呈现逐渐上升的趋势,200日龄时增大明显,变化为0.10~0.52 IU;脂肪酶的活力呈上升的趋势[19]。此阶段瘤胃微生物区系稳定,优势菌群明显。

表2 不同日龄的小尾寒羊瘤胃发育[19]
瘤胃内的pH稳定在6.3~7.0,且不随日龄变化[8]。氨态氮浓度随日龄略有增加,在100日龄时达到稳定,达到最高值[8],但在200日龄可能会再次明显增加[19]。瘤胃内的总VFA浓度处于60~130 mmol/L,但是瘤胃内的乙酸、丙酸、丁酸的浓度及相关比例与饲喂日粮有关[5]。
综上所述,反刍阶段羔羊瘤胃绝对质量增加,但是相对于在此阶段羔羊其他消化道的发育,瘤胃组织结构发育处于稳定状态,瘤胃生理代谢功能变化较小。
2 羔羊瘤胃发育的影响因素
2.1 日粮类型与结构乳头的长度和密度随日粮精料比例和营养水平增加而增加,精料的采食量增加能够增加瘤胃基层的厚度[20],以饲喂精料为主的羔羊,瘤胃上皮颗粒细胞层厚度增加[21]。与饲喂干草的羔羊比较,饲喂精料的羔羊瘤胃重量、上皮细胞DNA含量、蛋白质合成能力高[22]。过度饲喂精料会导致乳头角质化不全,形态异常。高比例的精料会加速瘤胃微生物区系的建立,进而通过增加VFA和NH3-N的浓度来增加瘤胃代谢活性。瘤胃发酵速率过快,丙酸转化为乳酸含量增加,虽然乳酸也可进一步转化为葡萄糖,但容易造成瘤胃酸中毒,影响纤维物质等的消化率、采食量及机体健康。为此,合理的精粗比例是健康瘤胃发育的关键,但是在羔羊早期是否饲喂干草还没得到统一定论。Norouzian等[23]认为开食料中含有15%的苜蓿在不影响羔羊体重的情况下可以降低瘤胃角质层,增加瘤胃壁肌肉层厚度。
除了日粮精粗比对羔羊瘤胃有不同程度的影响外,饲料原料和日粮颗粒大小也会影响羔羊瘤胃的发育。蔡健森[24]在研究羔羊早期断奶时发现,饲喂植物性蛋白和乳源性蛋白的羔羊可增加瘤胃乳头的数量,但不增加羔羊90 d瘤胃的重量。较小颗粒的日粮可减少瘤胃乳头的生长[23]。日粮的颗粒较粗糙会降低乳头角质层厚度,减小代谢产物通过瘤胃上皮的阻力,但是Norouzian等[25]研究发现,紫花苜蓿的颗粒大小对羔羊的瘤胃乳头的长度、宽度、密度以及瘤胃上皮的厚度和面积没有影响。
2.2 日粮营养素水平孙志洪[26]报道羔羊28日龄断奶后,限制营养水平,羔羊瘤胃乳头宽度、长度和绒毛表面积明显减少。苏月菊等[27]研究表明,口服重组胰岛素生长因子(rhIGFI)粗制品促进新生羔羊瘤胃上皮乳头发育,刺激瘤胃上皮细胞分裂增加细胞数量,对单个细胞体积的影响相对较小。朱文涛等[8]研究30日龄的断奶羔羊表明日粮中高能量可增加瘤胃液内的丙酸的浓度,降低丁酸的浓度,但是对乙酸浓度影响较小,同时还会影响瘤胃液中原虫的数量。高蛋白水平可增加羔羊瘤胃液中原虫和大杆菌的数量,增加瘤胃液氨态氮浓度。能量水平对瘤胃发育的影响主要体现在对瘤胃上皮和瘤胃微生物细菌的影响,但是蛋白质水平影响瘤胃上皮的发育相关报道较少。
2.3 饲养管理放牧条件下羔羊瘤胃内的VFA浓度在56日龄前逐渐增加,到56日龄后接近成年水平并趋于稳定的状态,pH稳定在6.03~6.67[4]而舍饲条件下VFA总浓度在42 d达到最大并趋于稳定,pH稳定在5.30~6.03[7]。放牧条件下瘤胃内容物中微生物蛋白酶、A-淀粉酶和纤维素酶比活性出现在14、42、28 d[4],舍饲条件下3种酶的活性最高出现在21、7、14 d[12]。饲养模式影响瘤胃代谢参数和消化功能可能与采食的饲料种类以及数量有关。
断奶时间与断奶方式,是否饲喂固体饲料以及饲喂固体饲料的时间同样都会影响瘤胃的发育。Norouzian等[23]认为人工喂养的羔羊可增加瘤胃角质层的厚度,血液中BHBA的浓度高,瘤胃代谢能力强,但不增加瘤胃的乳头长度、宽度和密度。岳喜新[11]认为使用代乳品进行早期断奶可增加羔羊瘤胃重量和瘤胃乳头长度。饲喂代乳料的羔羊瘤胃乳头长度、宽度均要高于饲喂精料的羔羊瘤胃乳头[8]。但是新生反刍动物仅喂乳汁或代乳品,会延滞瘤胃的发育,瘤胃比同龄的正常动物小,胃壁较薄,乳头缺乏正常的发育和色泽[28],总VFA浓度较低。Lane等[29]通过瘤胃上皮细胞培养发现,饲喂固体日粮的羔羊,培养基中β-羟丁酸产量增加;延迟固体日粮饲喂时间使得瘤胃上皮细胞的代谢功能受到阻滞。因此,在生产中掌握合适的断奶时间以及补饲开食料有利于瘤胃的发育。
3 结论
综上所述,羔羊瘤胃发育一直受到学者们的高度关注。目前的主要研究进展依然集中在羔羊瘤胃的组织形态、消化代谢功能及其影响因素等方面,对于瘤胃组织形态的发育机制尤其是瘤胃上皮组织代谢的机制研究依然非常缺乏,这也是作者认为以后需要加强研究的重点。现代分子生物技术的发展已经为我们从细胞水平与基因水平上开展研究提供了许多技术手段和方法,因此,建议针对瘤胃上皮代谢机制及相关主效基因的表达,瘤胃微生物代谢及相关调控机制等方面开展研究以提高我们对羔羊瘤胃发育机制的认知水平。
[1]韩正康,陈杰.反刍动物瘤胃的消化和代谢[M].北京:科学出版社,1988.
[2]王彩莲,郝正里,李发弟,等.0~56日龄放牧羔羊消化道的特点和瘤胃功能变化[J].畜牧兽医学报,2010,41(4):417-424.
[3]Warner R G,Flatt W P,Loosli J K.Ruminant nutrition,dietary factors influencing development of ruminant stomach[J].J Agri Food Chem,1956,4(9):788-792.
[4]Wardrop I D,Coombe J B.The post-natal growth of the visceral organs of the lamb I.The growth of the visceral organs of the grazing lamb from birth to sixteen weeks of age[J].J Agric Sci, 1960,54(1):140-143.
[5]Poe S E,GIimp H A,Deweese W P,et al.Effect of preweaning diet on the growth and development of early-weaned lambs[J].J Anim Sci,1969,28(3):401-405.
[6]赵恒波.复合酶制剂对羔羊早期生长发育的影响及作用机理初探[D].北京:中国农业大学,2006.
[7]郭江鹏,张元兴,李发娣,等.0-56日龄舍饲肉用羔羊胃肠道发育特点研究[J].畜牧兽医学报,2011,42(4):513-520.
[8]朱文涛.15-150日龄羔羊日粮消化率和瘤胃消化功能变化的研究[D].新疆农业大学,2005.
[9]Z~itnan R,Bomba A,Sommer A,et al.Development of rumen metabolism and ruminal epithelium in lambs[J].Arch Anim Nutr,1993,44(3):227-233.
[10]Fonty G,Gouet P,Jouany J P,et al.Establishment of the microflora and anaerobic fungi in the rumen of lambs[J].J General Microbiol,1987,133(7):1835-1843.
[11]岳喜新.蛋白水平及饲喂量对早期断奶羔羊生长性能及消化代谢的影响[D].阿拉尔:塔里木大学,2011.
[12]郭江鹏,潘建忠,李发娣等.0-56日龄舍饲肉用羔羊前胃功能发育研究[J].草业学报,2011,20(3):128-135.
[13]Baldwin R L,Jesse B W.Developmental changes in glucose and butyrate metabolism by isolated sheep ruminal cells[J].J Nutr, 1992,122(5):1149-1153.
[14]Mears G J.The relationship of plasma somatomedin(IGF-1)to Iamb growth rate[J].Canadian J Anim Sci,1995,75:327-331.
[15]Walker D M,Walker G J.The development of the digestive system of the younganimal V.The development of rumen function in the young lamb[J].J Agric Sci,1961,57(2):271-278.
[16]Minato H,Otsuka M,Shirasaka S,et al.Colonization of microorganisms in the rumen of young calves[J].J General Appl Microbio,1992,38(5):447-456.
[18]Lane M A,Baldwin R L,Jesse B W.Developmental changes in ketogenic enzyme gene expressionduring sheep rumen development[J].J Anim Sci,2002,80(6):1538-1544.
[19]张宁.不同日龄小尾寒羊与滩羊消化器官及其胃消化酶活性的比较研究[D].杨凌:西北农林科技大学,2008.
[20]Zitnan R,Kuhla S,Nurnberg K et al.Influence of the diet on the morphology of ruminal and intestinal mucosa and on intestinal carbohydrate levels in cattle[J].Vet Med Czech,2003, 48:177-182.
[21]Graham C,Simmons N L.Functional organization of the bovine rumen epithelium[J].Am J Physiol-Regul I,2005,288(1): R173-R181.
[22]Mcleod K R,Baldwin R L.Effects of diet forage:concentrate ratio and metabolizable energy intake on visceral organ growth and in vitro oxidative capacity of gut tissues in sheep[J].J Anim Sci,2000,78:
[23]Norouzian M A,Valizadeh R,Vahmani P.Rumen development and growth of Balouchi lambs offered alfalfa hay pre-and postweaning[J].Tropical Anim Health Product,2011,43:1169-1174.
[24]蔡建森.蛋白质来源对早期断奶羔羊生产性能和器官发育及血清生化指标的而影响[D].北京:中国农业科学院,2007.
[25]Norouzian M A,Valizadeh R.Effect of forage inclusion and particle size in diets of neonatal lambs on performance and rumen development[J].J Anim Physiol Anim Nutr,2014,98(6): 1095-1101.
[26]孙志洪.关键营养素限制对早期断奶羔羊胃肠道发育程序化及营养干预研究[D].北京:中国科学院,2010.
[27]苏月菊,沈赞明,陈杰.口服rhIGF-1粗制品对新生羔羊瘤胃上皮生长发育的影响[D].北京:全国反当动物第三次学术会议,2004:45.
[28]Heinrichs J.Rumen development in the dairy calf[J].AdvancesDairy Technol,2005,17:
[29]Lane M A,Baldwin R L,Jesse BSheep rumen metabolic development in response to age and dietary treatments[J].JAnim Sci,
Advance in Ruminal Development and Its Influencing Factors in Lambs
QI Min-li,DIAO Qi-yu,ZHANG Nai-feng*
(Feed Research Institute of Chinese Academy of Agricultural Sciences,Key Laboratory of Feed Biotechnology of the Ministry of Agriculture,Beijing 100081,China)
Rumen is a particular digestive organ of the ruminant animal.The development of the lamb rumen directly affect the feed intake,digestion ability and production performance of later adults.In this paper,the research progress on tissue morphology and physiological metabolism function for lamb rumen were reviewed according to the development processes of rumen,and the influencing factors of rumen development were also discussed.
lambs;rumen;tissue morphology;physiological function
S827.2
A
0258-7033(2015)09-0077-04
2014-09-02;
2015-01-19
公益性行业(农业)科研专项(201303143)
祁敏丽(1990-),女,河北保定人,硕士研究生,主要从事幼畜营养与生理研究,E-mail:minliqi@yeah.net
*通讯作者:张乃锋,副研究员,硕士生导师,E-mail:zhangnaifeng@caas.cn
