Responses in gas exchange and water status between drought-tolerant and-susceptible soybean genotypes with ABA application
2015-12-21MdMokterHossainHonMingLamJianhuaZhang
Md.Mokter Hossain,Hon-Ming Lam,Jianhua Zhang*
School of Life Sciences and Center for Soybean Research of the State Key Laboratory of Agrobiotechnology,The Chinese University of Hong Kong,Shatin,Hong Kong,China
Responses in gas exchange and water status between drought-tolerant and-susceptible soybean genotypes with ABA application
Md.Mokter Hossain,Hon-Ming Lam,Jianhua Zhang*
School of Life Sciences and Center for Soybean Research of the State Key Laboratory of Agrobiotechnology,The Chinese University of Hong Kong,Shatin,Hong Kong,China
A R T I C L E I N F O
Article history:
12 August 2015
Accepted 3 September 2015
Available online 14 October 2015
Drought stress Exogenous ABA Leaf relative water content Stomatal conductance Soybean genotypes
The purpose ofthis study was to investigate the physiologicalresponses ofdrought-tolerantand drought-susceptible soybean genotypes to exogenous abscisic acid(ABA)application during progressive soildrying at seedling stages.Five-day old soybean seedlings were transplanted into PVCtubes filled with soilmixture.Seedlings were watered daily with similar water volumes until second trifoliate leaves emerged,and thereafter soil drying with or without exogenous ABA application was imposed.Halfofthe seedlings ofeach genotype were leftfor regular watering as control plants.Soil water status declined significantly over seven days of withholding water supply for both genotypes.Leaf expansion rate,stomatal conductance(gs),leaf water potential (ψw),and relative water contentofleaves(%RWC)declined significantly undersoildrying as well as soil drying with ABA application,compared to their values for well-watered soybean genotypes.However,a drought-tolerant genotype(C12)responded more rapidly than a drought-susceptible genotype(C08)after imposition of soil drying and soil drying with exogenous ABA.In addition,application of exogenous ABA to water-restricted soybeans resulted in higher%RWC andψwin the drought-tolerant than in the drought-susceptible genotype.Compared to the drought-susceptible genotype,the drought-tolerant genotype was more responsive to exogenous ABA application,resulting in a higher root-to-shoot ratio.
©2015 Crop Science Society of China and Institute of Crop Science,CAAS.Production and hosting by Elsevier B.V.This is an open access article under the CC BY-NC-ND license
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
1.Introduction
Droughtstress is a key environmentalconstraintto productivity in worldagriculture.Waterstress impairs numerous physiologicalas well as biochemical processes of crop plants.Photosynthesis is a major process affected by water deficit,via decreased CO2diffusion to the chloroplast and metabolic constraints[1]. Water-restricted plants show immediate low stomatal conductance without alteration of shoot water potential.The reason for this response is thatleaf stomata cannotopen because plants cansense the availability of water in the soil and regulate stomatal behavior accordingly,whatever the water status oftheir leaves[2]. The consequences of water stress to crop plants are thus a reduction in leaf expansion rate,accelerated leaf senescence,and reduction in production and partitioning of photo assimilates to the plants.
Drought tolerance is thus a key trait for increasing and stabilizing crop production[3,4].Abscisic acid(ABA)accumulates in response to low-temperature and water stress[5,6], and is now known to be involved in several developmental processes as well as acclimation to environmental stresses (cold,salt,and drought)[7,8].ABA is produced in plant roots and transported to leaves via xylem flow under water-limiting conditions[9].It triggers stomatal closure in response to drought stress,thereby reducing photosynthetic CO2assimilation[10].The influence of plant hormones such as ABA in plants under drought stress is thus important in determining physiological responses that may ultimately lead to adaptation to unfavorable environmental conditions[11].
Exogenous ABA application reduced photosynthetic rate, stomatal conductance,and transpiration rate in cotton[12].Liu et al.[13]found that exogenous ABA application decreased photosynthetic rate and pod set in well-watered soybean plants but that those parameters were increased when ABAwas applied to drought-stressed plants.Lam[14]genomically evaluated 31 soybean accessions at the Chinese University of Hong Kong for developing drought-tolerant soybean varieties,and identified promising genotypes in a hydroponic culture system using polyethylene glycol(PEG)and salts.Field performance of two drought-tolerant and susceptible genotypes(C12 and C08)has been partially tested in Dunhuang,China.
Given that drought-stressed plants produce ABA in leaves and regulate their stomata by maintaining higher water status in leaves,we hypothesized that the application of exogenous ABA to leaves of soil drying soybean plants would encourage earlier stomatal closure and reduce water loss from plants,thereby leading to higher relative water content and water potential in leaves.This study was accordingly undertaken to investigate the differences in physiological response to exogenous ABA application of drought-tolerant and drought-susceptible soybean genotypes during progressive soil drying.
2.Materials and methods
2.1.Plant materials and exogenous ABA application
The drought-tolerant Jindou 21(C12)and drought-susceptible Union(C08)soybean genotypes were used for this study. Seedlings were grown in a plastic tray containing soil mixture (soil and peat moss)in the greenhouse.Five-day old seedlings were transplanted into PVC tubes(50 cm length×5 cm inner diameter)filled with soil mixture(soil and peat moss in a 1:1 volume ratio,with the addition of NPK at 14:14:14).Fertilizer granules were mixed at 5 g L-1of soil mixture.Plants were grown under natural sunlight in the greenhouse with average daytime temperature 28±2°C and relative humidity 60-70%. Plants were watered daily with similar water volumes until second trifoliate leaves emerged,after which soil drying treatment was imposed.One third of the seedlings of each genotype were kept for regular watering as control plants, another third subjected to soil drying,and the remaining third subjected to soil drying+exogenous ABA treatment.At the time of beginning soil drying,50 mol L-1solution of ABA with 0.05%Tween-20 was sprayed on soil drying plants twice daily (at 1000 h and 1400 h)for two days on both adaxialand abaxial surface of leaves and measurement was started 3 h after the first spray.
2.2.Measurement of stomatalconductance
After the onset of soil drying along with spraying of exogenous ABA on leaves,the stomatal conductance(gs)of fully expanded leaves was measured 3 h after ABA application.Thereafter,gswas measured daily with a leaf porometer (Decagon Devices,Inc.USA)until a week of soil drying.
2.3.Measurement of leaf area expansion
Leaf area(LA)was measured with a portable leaf area meter (LI-3100;Li-COR,Inc.USA).After imposition of soil drying followed by application of exogenous ABA newly emerged leaves(center leaflet of second trifoliate leaf)were tagged for measuring LA each day.Leaf length(LL)and width(LW)were measured daily with a measuring ruler and the relationship between the product LL×LW and LA was determined for each genotype from individual leaf measurements of 15 leaves.The regression of LA on LL×LW was fitted as LA= k×LL×LW,where k is the slope of the linear function.
2.4.Measurement of relative water content
Relative water content(%RWC)of fully expanded youngest mature leaves was measured on both soil drying and soil drying+ABA-treated plants 0,1,3,and 7 days after imposition of treatments.To minimize solute leakage and cut surface effect,the entire leaf was used.Leaf petioles were carefully detached from plants,fresh weights were recorded, and then kept in water holding plastic tube in a closed container in an atmosphere saturated by means ofwet tissue paper around the inner wall of the container.Turgid weight was measured after 24 h and dry weight was measured after oven-drying for 48 h at 65°C.Leaf relative water content was calculated by the following equation:
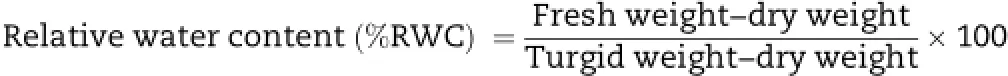
2.5.Measurement of leaf water potential
Leaf water potential(ψw)was measured at 0,1,3,and 7 days after soildrying and soildrying+exogenous ABA application using a pressure chamber(Soilmoisture Equipment Corp, Santa Barbara,California,USA).The leaf petiole was sealed into a pressure chamber and the chamber was gradually pressurized until the meniscus of the xylem sap becomevisible at the cut surface,at which time the pressure reading was recorded.
2.6.Determination of weight loss of excised leaves
Water loss from detached youngest mature leaves was determined following Okamoto etal.[15].One hour after application of exogenous ABA,leaves from well-watered and ABA-treated plants were excised with petiole and placed on a sheet of paper for air-drying in the greenhouse.Measurementwas performed on a clear,sunny day with 33±1°C temperature and 55%relative humidity.Weightloss(%)due to water loss from detached leaves was recorded at 10 min intervals with an analytical balance (Shimadzu AUW220D,Shimadzu Corporation,Tokyo,Japan).
2.7.Estimation of stomata on the leaf surface
Stomata on the youngest mature leaves of well-watered plants were counted under a light microscope at 40×(Nikon Eclipse 80i,Diagnostic Instrument).
2.8.Measurement of shoot and root weight and length, root-to-shoot ratio,and soil water content
After 7 days of progressive soil drying as well as soil drying+application of exogenous ABA,shoot and root fresh weight and length in drought-tolerant and droughtsusceptible soybean genotypes were measured.Root-to-shoot ratio was determined after oven drying(65°C for 48 h)of roots and shoots.Soilwater contentwas determined atthe beginning of soil drying(day 0)and after 7 days of drying.Soil was collected at a depth of 10-15 cm in the soil profile and oven dried at 105°C for 72 h,and water content was determined and expressed as a percentage on an oven-dry weight basis.
2.9.Statistical analysis
Data were subjected to one-way analysis of variance (ANOVA)followed by post hoc multiple comparisons using Tukey test to identify significant differences between treatments.The results presented are means with standard deviations of three to six replicates.The minimum accepted P-value for significance was 0.05.
3.Results
3.1.Soilwater status
Soil water content was monitored during the imposition of treatments as well as after the ending of soil drying for the drought-tolerant and-susceptible soybean genotypes.Soilwater content declined after seven days of the experiment(Fig.1).Soil water contents of C12 and C08 were 36.41%and 37.63%on day 0, falling to 23.48%and 23.57%,respectively at the end of the experiment(Fig.1).

Fig.1–Soil water content(%)at the day of imposition of soil drying and soil drying+ABA application(day 0)and after 7 days of soil drying for drought-tolerant(C12)and -susceptible(C08)soybean genotypes.Values of points are means±standard deviation(SD)(n=3).
3.2.Influence ofexogenous ABAon leafexpansion rate ofsoybean

Fig.2–Leaf expansion rate of drought-tolerant(C12)and-susceptible(C08)soybean genotypes after imposition of treatments. Empty black circles and black triangle represent soil drying(sd),soil drying+exogenous ABA application(sd+ABA),and well-watered(ww)plants,respectively.Each data point represents mean±SD(n=5).
Leaf expansion rates of soybean genotypes were measured daily after imposition of soildrying with or without exogenousABA application.Under soildrying with exogenous ABA application,leaf expansion rates declined in all genotypes(Fig.2). Compared with the drought-susceptible genotype(C08),the drought-tolerant genotype(C12)showed a more rapid reduction in leaf expansion rate after exogenous ABA application(Fig.2).
3.3.Influence of exogenous ABA on leaf stomatal conductance of soybean
Stomatal conductance(gs)of soybean genotypes was measured 3 h after imposition of soildrying with or without application of exogenous ABA,followed by daily measurement.Stomatal conductance of leaves was markedly affected by application of exogenous ABA(Fig.3).A rapid reduction of gswas observed after 3 h in the drought-tolerant genotype(C12)compared to the drought-susceptible genotype(C08)(Fig.3).
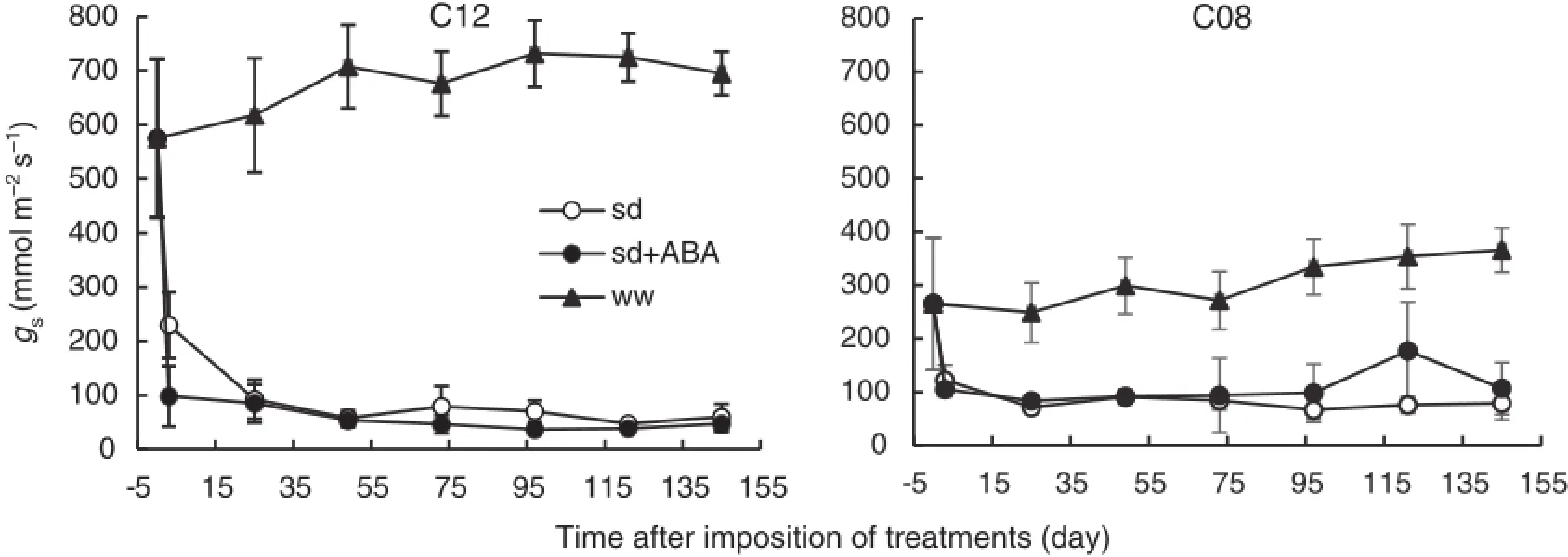
Fig.3–Stomatal conductance of drought-tolerant(C12)and susceptible(C08)soybean genotypes after imposition of treatments.Empty,black circles and black triangle represents soil drying(sd),soil drying+exogenous ABA application (sd+ABA)and well-watered(ww)plants,respectively.Each data point represents mean±SD(n=5).
3.4.Influence of exogenous ABA on%RWC andψwof soybean leaves
Percent relative water content(%RWC))and water potential(ψw) of soybean leaves were determined before and after imposition of treatments.As soil drying progressed,%RWC of leaves declined,but soil drying with application of ABA led to much slower reduction of%RWC in both soybean genotypes(Fig.4). The reduction tendency of%RWC of the drought-susceptible genotype(C08)was significantly faster from the initiation of soil drying and reached 65.7%after seven days,whereas that of the drought-tolerant genotype(C12)reached 78.5%.Exogenous ABA application may have induced stomatal closure in plants subjected to soil drying,reducing water loss through stomata and thereby preserving higher%RWC in leaves.
Exogenous application of ABA not only induced stomatal closure in leaves but also influenced leaf water potential(ψw) of soybean genotypes.Theψwof the drought-susceptible genotype fell sharply immediately after water was withheld and reached-2.3 MPa after seven days,while that of the drought-tolerant genotype reached-1.86 MPa(Fig.5).The influence of exogenous ABA on leafψwwas pronounced in the tolerant genotype C12.

Fig.4–Relative water content(%RWC)of leaves of drought-tolerant(C12)and susceptible(C08)soybean genotypes after imposition of treatments.Empty,black circles and black triangle represents soil drying(sd),soil drying+exogenous ABA application(sd+ABA)and well-watered(ww)plants,respectively.Each data point represents mean±SD(n=4).
3.5.Influence of exogenous ABA on water loss from excised soybean leaves

Fig.5–Leaf water potential(ψw)of drought-tolerant(C12)and susceptible(C08)soybean genotypes after imposition of treatments.Empty,black circles and black triangle represents soil drying(sd),soil drying+exogenous ABA application (sd+ABA)and well-watered(ww)plants,respectively.Each data point represents mean±SD(n=4).
Water losses(in%)from detached leaves of well-watered and exogenous ABA applied plants were determined.Fully expanded leaves were excised 1 h after ABA application along with well-watered leaves and left in the greenhouse for sun drying. Percentweightloss due to water loss fromleaves was recorded by weighing at 10 min intervals.Application of exogenous ABA inhibited water loss principally through leaf stomata,in comparison with well-watered leaves(Fig.6).Leaf water retention was pronounced 10 min after excision in the drought-tolerant genotype(C12)whereas the drought-susceptible genotype(C08) showed low water retention ability,such that%water loss was always higher in C08 even after application of ABA(Fig.6).Given that leaf stomata may have influenced%water loss from leaves of soybean genotypes,the stomata per square cm of leaf surface area were counted for both soybean genotypes under a confocalmicroscope.The presence of stomata on leaves was markedly different on abaxial and adaxial surfaces of both soybean genotypes but no significant difference was found between the genotypes(Fig.7).

Fig.6–Water loss(%)from well-watered(WW)and exogenous ABA applied leaves of drought-tolerant(C12)and susceptible (C08)soybean genotypes.Leaves were excised 1 h after application of ABA.Value of each data point represents a single leaf measurement.

Fig.7–Number of stomata on adaxial and abaxial surface of leaves of well-watered drought-tolerant(C12)and susceptible (C08)soybean genotypes.Value of each point is mean±SD(n=5).
3.6.Influence of exogenous ABA on root and shoot growth of soybean
To determine whether exogenous ABA affects root or shootgrowth ofsoybeangenotypes under water restriction during growth,root and shoot fresh weight and length and root-to-shoot ratio were measured at the end of the experiment.As a growth-retardantchemical,ABAmay affectshootand rootweight and length and root-to-shoot ratio in drought-tolerant(C12)and drought-susceptible(C08)soybean genotypes.Shoot and root lengths and fresh weight of the drought-tolerant genotype were reduced by soil drying in comparison with the drought-susceptible genotype,but not significantly(Fig.8A,B,C). Root fresh weight of both genotypes declined,but that of the drought-susceptible genotype was significantly reduced by water restriction for seven days(Fig.8D).The drought-tolerant soybean genotype(C12)showed a higher root-to-shoot ratio than the susceptible genotype(C08)under restricted soil water with application of exogenous ABA(Fig.8E).
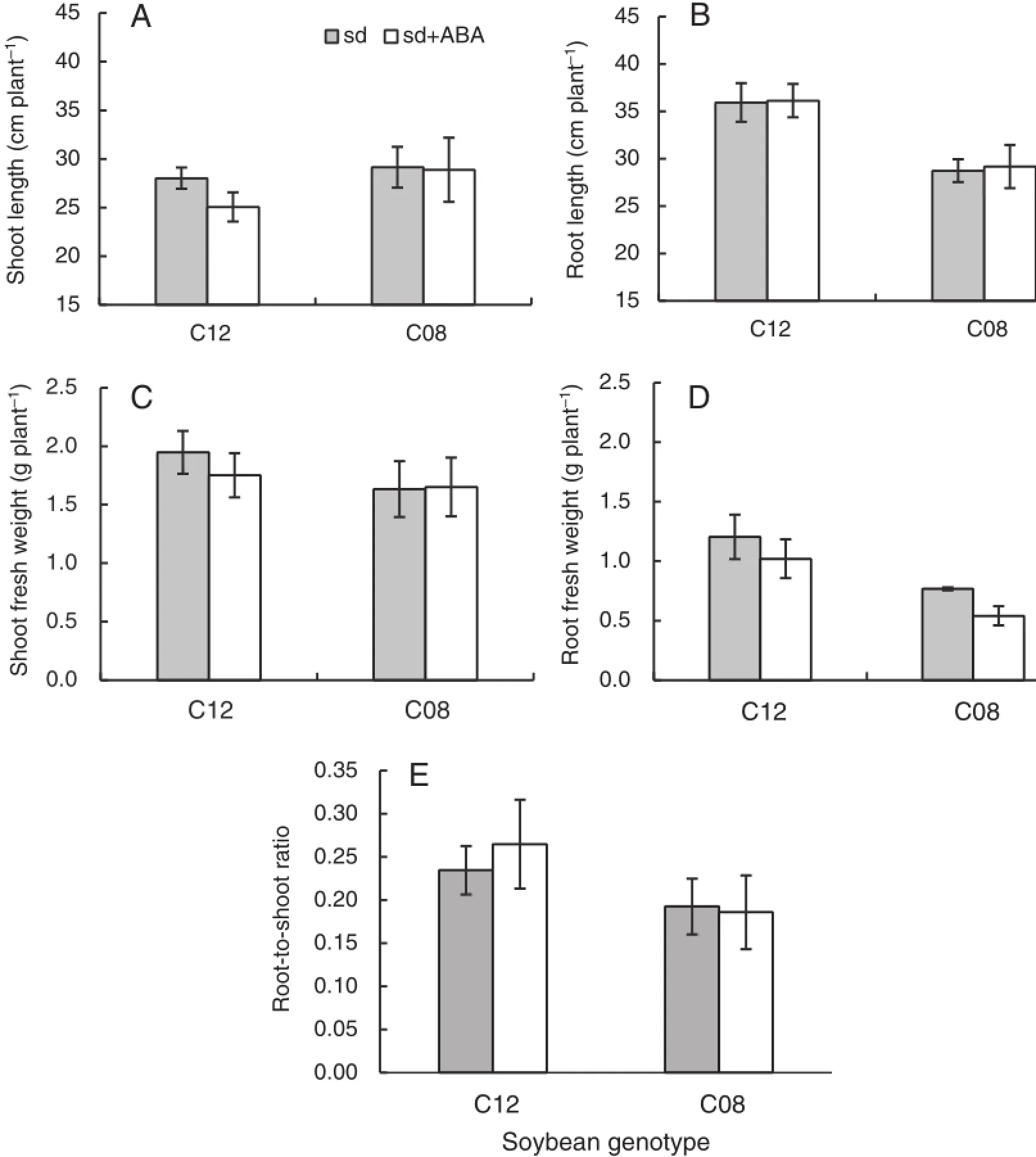
Fig.8–Shoot length(A),root length(B),shoot fresh weight(C),root fresh weight(D),and root-to-shoot ratio(E)of drought-tolerant(C12)and susceptible(C08)soybean genotypes after 7 days of soil drying with exogenous ABA application. Value of each point is mean±SD(n=7).
4.Discussion
The responses of tolerant and susceptible soybean genotypes to soildrying and to soildrying with ABAapplication were different. The drought-tolerant genotype responded more rapidly than the drought-susceptible one when water restriction along withexogenous ABA was imposed on the soybean seedlings. Water-restricted plants showed immediate low stomatal conductance while retaining higher leafwater potential.The possible reason behind these events is that leaf stomata could not open, because plants sense the availability of water in the soil and regulate stomatal behavior accordingly,whatever the water status of their leaves[2].ABA has been reported to reduce stomatal conductance,possibly reducing photosynthesis in plants butimproving water status in leaves[16,17].In the present experiment,leaf stomatal conductance(gs)of the droughttolerant soybean genotype declined within 3 h of exogenous ABA application followed by withholding ofwater to the soybean seedlings.Kimand Iersel[18]reported that ABAdrenches caused stomatal closure in salvia(Salvia splendens Sellow ex Schult.) within 3 h of application.In the present experiment,the tendency of water loss from the detached leaves of ABA-treated seedlings indicated that exogenous ABAcan act on stomata even 3 h after application,so that gsdeclined within this short period. Although progressive soil drying triggered the reduction of leaf water status and%RWC,these tendencies were different in the tolerant and susceptible genotypes.The reduction of gsin leaves was regulated by%relative water content ofleaves as wellas leaf water potential in the drought-tolerant genotype.Thus,the decline of%RWC and water potential were markedly slower in leaves of the drought-tolerant genotype than in those of the drought-susceptible genotype.Hossain et al.[19]proposed that drought-tolerant soybean genotypes maintained higher instantaneous water use efficiency(WUEi),%relative water content, water potential(ψw),and turgor potential(ψp)during progressive soil drying and subsequent rewetting than did susceptible genotypes.Application of exogenous ABA inhibited water loss from excised leaves by inducing stomatalclosure.In the present study,weight loss of excised leaves due to water loss was slower in the drought-tolerantgenotype.However,ABAaccumulation in plants is regulated by drought stress,which induces stomatal closure and restricts growth[20].The results ofour study suggest that exogenous ABA application induces stomatal closure, thereby helping to retain higher%RWC,leaf water potential, and restrict in weightloss fromexcised leaves throughwater loss, and that these responses differ between tolerant and susceptible soybean genotypes.
Acknowledgments
We thank Fuk-Ling Wong,Dr.Weifeng Xu and Dr.Ken Lau for technical support.This study was supported by the Hong Kong RGC Collaborative Research Fund(CUHK3/CRF/11G)to Prof.Hon-Ming Lam and Prof.Jianhua Zhang.
R E F E R E N C E S
[1]C.Pinheiro,M.M.Chaves,Photosynthesis and drought:can we make metabolic connections from available data?J.Exp. Bot.62(2011)869-882.
[2]W.J.Davies,J.Zhang,Root signals and the regulation of growth and development of plants in drying soil,Annu.Rev.Plant Physiol.Plant Mol.Biol.42(1991)55-76.
[3]P.Guo,M.Baum,S.Grando,S.Ceccarelli,G.Bai,R.Li,M.von Korff,R.K.Varshney,A.Graner,J.Valkoun,Differentially expressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage,J.Exp.Bot.60(2009) 3531-3544.
[4]B.Xoconostle-Cazares,F.A.Ramirez-Ortega,L.Flores-Elenes, R.Ruiz-Medrano,Drought-tolerance in crop plants,Am.J. Plant Physiol.5(2010)241-256.
[5]H.H.Chen,P.H.Li,M.L.Brenner,Involvement of abscisic acid in potato cold acclimation,Plant Physiol.71(1983)362-365.
[6]K.Cornish,J.A.D.Zeevaart,Abscisic acid metabolism in relation to water stress and leaf age in Xanthium strumarium, Plant Physiol.76(1984)1029-1035.
[7]J.Giraudat,F.Parcy,N.Bertauche,F.Gosti,J.Leung,P.C.Morris, M.Bouvier-Durand,N.Vartanian,Current advances in abscisic acid action and signaling,Plant Mol.Biol.26(1994)557-577.
[8]R.W.Wilen,L.V.Gustabolei,S.R.Abrams,B.E.Ewan,Effects of abscisic acid(ABA)and ABA analogs on freezing tolerance, low-temperature growth,and flowering in rapeseed,J.Plant Growth Regul.13(1994)235-241.
[9]F.Jiang,W.Hartung,Long-distance signaling of abscisic acid (ABA):the factors regulating the intensity of the ABA signal, J.Exp.Bot.59(2008)37-43.
[10]F.Liu,C.R.Jensen,M.N.Andersen,A review of drought adaptation in crop plants:changes in vegetative and reproductive physiology induced by ABA-based chemical signals,Aust.J.Agric.Res.56(2005)1245-1252.
[11]L.Zhang,M.Gao,J.Hu,X.Zhang,K.Wang,M.Ashraf, Modulation role of abscisic acid(ABA)on growth,water relations and glycinebetaine metabolism in two maize(Zea mays L.)cultivars under drought stress,Int.J.Mol.Sci.13 (2012)3189-3202.
[12]D.M.Pandey,C.L.Goswami,B.Kumar,Physiologicaleffects of plant hormones in cotton under drought,Biol.Plant.47(2003) 535-540.
[13]F.Liu,C.R.Jensen,M.N.Andersen,Pot set related to photosynthetic rate and endogenous ABA in soybean subjected to different water regimes and exogenous ABA and BA at early reproductive stages,Ann.Bot.94(2004)405-411.
[14]H.M.Lam,X.Xu,X.Liu,W.Chen,G.Yang,F.L.Wong,M.W.Li, W.He,N.Qin,B.Wang,J.Li,M.Jian,J.Wang,G.Shao,J.Wang, S.S.Sun,G.Zhang,Resequencing of 31 wild and cultivated soybean genomes identifies patterns of genetic diversity and selection,Nat.Genet.42(2010)1053-1059.
[15]M.Okamoto,F.C.Peterson,A.Defries,S.Y.Park,E.Nambara, B.F.Volkman,S.R.Cutler,Activation of dimeric ABA receptors elicits guard cell closure,ABA-regulated gene expression,and drought-tolerance,Proc.Acad.Natl.Sci.U.S.A.110(2013) 12132-12137.
[16]G.Egea,A.Verhoef,P.L.Vidale,Towards an improved and more flexible representation of water stress in coupled photosynthesis-stomatal conductance models,Agric.For. Meteorol.151(2011)1370-1384.
[17]S.Tonetto de Freitas,A.J.McElrone,K.A.Shackel,E.J. Mitcham,Calcium partitioning and allocation and blossom-end rot development in tomato plants in response to whole-plant and fruit-specific abscisic acid treatments, J.Exp.Bot.65(2014)235-247.
[18]J.Kim,M.W.van Iersel,ABA drenches induce stomatal closure and prolong shelf life of Salvia splendens,SNA Res. Conf.53(2008)107-111.
[19]M.M.Hossain,X.Liu,X.Qi,H.M.Lam,J.Zhang,Differences between soybean genotypes in physiologicalresponse to sequential soil drying and rewetting,Crop J.2(2014)1-15.
[20]S.R.Cutler,P.L.Rodriguez,R.R.Finkelstein,S.R.Abrams, Abscisic acid:emergence of a core signaling network,Annu. Rev.Plant Biol.61(2010)651-679.
5 June 2015
in revised form
Abbreviations:ABA,abscisic acid;gs,stomatalconductance;LA,leaf area;LL,leaf length;LW,leaf width;RWC,relative water content; ψw,leaf water potential.
.Tel.:+852 3943 6288;fax:+852 2603 6382.
E-mailaddress:jhzhang@cuhk.edu.hk(J.Zhang).
Peer review under responsibility of Crop Science Society of China and Institute of Crop Science,CAAS.
杂志排行

The Crop Journal的其它文章
- Interpreting genotype×environment interactions for grain yield of rainfed durum wheat in Iran
- Barnyard millet globalcore collection evaluation in the submontane Himalayan region of India using multivariate analysis
- Genetic gains in wheat in Turkey:Winter wheat for irrigated conditions
- Resistance to powdery mildew in the pea cultivar Xucai 1 is conferred by the gene er1
- A nucleotide substitution at the 5′splice site of intron 1 of rice HEADING DATE 1(HD1)gene homolog in foxtail millet,broadly found in landraces from Europe and Asia
- Effectofelevated[CO2]and nutrientmanagement on wetand dry season rice production in subtropicalIndia