杉木器管组织碳素含量变化特征
2015-12-21项文化
余 蓉 ,项文化 ,2
(1.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;2.湖南会同国家野外科学观测研究站, 湖南会同 418307)
杉木器管组织碳素含量变化特征
余 蓉1,项文化1,2
(1.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;2.湖南会同国家野外科学观测研究站, 湖南会同 418307)
在连续定位测定不同生长发育阶段杉木器管组织碳素含量的基础上,研究了不同生长发育阶段杉木器管碳素含量动态变化特征。结果表明:同一生长发育阶段,树叶的碳素含量(493.28~529.34 g·kg-1)>树皮(481.24 ~ 501.63 g·kg-1)>树根(473.25 ~ 483.61 g·kg-1)>树干(472.90 ~ 478.02 g·kg-1)>树枝(468.37 ~477.14 g·kg-1),杉木根头的碳素含量>大根>粗根>细根;不同器官之间碳素含量差异与器官的功能密切相关;不同生长发育阶段, 20年生杉木器官的碳素含量>18年生>14年生>11年生>7年生,多年生叶或枝的碳素含量>2 年生>1 年生>当年生;杉木器管组织碳素含量随林龄增大而增加,这与器管组织木质化程度提高有关。
杉木;器官组织;发育阶段;碳素含量;湖南会同
森林植物以光合作用形式同化大气中的CO2,将大气中CO2固定在植物组织內,对于减少大气中温室气体的浓度、降低温室效应以及应对全球气候变化,无疑具有十分重要的作用和地位。某种森林吸收多少大气中CO2,一般用森林生物现存量或生物产量(直接或间接测定的)乘以生物量中碳素含量(室内实验分析待岀)推算得岀。由此可见,各种森林植物生物量及其碳素含量是研究森林在降低温室效应、缓解全球气候变化作用大小的两个关键因子。因此,准确测定不同区域、不同森林植物的碳素含量是估算全国或区域森林生态系统降低温室效应能力的基础。目前,许多学者对我国不同森林植物群落的生物量(或生产力)进行了大量研究,并取得不少成果,但在森林群落组成树种的碳素含量的研究上却落后许多。
以往我国有关植物组织碳素含量的研究大都结合在研究某森林生态系统的碳素储量、分布和碳汇功能的研究中[1-12]。实际上这些研究大都只关注某种林分的碳素分配及其储量特征,对植物组织碳素含量只作为某一种计算参数,并未进行详细分析。虽然马钦彦等[13]专门研究了华北主要森林类型建群种的植物组织碳素含量,但也是研究某种植物某一生育阶段的组织碳素含量的静态特征。也有学者分析过某种植物不同生育阶段的组织碳素含量动态变化,如米老排Mytilaria laosensis[2]、杉木Cunninghamia lanceolata[14]、马尾松Pinus massoniana[15-16]、 楠 木Phoebe bourmei[17]、 落叶 松Larixganelini[10,18]等植物的不同生育阶段的器管组织碳素含量等,这些研究大都建立在“空间换时间”的基础上,由于不同空间的立地条件存在差异,研究结果可能与实际情况不一定相符。本研究利用国家野外科学观测研究站——中南林业科技大学湖南会同杉木林生态系统研究站的长期定位连续观测基础数据,探讨杉木不同生育阶段的其组织碳素含量动态特征,其目的是为杉木人工林碳汇功能的估算提供合理的参数。
1 研究地区概况
研究地设在我国著名“广木之乡”湖南会同县广坪深冲林场,由中南林业科技大学建立的会同杉木林生态系统国家野外科学观测研究站内进行。研究区地理位罝:109°45′E,26°50′N。研究地面向西北,坡地坡度平均约25°,海拔高度280~355 m。
湖南会同处于亚热带季风湿润气候区。年降水量1 100~1 400 mm,年均相对湿度80%以上,年均气温16.8 ℃。气候特点是:冬无严寒,夏无酷热,降水分布均匀,气候温和,这些条件均有利于杉木生长。林地土壤为中有机质厚层黄壤,土壤母质为震旦纪板溪系变质的板(页)岩。
该区域植被为常绿阔叶林,以壳斗科Fagaceae中常绿种类如青冈Cyclobalanopsis、石栎属Lithocarpus、栲属Castanopsis为建群种,然后是樟科Lauraceae楠属Phoebe、樟属Cinnamomum,山茶科Theaceae的木荷属Schoma、山茶属Camellia的一些树种组成。灌木主要有山苍子Litsea cuheha、杜茎山Maesa japonica、白桤木Pinus albicaulis、 白 背 叶Mallotusapelta、 冬青Ilex chinensis等,草本主要有梵天花Urenaprocumbens、鱼腥草Houttuynia cordata、仙鹤草Herba Agrimoniae、狗脊蕨Woodwardia japornica等。
研究区于1966年营造第1代杉木人工林(造林前原为阔叶次生残林),1987年冬季皆伐第1代杉木人工林。1988 年春,在采伐迹地全垦、挖大穴营造第2代杉木人工林(造林密度2 400株·hm-2)。第2代杉木人工林到2007 年时林龄为20 a。此时林分密度为2 310 株·hm-2,树木平均高度约13.8 m,树木平均胸径在14.1 cm左右,林分郁闭度0.9 左右,叶面积指数7.63[19]。
2 研究方法
2.1 样品取样
中南林业科技大学湖南会同杉木林生态系统研究站对2代杉木人工林进行多次生物量测定(分别在2代杉木人工林7.11、14、18 和20 年生时),测样样品的取样是在每次林分生物量测定时采集的。取样品的方法是在伐倒标准木(林木的平均测树因子分析得出)上采取树干、树皮检测样品。杉木根把它分成根径大于1 cm的大根、根径大于0.2 cm且少于1 cm的粗根、根径少于0.2 cm的细根和根头分别取样。杉木叶和枝则按多年生叶、枝,2 年生叶、枝,1 年生叶、枝和当年生叶、枝分别取样。
2.2 碳素含量的测定
将采集的杉木各器管样品在实验室烘干、粉碎、过筛后,植物组织碳素含量采用重铬酸钾氧化-外加热法测定。
3 结果与分析
3.1 同龄杉木各组织的碳素含量
从表1看出, 同一林龄级杉木的不同生长发育的叶片(多年生、2 年生、1 年生和当年生)碳素含量存在差异,其碳素含量大小顺序为多年生>2年生>1 年生>当年生。但是,多年生、2 年生、1 年生和当年生叶相互之间的碳素含量差异不显著(P≥0.05)。从表2可知,同一林龄级杉木的不同生长发育多年生、2 年生、1 年生和当年生枝的碳素含量的变化特征与同一林龄级杉木不同生长发育的叶片相同,也为多年生>2 年生>1 年生>当年生,且它们之间的碳素含量不存在显著差异(P≥0.05)。

表1 不同年龄阶段杉木叶组织的碳含量†Table 1 Carbon contents in needle organs of Chinese fir plantation in different age stages (g·kg-1)

表2 不同年龄阶段杉木枝组织的碳含量Table 2 Carbon contents in twig organs of Chinese fir plantation in different age stages (g·kg-1)
从表3可知,同一林龄级根随根径的减少其碳素含量也减少,碳素含量表现为根头>大根>粗根>细根。差异分析(见表4)表明,同一林龄杉木的大根与根头之间,7 年生和14 年生杉木粗根分别与7 年年和14 年生细根之间碳素含量差异并不显著(P≥0.05);20、18、11 年生杉木粗根分别与20、18、11 年生杉木的细根之间碳素含量存在显著差异(P<0.05);其余任一龄级内,不同根径的根之间碳素含量差异性见表4。
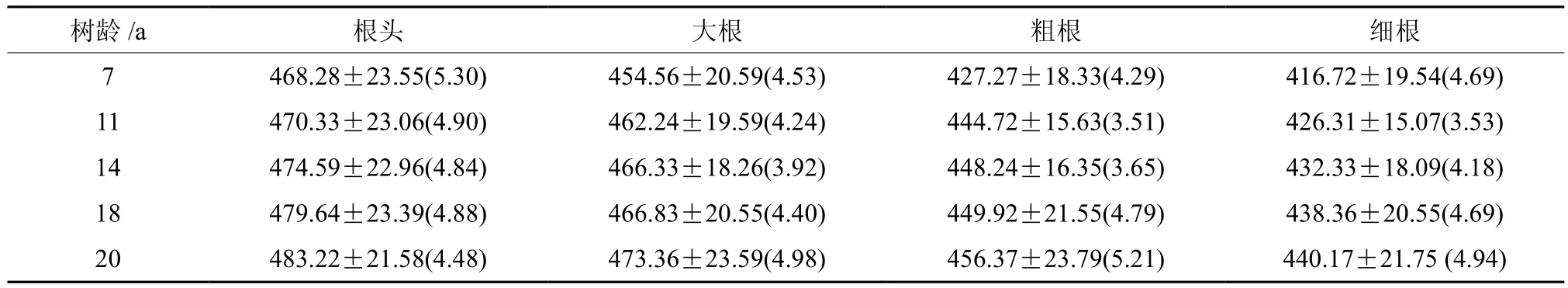
表3 不同年龄阶段杉木根的碳含量Table 3 Carbon contents in root organsof Chinese fir plantation in different age stages (g·kg-1)
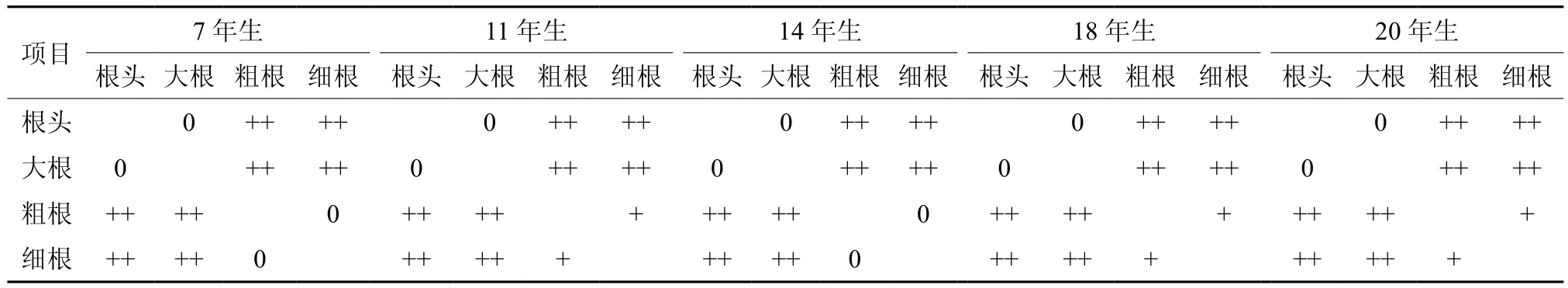
表4 同一林龄不同根径根的碳含量差异分析†Table 4 Difference analysis on roots’ carbon contents with different root diameter at same forest age
表5是将某一生长阶段(某一林龄如林分7年生)测定的多年生、2 年生、1 年生和当年生叶和枝,细根、粗根、大根和根头的碳素含量加权平均后,分别计算某一生长阶段的杉木叶、枝、根的平均碳素含量。从 表5中不难看出,同一林龄杉木不同器官组织彼此之间碳素含量存在差异,杉木各器官组织碳素含量大小顺序是:树叶(493.28~529.34 g·kg-1)>树皮(481.24~501.63 g·kg-1)>树根(473.25~483.61 g·kg-1) > 树 干(472.90 ~ 478.02 g·kg-1)>树枝(468.37 ~ 477.14 g·kg-1)。

表5 不同生长阶段杉木各组分碳含量†Table 5 Components' Carbon contents of Chinese fir plantation in different growth stages (g·kg-1)
同一生长阶段杉木各器管之间碳素含量差异分析(见表6) 表明,无论那一生长阶段的杉木叶与杉木树根、树枝、树皮及树干的碳素含量差异是很大的,有的达到极显著水平(P<0.01);同样,无论那一生长阶段内,杉木树皮与杉木树枝的碳素含量差异也达到显著水平(P<0.05)或极显著水平(P<0.01;20、18和14年生的树皮分别与20、18和14年生的树干以及与同龄生长的树根之间碳素含量差异极显著(P<0.01)或差异显著(P<0.05)。

表6 同一生长阶段不同杉木器官的碳含量差异分析†Table 6 Difference analysis on carbon contents in different components of Chinese fir plantation in same growth stage
3.2 不同林龄杉木器官的碳素含量
表5表明,随着林龄的增加,杉木各器官组织碳素含量(树叶、树枝、树根均为加权平均值)逐渐增大。不同生长阶段的同一器管组织碳素含量存在差异,但差异显著性有所不同。从表7可知,无论在那个生长阶段的杉木树根之间、树干之间碳素含量差异,但并不显著(P≥0.05);然而,树叶之间碳素含量,20 年生分别与7、11、14 年生差异极显著(P<0.01),18年生与20年生差异显著(P<0.05)。树皮之间碳素含量,20 年生分别与7 和11 年生,18 年与生11 年生差异显著(P<0.05)。树枝之间碳素含量,20 年生杉木树枝与7年生杉木树枝差异显著(P<0.05)。
前己分析,同一林龄的多年生、2 年生、1 年生和当年生叶之间,多年生、2 年生、1 年生和当年生枝之间的碳素含量差异不显著。但是,从表8可看岀,20 年生杉木当年生叶分别与7、11、14年生杉木当年生叶, 1年生叶和2年生叶分别与18、14、11和7年生杉木1年生叶、2年生叶,多年生叶分别与11和7年生杉木多年生叶;18年生杉木当年生叶与7年生杉木当年生叶, 1年生叶分别与14和7年生杉木1年生叶, 2年生叶分别与11和7年生杉木2年生叶,多年生叶与7年生杉木多年生叶之间的碳素含量差异极显著(P<0.01)。此外,叶之间的碳素含量差异显著(P<0.05):有18年生分别与20、14、11年生的杉木当年生叶;18年生与11年生的杉木1年生叶,14年生与7年生的杉木1年生叶;14年生分别与7、18年生的杉木2年生叶;18年生与11年生的杉木多年生叶,14年生与7年生的杉木多年生叶,20年生分别与14、18年生时的多年生叶。
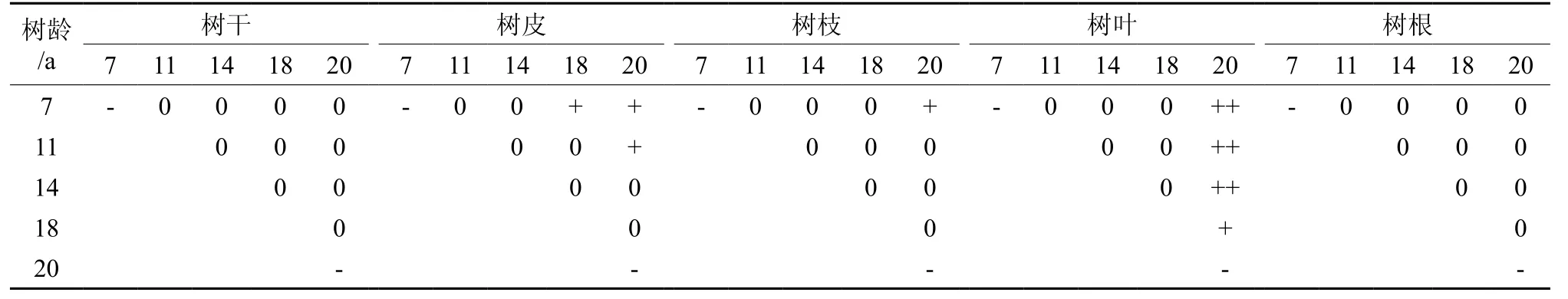
表7 不同生长阶段同一杉木器官的碳含量差异分析Table 7 Difference analysis on carbon contents in same component of Chinese fir plantation in different growth stages

表8 不同生长发育的杉木叶碳含量差异分析Table 8 Difference analysis on carbon contents of Chinese fir leaves in different growth and development stages
表9表明,不同生长阶段的不同生长发育枝的碳素含量有差异。碳素含量差异极显著(P<0.01)的有7年生杉木的多年生、2年生、1年生和当年生枝分别与18和20年生杉木多年生枝、2年生、1年生、当年生枝;碳素含量差异显著(P<0.05)的有7年生杉木的多年生与14年的多年生,7年生杉木的2年生枝与14年生的2年生枝。
表10表明,无论是根头、大根、粗根和细根,7年生分别与20年生时的碳素含量;18年生杉木细根和7年生细根,18年生杉木粗根和7年生粗根之间碳素含量差异极显著(P<0.01)。11年生和20年生杉木根头与根头、大根与大根、粗根与粗根,细根与细根,7年生和14年生杉木粗根与粗根,细根与细根之间碳素含量差异显著(P<0.05)。其余不同林龄同一根径的根之间碳素含量差异不显著(P≥0.05)。

表9 不同生长发育的枝碳含量差异分析Table 9 Difference analysis on carbon contents of Chinese fir branches in different growth and development stages

表10 不同根径的杉木根碳含量差异分析Table 10 Difference analysis on carbon contents of Chinese fir roots with different root diameters
4 结论与讨论
在我国杉木中心产区会同的同一林龄杉木的不同器官组织碳素含量不一样,其大小排序为:树叶>树皮>树根>树干>树枝。杉木各器官的功能是不同的,构建器官组织的有机物质也不相同,这就决定了各器官组织的碳素含量不一样。树木能挺立,主要依靠树干支撑,组成树干的有机化合物主要是低碳的结构形拉力较强的粗纤维等[20]。树叶是吸收大气二氧化碳和大阳能制造有机物质的器官,植物所需的养分和水分主要靠树木根系吸收,树皮是输送水分和营养物质的器管。一般而言,非结构形的粗脂肪和蛋白质等高碳有机化合物是组成叶、皮、根组织主要的有机化合物[21-22]。可能由于这种原因,使得同一龄级的杉木叶、皮、根组织的碳素含量高于杉木的干与枝。不言而喻, 根头木质化程度要依次高于大根、粗根、细根,木质素也属高碳有机化合物[23],因此,同一林龄级中根的碳素含量隨根径减少而降低。
本研究结果与闽北林区同一林龄杉木皮>叶>干>根>枝[14],广西大青山13年生杉木皮>叶>干>根>枝[1],南京下蜀杉木叶>皮>干>技>根[23]稍有不同;也与福建顺昌马尾松干>根>皮>枝>叶[16],广西大青山13年生马尾松干>根>枝>皮>叶[1],楠朩皮>干>根>枝>叶[17],华北8个乔木树种均为皮>根>叶>干>枝[14],南京下蜀的栓皮栎和国外松叶、皮、干材的碳素含量较高,枝和根系中较低[23],浙江建德青冈器官碳含量叶或地上部分器官中相对较高而根中相对较低[5]等存在差别。不同的立地以及不同的气候条件下都可能引起植物器官碳素含量的变化[24]。由此可见,即使同一树种在不同的立地条件下其组织碳素含量也不相同。会同杉木与其它地方其它树种器官碳素含量的差别,这里不仅有空间异质性的原因,而且物种不同,可能是由于不同树种的遗传特性,因而其组织碳素含量也不一样。
会同杉木器官碳素含量随着树木年龄增大而增大。这与尉海东研究的杉木中龄林最大, 成熟林次之,幼龄林最小[14],华北落叶松树干、树枝、树皮、树叶、根含碳率并不一定随林龄增长而增大[25];而与马尾松成熟林最大,中龄林次之,幼龄林最小[16];楠木成熟林>中龄林>幼龄林[17];米老排各器官碳素含量27年生>20年生>10年生[2]的结果相一致。树木在生长过程中,构成各器官组织的有机物质及各机物质成构成比例会发生一定的变化。树木的林龄越大, 树木器官组织木质化程度也就越高。木质素属高碳有机化合物,意味着其组织内碳素含量也就增加了。另外, 树木在其生长过程中,并不是每年的气候条件都可能一样,在不同的太阳光能、不同的温度以及不同的降水量情况下,都可能引起植物器官碳素含量的变化[24]。但要做一个合理精确的解释必须建立在不同生长阶段其器官有机物质组成,以及有机物质的化学成分的生物化学定量分析上。
本研究的不同生长阶段杉木碳素含量动态特征是建立在定位连续测定的基础上。研究结果不仅具有时间尺度的连续性,而且也保障了空间尺度的一致性,因而结果更加逼近实际,这就是本研究在实际应用上的重要意义所在。因为,以往估算森林生态系统碳贮量,对森林植物含碳率的选择,一些研究者采用0.45[12,26-27],也有采用0.5的[28-35],很少用实测的含碳率估算。实际上不同种类的植物组织器官中碳素含量不同,同一种植物的不同组织器官其碳素含量也可能存在差别,即使同一立地条件的不同生长阶段某种树木的同一器官的碳素含量也不一样。这说明估算某个国家或某个区域的森林生态系统碳贮量时,应根据研究区域内不同生长阶段不同树种的森林群落植物组织碳素含量和生物量分别估算,若借用其它的资料或取某一平均数,其结果与实际可能会产生偏移。这也是以往不同学者的估算值间存在差异的主要原因之一。
由于在国家尺度甚至区域尺度还缺乏长期的积累资料和足够可信的观测数据,致使目前森林生态系统碳汇能力,还是其碳贮量的估算结果带有很大的不确定性。这种估算的不确定性影响了对全球变化预测的科学性和可靠性。全球碳平衡研究中所出现的“碳失汇”问题,很有可能是在对森林生态系统在许多方面还缺乏足够的了解所引起的[36-38]。因此,建立不同地区不同生长发育阶段的主要森林类型植物碳素含量和生物量数据库,是全球碳平衡研究中迫切需要解决的问题。
[1] 康 冰, 刘世荣, 张广军,等.广西大青山南亚热带马尾松、杉木混交林生态系统碳素积累和分配特征[J].生态学报,2006, 26(5): 1320-1329.
[2] 刘 恩,刘世荣.南亚热带米老排人工林碳贮量及其分配特征[J].生态学报,2012,32(16): 5103-5109.
[3] 方 晰,田大伦,项文化.速生阶段杉木人工林碳素密度、贮量和分布[J].林业科学,2002,38(3):14-19.
[4] 雷丕锋,项文化,田大伦,等.樟树人工林生态系统碳素贮量与分布研究[J].生态学杂志,2004,23(4):25-30.
[5] 李铭红, 于明坚, 陈启常, 等.青冈常绿阔叶林的碳素动态[J].生态学报, 1996,16(6): 645-651.
[6] 方运霆,莫江明.鼎湖山马尾松林生态系统碳素分配和贮量的研究[J].广西植物,2002,22(4):305-310.
[7] 涂 洁,刘琪璟.亚热带红壤丘陵区湿地松人工林生态系统碳素贮量与分布研究[J].江西农业大学学报, 2007, 29(1):48-54.
[8] 周国模,姜培坤.毛竹林的碳密度和碳贮量及其空间分布[J].林业科学,2004,40(4):20-24.
[9] 吴仲民, 李意德, 曾庆波, 等.尖峰岭热带山地雨林C 素库及皆伐影响的初步研究[J].应用生态学报, 1998,9(4) : 341- 344.
[10] 孙玉军,张 俊,韩爱惠,等.兴安落叶松幼中龄林的生物量与碳汇功能[J].生态学报2007,27(5): 1756-1762.
[11] 王金叶, 车克钧, 蒋志荣.祁连山青海云杉林碳平衡研究[J].西北林学院学报, 2000,15(1): 9-14.
[12] 王效科, 冯宗炜, 欧阳志云.中国森林生态系统植物碳储量和碳密度研究[J].应用生态学报, 2001,12(1): 13-16.
[13] 马钦彦,陈遐林,王 娟,等.华北主要森林类型建群种的含碳率分析[J].北京林业大学学报,2002, 24(6):96-100.
[14] 尉海东, 马祥庆.中亚热带不同发育阶段杉木人工林生态系统碳贮量研究[J].江西农业大学学报, 2006,28(2): 240-244.
[15 方 晰,田大伦,胥灿辉.马尾松人工林生产与碳素动态[J].中南林学院学报,2003,23(3):11-15.
[16] 尉海东, 马祥庆.不同发育阶段马尾松人工林生态系统碳贮量研究[J].西北农林科技大学学报:自然科学版,2007, 35(1):171-174.
[17] 尉海东, 马祥庆.不同发育阶段楠木人工林生态系统碳贮量研究[J].烟台师范学院学报:自然科学版,2006,22(3):15-19.
[18] 张田田,马履一,贾忠奎,等.华北落叶松幼中龄林的生物量与碳汇功能[J].北京林业大学学报, 2012,40(12):32-38.
[19] 杨 超,田大伦,胡曰利,等.连栽杉木林林下植被生物量动态格局[J].生态学报, 2011,31(10): 2737- 2747.
[20] Lieth H, Whittaker R.Primary productivity of the biosphere[M].New York: SpringerVerlag, 1975:119-129.
[21] Sinclair T R.Nitrogen in fluence on the physiology of crop yield [C]/ /Rab inge R, Goudrin J, Penningg deVries FW T,et al.Theoretical Producion Ecology: Reflect ion s and Prospects.Wageningen: Pudoc, 1990: 41-55.
[22] Qu G H, Wen M Z, Guo J X.Energy accumulation and allocation of main plant populations in Aneurole pidium chinenes grasslandin Songnen Plain[J].Chinese Journal of Applied Ecology, 2003 ,14(5): 685-689.
[23] 阮宏华,姜志林,高苏铭,等.苏南丘陵主要森林类型碳循环研究[J].生态学杂志,1997,16(6):17-21.
[24] Jordan C F.Productivity of a tropical forest and its relation to a world pattern of energy storage[J].J.Ecol., 1971, 59: 127-142.
[25] 张田田,马履一,贾忠奎,等.华北落叶松幼中龄林的生物量与碳汇功能[J].北京林业大学学报, 2012,40(12):32-38.
[26] 周玉荣, 于振良, 赵士洞.我国主要森林生态系统碳贮量和碳平衡[J].植物生态学报, 2000,24(5) 518-522.
[27] 赵士洞, 汪业勖, 于振良, 等.中国森林生态系统碳循环研究[C].见:中国生态学会编.中国生态学会通讯 (特刊), 2000:50-52.
[28] 方精云.中国森林生产力及其对全球气候变化的响应[J].植物生态学报, 2000,24(5): 513-517.
[29] 马钦彦, 谢征鸣.中国油松林储碳量基本估计[J].北京林业大学学报, 1996,18(3): 31-34.
[30] 贺庆棠.森林对地气系统碳素循环的影响[J].北京林业大学学报, 1993,15(3): 132-136.
[31] Houghton R A, Skole D L, Nobre C A,et al.Annual fluxes of carbon from deforestation and regrowth in the Brazilian Amazon[J].Nature , 2000,403: 301-304.
[32] Fang J Y, Chen A P, Peng C H,et al.Changes in forest biomass carbon storage in China between 1949 and 1998[J].Science,2001, 292: 2320-2322.
[33] Nabuurs C J.Significance of wood product s in forest sector carbon[C]//Apps M J, Price D T.Forest Ecosystems, Forest Management and the Gl obal Carbon Cycle.Berlin: Springer-Verlag, 1996:245-256.
[34] Sykes M,Prentice C.Carbon storage and climate change in Swedish forest:a comparison of static and dynamic modeling approaches[C]//AppsM J, Price D T.Forest Ecosystems, Forest Management and the Global Carbon Cycle.Berlin Heidelberg:Springer-verlag, 1996: 69-77.
[35] 刘国华, 傅伯杰, 方精云.中国森林碳动态及其对全球碳平衡的贡献[J].生态学报, 2000,20(5) : 733-740.
[36] Tans P P, Fung I Y, Takahashi T.Observational constraints on the globalatmospheric CO2budget[J].Science, 1990,247: 1431-1438.
[37] Keeling R F, Piper S C, Heimann M.Global and hemispheric CO2sinks deduced from changes in atmospheric CO2concentration[J].Nature , 1996,381: 218-221.
[38] 张胜利,项文化,雷丕锋,等.湖南省2009年杉木林碳贮存量及未来固碳潜力分析[J].中南林业科技大学学报, 2014,34(6): 53-59.
Changing characteristics of carbon content inCunninghamia lanceolataorgan tissues
YU Rong1, XIANG Wen-hua1,2
(1.School of Life Science, Central South University of Forestry and Technology, Changsha 410004, Hunan, China; 2.National Field Station for Scientific Observation & Experiment in HuitongCounty, Huitong 418307, Hunan, China)
Based on continuously measured organ carbon content of Chinese fir (Cunninghamia lanceolata) at different growing stages in National Field Station for Scientific Observation & Experiment in Huitong County of Hunan province, the changing characteristics of organ carbon contents in Chinese fir at different growing stages were studied.The results show that the organ carbon contents in Chinese fir were ranked as follows: needle (493.28 ~ 529.34 g·kg-1) > bark (481.24 ~ 501.63 g·kg-1) > root(473.25 ~ 483.61 g·kg-1) > trunk(472.90 ~ 478.02 g·kg-1) > twig (468.37 ~ 477.14 g·kg-1), and the carbon contents in roots decreased with decreasing roots’ diameter,they ranked from big to small as: root head > big roots> coarse roots > fine roots at the same growing stage; Carbon contents were closely related to the differences between different organs and organ functions; The carbon content in organs in Chinese fir at different growing stages ranked as: 20-year-old> 18-year-old> 14-year-old> 11-year-old> 7-year-old, and the carbon contents ranked as perennial leaves or branches > 2-year-old > 1-year-old > current-year-old; The findings showed that the organ tissues carbon contents in Chinese fir increased with increasing stand age, which is related to organs tissue lignification improvement.
Chinese fir (Cunninghamia lanceolata); organ tissues; developmental stage; carbon content; Huitong county of Hunan province
S718.55
A
1673-923X(2015)09-0121-07
10.14067/j.cnki.1673-923x.2015.09.021
2015-04-10
国家林业软科学研究项目(2013-R10)
余 蓉, 讲师,博士研究生 通讯作者:项文化,教授,博士;E-mail:Xiangwh2005@163.com
余 蓉,项文化.杉木器管组织碳素含量变化特征[J].中南林业科技大学学报,2015, 35(9): 121-127.
[本文编校:谢荣秀]
