林分生物量碳计量模型的比较研究
2015-12-21胡砚秋苏志尧李佩瑗李文斌
胡砚秋,苏志尧,李佩瑗,李文斌
(华南农业大学 林学院,广东 广州 510642)
林分生物量碳计量模型的比较研究
胡砚秋,苏志尧,李佩瑗,李文斌
(华南农业大学 林学院,广东 广州 510642)
选取3个天然林群落作为研究对象,利用3种包含不同计量参数的生物量碳计量模型,即生物量因子法、异速生长方程法及材积源生物量法,分别计算林分碳储量并比较分析各模型计量结果的差异。结果表明:生物量因子法与材积源生物量法计算所得林分平均碳密度相近,分别为155.56和152.82Mg·hm-2,异速生长方程法的结果偏低,为118.44Mg·hm-2,生物量因子法计算的不同群落的林分碳储量比异速生长方程法的高22.11% ~43.02%;各群落的立木结构及物种组成存在显著差异,均方根误差分析显示生物量因子法对群落碳密度的差异反应最为敏感,计量精度最高;各方法计量结果均显示中、大径级立木是林分碳储量的主要贡献者,中径级立木与大径级立木中计量精度较高的模型分别是异速生长方程法与生物量因子法。综合考虑计量精度及参数获取的便利性,3种计量模型各有优势,在实际应用中可以根据具体情况选择较为适合的模型,一般情况下可使用材积源生物量法,能便利地获得与采用包含木材密度参数的生物量因子法最接近的计量结果。
林分生物量;碳计量模型;木材密度;生物量因子法;异速生长方程;材积源生物量法
大气中以CO2为主的温室气体浓度不断升高,导致全球变暖问题日益严峻。森林是陆地生态系统的主体,也是陆地上最大的碳库[1],其CO2的源汇功能在全球碳循环中占有重要地位[2]。清洁发展机制(Clean Development Mechanism,CDM)中明确指出造林和森林管理是增加碳汇的可行方式。作为碳循环研究以及碳汇造林的基础环节,森林碳汇的精确计量是当前碳汇研究的重要议题之一[3]。
由生物量推算碳储量是目前森林碳汇计量最为常用的方法[4-5],生物量因子法、异速生长方程法以及材积源生物量法均是常见的生物量碳计量模型。生物量因子法是政府间气候变化专门委员会(IPCC)重点推荐的计量方法,基本木材密度(Basic wood density,WD)是该方法的重要参数之一。木材密度是木材的主要性质,也是树木重要的功能性状之一[6]。同时,木材密度也是森林地上生物量重要的决定因素[7]。研究表明木材密度与林木生长速率、耐阴性均有显著的相关关系[8-9],对于林木生长环境(温度、降水、土壤肥力)等均有良好的响应[10]。木材密度可以通过树木生长锥取样进行测定[11]。此外,生物量因子法计量参数还包括生物量转化与扩展因子(Biomass conversion and expansion factor,BCEF)、生物量扩展因子(Biomass expansion factor,BEF)、根茎比(Root:shoot ratio,R)等。异速生长方程法及材积源生物量法均通过实测数据建立立木胸径、树高或是蓄积量与生物量的拟合方程,再将方程推广以计算森林生物量。
随着碳汇造林的开展,林分尺度碳汇计量的需求进一步扩大。生物量因子法是IPCC重点推荐的计量方法,因计量模型中包含了木材密度等参数,计量精度高,但同时相关参数的数据采集耗费的人力物力大。如何兼顾碳汇计量的效率与精确性对碳汇计量提出了更高的要求。本文中在林分尺度上对3种常见的生物量碳计量模型进行对比研究,探索适用于小尺度碳汇计量的有效方法,以期为碳汇精确计量提供科学依据。
1 研究区域概况
研究样地位于广东东源康禾省级自然保护区内(23°44′~23°53′N,115°04′~115°09′E)。该保护区处于低山丘陵地带,山体呈东北-西南走向,最高峰白石岗海拔为839.7m。区内气候属亚热带季风气候,光热充足。据保护区资料记载,该区年平均气温20.3~21.1℃,无霜期达345~350d,年均降水量2 142.6mm,年平均相对湿度为77.0%。区内土壤土层深厚,主要类型为红壤。植物资源丰富,地带性典型植被为亚热带常绿阔叶林,一些林分内仍存在杉木 Cunninghamia lanceolata等演替早期的针叶类树种。
2 研究方法
2.1 样地调查
在保护区天然次生常绿林内中选取3个面积各为1hm2的样方,利用全站仪按水平距离将每一样方划分为25个20m ×20m小样方进行调查。采用相邻格子样方法对样方内胸径(Diameter at Breast Height,DBH)≥ 1cm以上的乔木立木进行每木检尺,记录种名、胸径和树高。其中,DBH ≤6cm的立木胸径用游标卡尺测量,DBH>6cm的立木则用胸径尺测量,树高以目测为主。
2.2 立木蓄积量的计算
将每木调查所得立木胸径及树高代入各类立木相应的二元立木材积方程,以计算单株立木的蓄积量。林分蓄积量为林分内所有单株立木蓄积量之和。不同种类立木二元立木材积方程①二元立木材积式引自:广东省林业局&广东省林业调查规划院.《广东省森林资源调查常用数表》.2009。如下。
针叶类:

软阔类:

硬阔类:
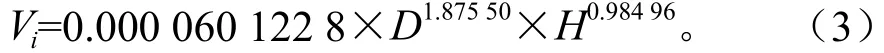
式中:Vi为单株立木的蓄积量(m3);D为胸径(cm);H为树高(m)。
2.3 生物量的计算
生物量计算通过3种方法计算,即生物量因子法、异速生长方程法、材积源生物量法。计算过程中所需的各类参数优先考虑来自当地的参数及最新的国家水平的参考值,如果没有对应的参数值,则选用IPCC提供的参考值。
2.3.1 生物量因子法
采用包含基本木材密度、生物量扩展因子及根茎比的生物量因子法。基本木材密度也称为树干材积密度,即每立方米木材所含干物质质量。林分生物量(B)计算公式[12-13]如下:

式中,V为林分蓄积量(m3);D为基本木材密度(Mg·m-3);F即林木地上生物量与树干生物量的比,无量纲;R为根茎比。群落主要优势种木材密度按树种区分,其余树种按立木类型取不同数值(见表1)。F按立木类型区分,硬阔类取1.79,软阔类取1.54[14]。R统一取0.24[13]。
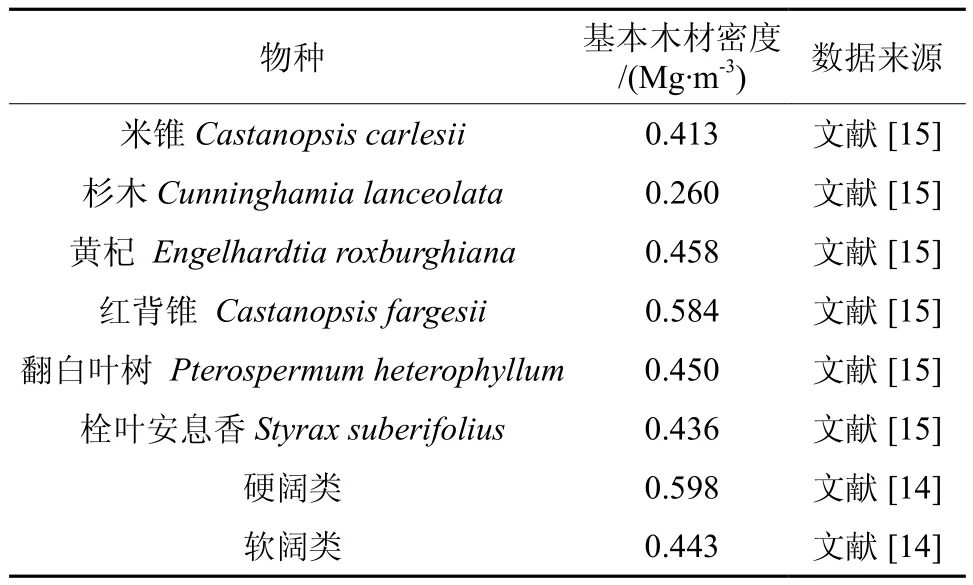
表1 群落优势种基本木材密度Table 1 Wood density of dominant species in community
2.3.2 异速生长方程法
异速生长方程是在实地测量标准木生物量的基础上,拟合生长曲线,建立胸径、树高与生物量的拟合方程。立木各部分异速生长方程见表2。其中,D为胸径(cm),H为树高(m)。全树总生物量W 由各部分生物量汇总求得。

表2 各类立木异速生长方程†[16]Table 2 Allometric equations of different wood types
2.3.3 材积源生物量法
林分材积综合反映了林龄、立地、个体密度和林分状况等因素的变化。材积源生物量法是建立蓄积量与生物量之间的函数关系,由蓄积量推算生物量。立木生物量在林分蓄积量的基础上以群落为单位,依照徐新良等[17]建立的常绿阔叶林生物量-蓄积量拟合方程进行计算:
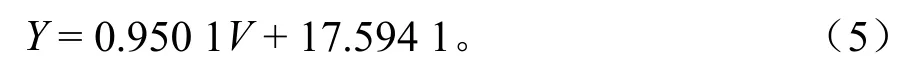
式中:Y为林分生物量(Mg);V为蓄积量(m3)。
2.4 碳储量的计算
不同植物的含碳率不同,但含碳率与生态特征(木材密度、树高)或统计特征(相对生长速率、死亡率)无显著相关性[18]。碳储量由生物量乘以含碳率计算求得,本文含碳率统一采用0.47计算[13]。
3 结果与分析
3.1 群落立木结构及物种组成
样地中共记录到立木14 136株,隶属于103种41科67属。3个群落同为常绿阔叶林,群落结构与起源基本一致,主要优势种相近。群落优势种多为硬阔类树种,仅黄杞Engelhardtia roxburghiana、翻白叶树 Pterospermum heterophyllum为软阔类树种。群落中亦混生有少量针叶类的杉木,但不占优势,不影响群落的性质。群落间胸径及树高差异极显著(p<0.001),群落2立木株数较少,但平均胸径及平均树高最大,其蓄积量也最大(见表3)。MRPP在检验多实体间差异性时,不需要考虑多元正态分布和方差齐性等分布假设,因此常用于群落生态学检验群落差异的研究中[19]。T是描述组间分离的检验统计量,T为负值时,其绝对值越大,组间分离越强。MRPP分析结果表明,各群落物种组成存在显著差异(见表4)。
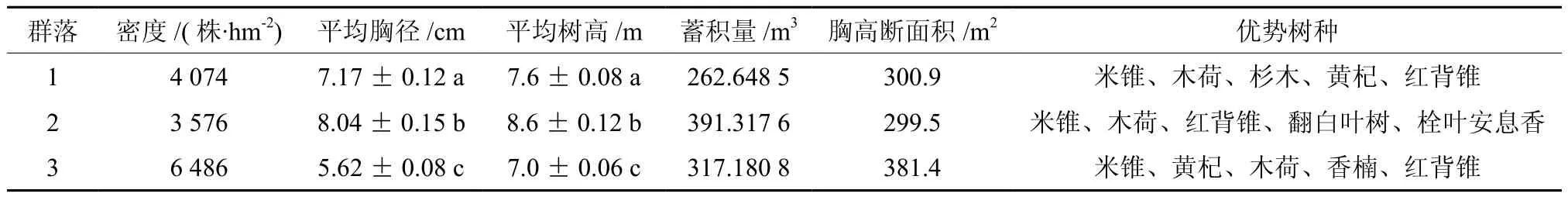
表3 各群落立木结构及物种组成概况†Table 3 Species composition and stand structure of different communities
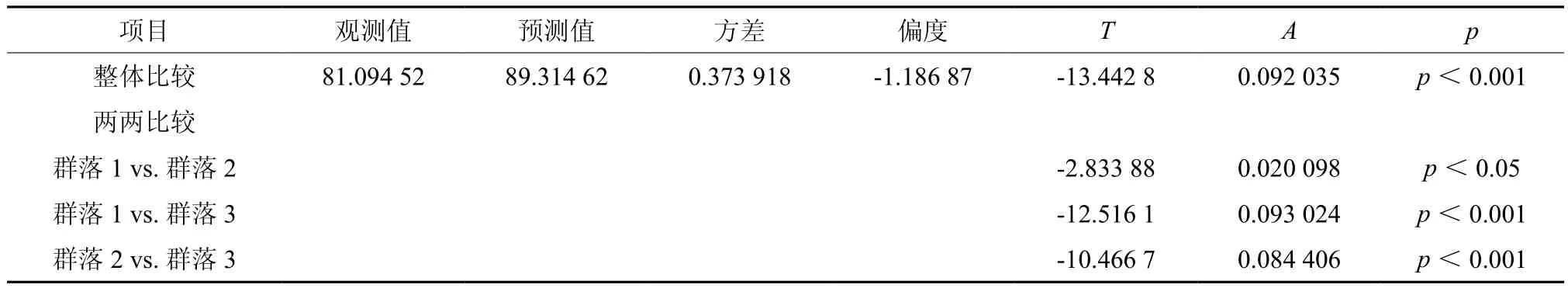
表4 群落物种组成MRPP分析Table 4 MRPP for species composition among different communities
3.2 群落林分碳储量计量
3种方法计算所得群落平均碳密度不同,从大到小依次为生物量因子法(155.56Mg·hm-2)、材积源生物量法(152.82Mg·hm-2)、异速生长方程法(118.44Mg·hm-2)。各群落中生物量因子法与材积源生物量法所得碳储量结果相近,但高于异速生长方程法的22.11%~43.02%(见表5)。采用均方根误差(Root Mean Square Error,RMSE)评价各计量方法精度,同一方法对不同群落计量结构的RMSE可以反映群落间碳储量的离散程度,RMSE越大,变异程度越大。由于所研究的群落其结构及物种组成均存在显著差异,RMSE较大说明该方法对群落间的差异敏感,3种计量方法中生物量因子法的RMSE最大(19.95),表明生物量因子法对群落间的差异敏感,准确性较高,其次是材积源生物量法,异速生长方程法精度最低。在蓄积量较高的群落中,所得计量结果离散程度高,蓄积量最大的群落2中,RMSE最大为17.91,变异系数为18.16%(见表5)。
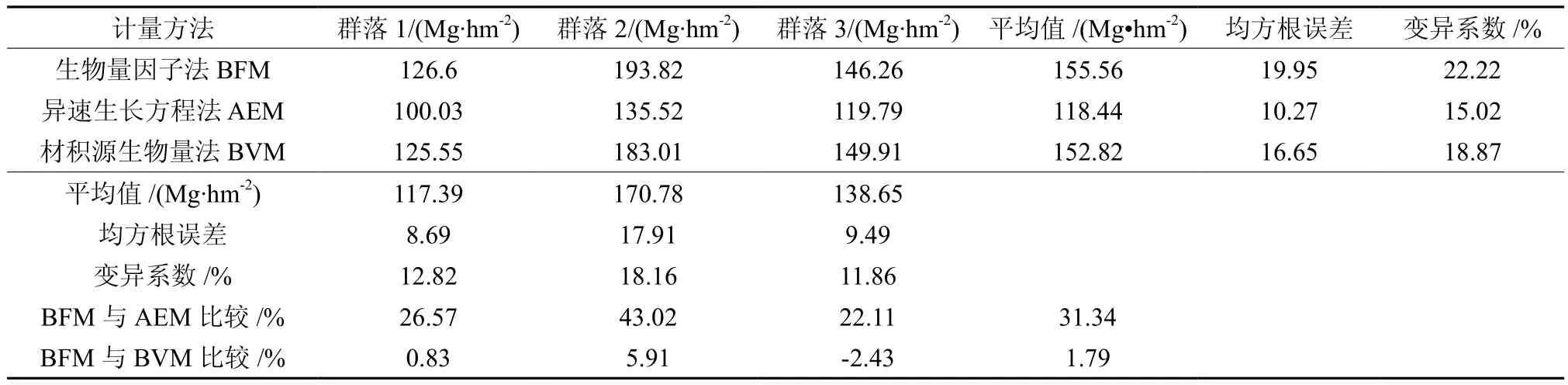
表5 不同碳计量方法群落林分碳储量计算结果比较Table 5 Comparison of community stand carbon storage calculated by different carbon accounting methods
3.3 各径级立木碳储量计量
3种方法计算所得各径级立木碳储量变化趋势一致,小径级立木碳储量最小,中径级立木(占总碳储量的35%~39%)及大径级立木(占总碳储量>57%)是林分碳储量的主要贡献者。总体来看,生物量因子法所得各径级立木碳储量计量结果大于另2种方法(见图1)。在小径级及大径级立木中生物量法的RMSE最大,精度最高,中径级立木精度最高的方法是异速生长方程法(见表6)。
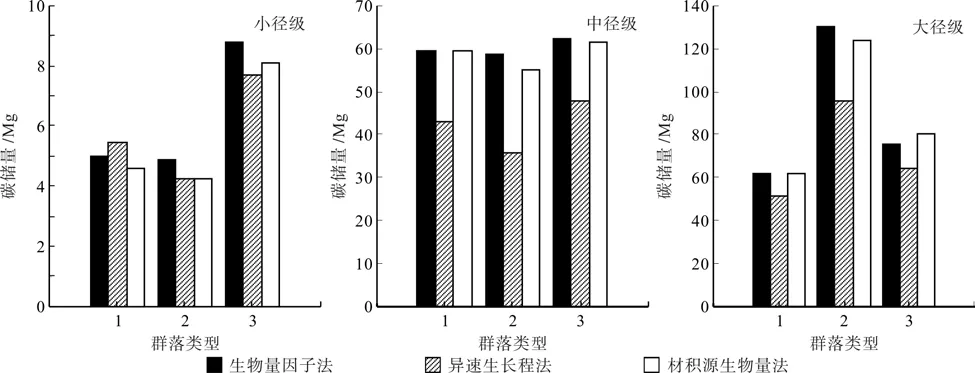
图1 不同方法计算所得各径级立木碳储量Fig.1 Stand carbon storage of individuals in different DBH classes by different methods
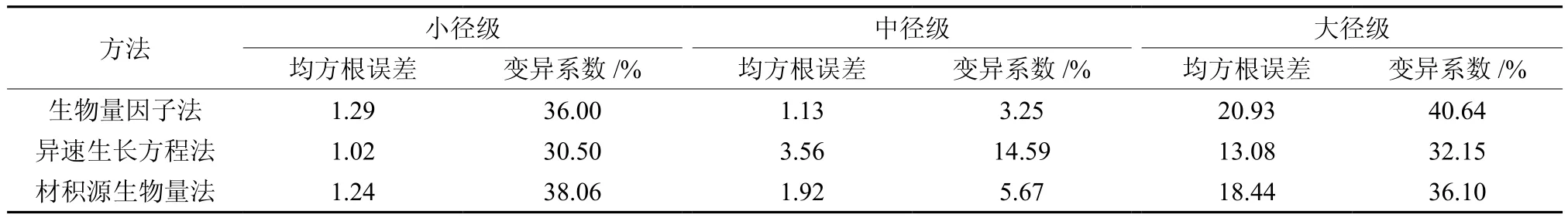
表6 不同方法所得各径级立木碳储量计量结果比较†Table 6 Comparison of stand carbon storage of different DBH classes by different methods
4 结论与讨论
研究中的3个群落均为地带性亚热带常绿林,优势种相近,多为米锥、木荷,但群落立木结构及物种组成间均存在显著差异。不同方法的计量结果显示,生物量因子法与材积源生物量法所得结果相近,异速生长方程法所得碳储量最小。其中,生物量因子法对群落间差异敏感,精度最高。分别计算不同径级立木的碳储量并对比,结果表明在中径级立木的碳储量计量中异速生长方程法精度最高,小径级及大径级立木均为生物量因子法中精度最高。
森林尤其是天然林中,物种组成及立木结构复杂,生物量分布空间变异较大,若按照单一的生物量换算因子必然造成计量误差,采用能够反映森林空间变异的碳计量参数是十分必要的。木材密度对于林分物种组成及生境条件均有良好的反映,同时,木材密度可通过生长锥取样测定,测定快速且对树木损伤小,实地测定简单易行。根据IPCC相关规定,生物量因子法可以用于区域的森林生物量计算,也可用于林分生物量计算,研究结果也表明生物量因子法计量精度最高。可以认为生物量因子法是较理想的碳计量方法,但是该方法包含参数多,若要获取全部参数资源耗费较大。材积源生物量法与生物量因子法计量结果相近,计量精度也较高,相对于生物量因子法参数获取较为便利。异速生长方程作为林业实践中常用参数,反映区域特征的地方性参考值虽较易获得,但从计量精度考虑则逊于其他两种方法。
随着碳汇研究的发展,林分尺度碳汇计量需求也越来越大,一些较为先进的方法如碳通量法、遥感法,限于技术要求较高,在小尺度碳汇计量中难以推广。生物量碳计量模型中的各计量参数若要获得基于样地实测的参数,需伐倒树木进行测量,破坏性强,人力时间耗费巨大。林分尺度的碳汇计量必须同时兼顾结果的精确性与资源的有限性,合理利用现有参数提供了寻求二者平衡的有效途径。本文对比研究的3种生物量碳计量模型在计量中各有优势,在开展小尺度碳汇计量时,应在前期工作中,总结区域范围内已开展的研究,考虑现有研究已获得的数据,选择适合的计量模型,再结合实地测量实现较为精确的碳计量。
[1] Pan Y,Birdsey R A,Fang J,et al.A large and persistent carbon sink in the world’s forests[J].Science,2011,333(6045):988-993.
[2] 方精云,陈安平.中国森林植被碳库的动态变化及其意义[J].植物学报,2001,43(9):967-973.
[3] Brown S.Measuring carbon in forests:Current status and future challenges[J].Environmental Pollution,2002,116(3):363-372.
[4] 孟晓清,刘琪璟,陶立超,等.大兴安岭地区南北部幼中龄林碳储量研究[J].中南林业科技大学学报,2014,34(06):37-43.
[5] 杨加猛,杜丽永,蔡志坚,等.江苏省森林碳储量的区域分布研究[J].中南林业科技大学学报,2014,34(07):84-89.
[6] Kraft N J,Valencia R,Ackerly D D.Functional traits and nichebased tree community assembly in an Amazonian forest[J].Science,2008,322(5901):580-582.
[7] Baker T R,Phillips O L,Malhi Y,et al.Variation in wood density determines spatial patterns in Amazonian forest biomass[J].Global Change Biology,2004,10(5):545-562.
[8] Muller-Landau H C.Interspeci fi c and inter-site variation in wood speci fi c gravity of tropical trees[J].Biotropica,2004,36(1):20-32.
[9] Thomas S C.Asymptotic height asa predictor of growth and allometric characteristics in Malaysian rain forest trees[J].American Journal of Botany,1996,83(5):556-566.
[10] Wiemann M C,Williamson G B.Geographic variation in wood specific gravity:Effects of latitude,temperature,and precipitation[J].Wood and Fiber Science,2002,34(1):96-107.
[11] 胡传双,苏志尧,云 虹,等.流体静力称衡法测定生长锥取样木材密度的改进方法[J].华南农业大学学报,2010,31(03):105-108.
[12] Ipcc.Good practice guidance for land use,land-use change and forestry[M].Hayama,Japan:IPCC National Greenhouse Gases Inventory Programme,Institute for Global Environmental Strategies(IGES),2003.
[13] Ipcc.IPCC guidelines for national greenhouse gas inventories[M].Japan:Institute for Global Environmental Strategies(IGES),2006.
[14] 中华人民共和国.中华人民共和国气候变化初始国家信息通报[M].北京:中国计划出版社,2004.
[15] 成俊卿,杨宗驹,刘 鹏.中国木材志[M].北京:中国林业出版社,1992.
[16] 国家林业局应对气候变化和节能减排工作领导小组办公室.造林项目碳汇计量与监测指南[M].北京:中国林业出版社,2008.
[17] 徐新良,曹明奎,李克让.中国森林生态系统植被碳储量时空动态变化研究[J].地理科学进展,2007,26(6):1-10.
[18] Martin A R,Thomas S C.A reassessment of carbon content in tropical trees[J].PLoS One,2011,6(8):23533.
[19] Mccune B,Mefford M J.PC-ORD.Multivariate Analysis of Ecological Data.Version 6.0 [M].MjM Software,Gleneden Beach,Oregon,USA:2011.
Comparative studies on biomass-carbon accounting models at forest stand scale
HU Yan-qiu,SU Zhi-yao,LI Pei-yuan,LI Wen-bin
(College of Forestry,South China Agricultural University,Guangzhou 510642,Guangdong,China)
To calculate the differences among carbon accounting models,the stand carbon storages in three natural evergreen forest communities were measured and compared by three biomass-carbon accounting models with different parameters,including biomass factor method(BFM)containing wood density,allometric equation method(AEM)and biomass-volume method(BVM).The results show that the average community carbon density calculated by BFM(155.56Mg·hm-2)and BVM(152.82Mg·hm-2)were close each other,while the value of AGM(118.44Mg·hm-2)was relatively lower,Carbon storage determined by BFM was 22.11%~43.02%higher than AGM in each community; Stand structure and species composition of the communities were signi fi cantly different,and the analysis of root mean square error(RMSE)showed that BFM was sensitive to the variability in carbon density,indicating higher measurement accuracy; Standing carbon storages by diameter class obtained by the three models changed consistently,ina way that individuals in middle and large diameter class were the major contributors to community carbon stocks,in the middle diameter class,AGM showed higher accuracy,while the accuracy of BFM was higher for large diameter class; Considering the convenience of access to the parameters and measurement accuracy,three models all had advantages and disadvantages,a reasonable model should be chosen according to the speci fi c condition in practice; In most cases,especially whena forest carbon accounting project requires high ef fi ciency and less effort,BVM is recommended because it isa convenient way to achieve the results that is most close to BFM,which requires the effort to collect wood density data and other local parameters.
stand biomass; carbon accounting model; wood density; biomass factor method; allometric equation; biomass-volume method
S718.5
A
1673-923X(2015)01-0084-05
10.14067/j.cnki.1673-923x.2015.01.016
2014-01-12
广东省林业科技创新专项(2010KJCX012-02,2011KJCX031-02)
胡砚秋,博士研究生;E-mail:xyzhyq@foxmail.com
苏志尧,教授,博导;E-mail:zysu@scau.edu.cn
胡砚秋,苏志尧,李佩瑗,等.林分生物量碳计量模型的比较研究[J].中南林业科技大学学报,2015,35(1):84-88.
[本文编校:谢荣秀]
