宽颌辽西翼龙(翼龙目)的舌器以及其摄食行为的启示
2015-12-16吕君昌
吕君昌
中国地质科学院地质研究所, 国土资源部地层与古生物重点实验室, 北京 100037
宽颌辽西翼龙(翼龙目)的舌器以及其摄食行为的启示
吕君昌
中国地质科学院地质研究所, 国土资源部地层与古生物重点实验室, 北京 100037
在翼龙类不同的类群中其摄食行为是不同的, 从其牙齿形状以及石化的胃容物来判断, 其摄食行为包括食鱼类、食昆虫类、滤食水中小的水生生物、食螃蟹和蜗牛等带壳的动物, 以及食水果类等。由于生存的需要, 它们占据不同的生态灶, 决定了其食物来源的不同。本文描述了宽颌辽西翼龙一件几乎完整、保存完好的与下颌在一起的舌器化石标本, 并且它与现生蜥蜴类变色龙的舌器对比。它长的舌突与运用其舌的外翻来摄取食物的现生蜥蜴类-变色龙相类似, 从而暗示了辽西翼龙可能具有类似的、用舌获取食物的行为, 加上其独特的牙齿形态, 进一步提出辽西翼龙是食昆虫类而不是食鱼的翼龙。
舌器; 辽西翼龙; 翼龙目; 中国
Pterosaurs, a group of Mesozoic flying reptiles, have varying feeding-behaviors, judging from their dentitions and fossilized stomach contents (Wellnhofer, 1991). At present, about 50 genera belonging to 12 families of pterosaurs have been discovered in China, and make up almost half the number of known pterosaurs in the world. More and more species will continue to be discovered with the further research. China has become the richest country with pterosaur fossils discovered in the world (Lü et al., 2013). During the last 10 years, more than 20 new species of pterosaurs have been found in north-east China and this rate of discovery shows little signs of slowing. Among them, there are two important discoveries: One is the discovery of Darwinopterus (Lü et al., 2010a; Lü, 2010a), an evolutionarily intermedi-ate form that bridges the gap between the two main groups of pterosaurs, and it provides strong evidence for an unusual type of ‘modular’ evolution. The other is the discovery of a female Darwinopterus with her egg preserved (Lü et al., 2011a), this resolved the gender identification problem in pterosaurs, and also the question of the significance of the cranial crest which confused scientists for more than a hundred years has been answered.
Although there are many beautifully preserved pterosaurs found from middle Jurassic and Early Cretaceous deposits (Lü et al., 2013), only several pterosaurs were reported with hyoids preserved, such as Liaoxipterus brachyognathus (Dong and Lü, 2005), Feilongus youngi (Wang et al., 2005), Darwinopterus robustodens (Xu et al., 2010; Lü et al., 2011b) and Kunpengopterus sinensis (Wang et al., 2010), however, the structures of hyoid apparatus were not described in detail, and much less their functions.
Compared with the other skeletal parts, even a partial hyoid apparatus of a pterosaur is rare. The holotype specimen of Liaoxipterus includes an almost completely preserved hyoid apparatus (Dong and Lü, 2005; Lü et al., 2008). Its long processus lingualis (processus entoglossus) is similar to that of the modern reptile Chameleon, which uses it for lingual feeding behavior. Thus, it is possible that Liaoxipterus practiced lingual feeding behavior, which would be expected if it was an insect-eating pterosaur.
The feeding behavior of pterosaurs was varied in different groups. Some pterosaurs are filter-feeding, such as Ctenochasma (Meyer, 1851), Gnathosaurus (Meyer, 1834), Huanhepterus (Dong, 1982) and Pterodaustro (Bonaparte, 1971; Chiappe et al., 2000; Codorniú and Chiappe, 2004), and used their numerous long, slender teeth to filter small aquatic organisms out of the water (Wellnhofer, 1991; Codorniú and Chiappe, 2004). The strong crushing teeth of dsungaripterids implies that fish is probably not the only food for them, and some shelly organisms such as mollusks, snails and crabs may have been part of their diets too (Young, 1973; Wellnhofer, 1991). Many pterosaurs were piscivorous, including Rhamphorhychus, Eudimophodon, and Pterodactylus, whose stomach contents are made up of the remains of fish (Wellnhofer, 1991). Some long beaked, toothless pterosaurs, such as Queztalcoatlus, may have used their slender beaks to probe for molluscs and arthropods in shallow water (Langston, 1981; Wellnhofer, 1991) or for carrion and small animals (Lawson, 1975; Witton and Naish, 2008). Insect-eating pterosaurs included Anurognathus ammoni (Wellnhofer, 1991).
Liaoxipterus (Fig. 1) was initially assigned to the Ctenochasmatidae, based on the lower jaw and the tooth number which is fewer than typical ctenochasmatid pterosaurs (Dong and Lü, 2005). However, further repairing the specimen shows that the tooth and the lower jaw end morphologies of Liaoxipterus are similar to those of Istiodactylus, thus it is reassigned to Istiodactylidae (Lü et al., 2006).

图1 和舌器保存在一起的辽西翼龙的下颌骨(A)和下颌前部的放大(B)Fig. 1 The lower jaw of Liaoxipterus with a hyoid apparatus preserved (A) and close up of the anterior portion of the lower jaw (B)
The hyoid apparatus supports and moves the tongue and, to a lesser degree, the larynx (Romer, 1956). While the hyoid apparatus is informative with respect to the function of the tongue, and therefore in the inference of diet, none of these interpretations of pterosaur diet rely on the anatomy of the hyoid apparatus because, compared with the other skeletal parts, remains of the hyoid apparatus in pterosaurs are rare. Therefore, the almost complete hyoid apparatus of Liaoxipterus described here is particularly significant.
1 Description
The hyoid apparatus is derived from the hyoid arch and the two succeeding visceral arches (Romer, 1956). It lies in the floor of the mouth and pharynx and is associated with the musculature of that region. In derived lizards, it consists of an unpaired body, the corpus, tapering in a cranial direction into a narrower process, the processus lingualis (or processus entoglossus), which varies in length and is bound into the posterior part of the tongue, and three pairs of horns connected laterally in various ways with the corpus. The anterior pair is called the cornu hyale, the middle pair is the cornu branchiale I, and the posterior pair is the cornu branchiale II (Fürbringer, 1922; Colbert, 1945).
In modern reptiles, such as Anolis, the hyoid apparatus is entirely cartilaginous, except for the ceratobranchials I (Bels, 1990). Because the reptilian hyoid elements are relatively small, separate from major skeletal structures, and often unossified, their fossilforms, if are found, are consisted of a single pair of elements, which, by analogy with recent forms, are reasonably identified as first ceratobranchials (Romer,1956). Thus it is inferred that the two long, slender posteriorly extended branches of the hyoid apparatus in Liaoxipterus are ceratobranchials I and its ceratobranchials II are not preserved.
The hyoid apparatus of Liaoxipterus (Figs.1A, 2) is a Y-shaped group of small bones. The length of the corpus including the hypohyal or lingual process is 26.3 mm, and the diameter of the corpus is 1.27 mm. The whole length of the hyoid apparatus (from the tip of the lingual process to the distal end of the ceratobranchial I) is 107.85 mm. The divarication of the ceratobranchial I is 21º (Lü et al., 2008). The highly elongated lingual process of the hyoid apparatus in Liaoxipterus is similar to that of Chameleon, while the elongated ceratobranchial I is similar to that of Vipera (Fig. 2), however, it is difficult to know whether the elongated ceratobranchials have a same function or not. Because the the ceratobranchial II in Liaoxipterus is cartilaginous, and it is not preserved as fossil element, the shape of the hyoid apparatus is different from that of the chameleon (Fig.2D). The tongue is connected to the relatively long entoglossal process (Bels, 1990), the elongated lingual processes between these two taxa are similar, thus they may play the same function.
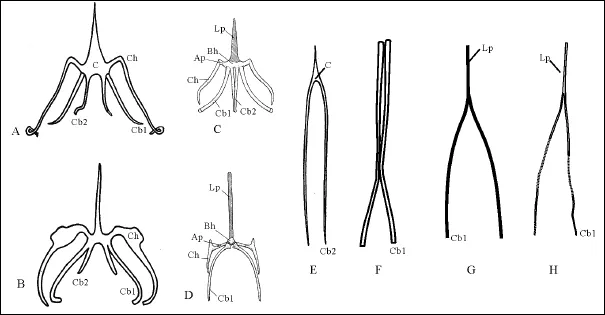
图2 现生爬行动物及其他翼龙类的舌器与辽西翼龙的舌器比较Fig. 2 A comparison of hyoids in other pterosaurs and the living reptiles with that of LiaoxipterusA-喙头目: 楔齿蜥属; B-有鳞目: Lacerta; C-蜥蜴; D-变色龙; E-有鳞目: 蝰蛇; F-壮齿达尔文翼龙; G-玩具翼龙属; H-辽西翼龙; A, B, E据Colbert, 1945; C, D据Kardong, 1998; 未按比例尺; 缩写: AP-前突起; Bh-基舌骨; C-主体, 包括下舌骨或舌内; Ch-cornu hyale角舌骨和上舌骨; Cbl-第一角鳃骨; Cb2-第二角鳃骨; Lp-舌突A-Rhynchocephalia: Sphenodon; B-Squamata: Lacerta; C-a lizard without a projectile tongue; D-the chameleon (the hyoid apparatus includes an elongated lingual process (LP) along which the tongue slides during launch; E-Squamata, Vipera; F-Darwinopterus robustodens (drawn from Lü et al., 2011b); G-Ludodactylus (Drawn from Frey et al., 2003); H-Liaoxipterus; A, B, E are from Colbert, 1945; C, D are from Kardong, 1998; Not to scale; Key to abbreviations: AP-anterior process; Bh-basihyal; C-corpus, including the hypohyal or entoglossal; Ch-ceratohyal and epihyal, cornu hyale; Cbl-ceratobranchial I, cornu branchiale I; Cb2-ceratobranchial II, cornu branchiale II; Lp-lingual process
2 Comparison and Discussion
The hyoid apparatus of pterosaurs is rarely preserved, especially for the complete ones. This is perhaps due to the poor ossification and fragility of the hyoid apparatus in pterosaurs. At present, only few cases of pterosaur hyoid have been reported (Wellnhofer and Kellner, 1991; Frey et al., 2003; Wang et al., 2005; Lü et al., 2011b), but they were only briefly mentioned.
The first ceratobranchials of hyoid apparatus in Liaoxipterus are fused at their distal ends, and then extends anteriorly forming a long lingual process (Ap), which is similar to that of Ludodactylus (Fig.2G, 2H). They are different from those of Darwinopterus robustodens (Fig. 2F) and Tapejara wellnhoferi (Wellnhofer and Kellner, 1991), where the first ceratobranchials of hyoid apparatus are separated from each other.
The tooth morphologies of pterosaurs from western Liaoning and its surrounding areas indicate that various food-catching behaviors exist among them. The pterosaurs with long, pointed teeth, such as Feilongus (Wang et al., 2005), Boreaopterus (Lü and Ji, 2005), Zhenyuanopterus (Lü, 2010b), Liaoningopterus (Wang and Zhou, 2003), Gegepterus (Wang et al., 2007) and Fenghuangopterus (Lü et al., 2010b) are clearly fish-eating pterosaurs. The toothless forms, such as tapejarid pterosaurs, Sinopterus (Wang and Zhou, 2003; Li et al., 2003), Huaxiapterus (Lü and Yuan, 2005; Lü et al., 2006) may also be meat-eating pterosaurs or fruit-eating as proposed by Wellnhofer and Kellner (1991) for Tapejara, although there is no direct evidence to support it. Darwinopterus is regarded to feed its contemporaneous fliers (Lü et al., 2010a), which include pterosaurs (Lü et al., 2006; Lü,2009), feathered maniraptorans (Czerkas and Yuan 2002; Zhang et al. 2002; Xu et al. 2009) and gliding mammals (Meng et al., 2006). The broad mouth and peg-like teeth in Jeholopterus and Dendrorhynchoides (=Dendrorhynchus) curvidentatus (Ji and Ji, 1998; Ji et al., 1999) indicate that these animals were probably insectivorous, just like Anurognathus and Batrachognathus (Wellnhofer and Kellner, 1991). The teeth of Liaoxipterus are rhombic with moderately sharp tips, they are different from the much longer, slenderer teeth with sharp tips of the fish-eating pterosaurs, such as Rhamphorhynchus (Wellnhofer, 1991), Feilongus (Wang et al., 2005), Boreaopterus (Lü and Ji, 2005), and Zhenyuanopterus (Lü, 2010b). The tooth morphology plus the complete hyoid apparatus of Liaoxipterus may provide the possible evidence to support its insect-eating behavior.
3 Conclusion
The long processus lingualis (processus entoglossus) of the hyoid apparatus (Colbert, 1945; Romer, 1956) indicates that Liaoxipterus may have practiced lingual feeding behavior, similar to the modern reptile Chamelon (Kardong, 1998). This may be an evidence to support lingual feeding behavior of pterosaurs, and it also implies that Liaoxipterus is an insect-eating pterosaur rather than a fish-eating pterosaur.
Acknowledgements: I am indebted to Professors Louis Jacobs (Southern Methodist University, USA) and Philip Currie (University of Alberta, Canada), who read the first draft of this paper and made constructive suggestions, and to Dr. Dale Winkler (Southern Methodist University, USA) for his help in different ways. This research was supported by grants from the National Natural Science Foundation of China (grant no.:41272022) and the China Geological Survey (grant no.12120114026801) to LÜ Jun-chang.
姬书安, 季强. 1998. 记辽宁一新翼龙化石(喙嘴龙亚目)[J]. 江苏地质, 22(4): 199-206.
李建军, 吕君昌, 堃张宝. 2003.记中国辽宁西部九佛堂组发现的中国翼龙一新种[J]. 古生物学报, 42(3): 442-447.
吕君昌. 2010. 达尔文翼龙的发现及其意义[J]. 地球学报, 31(2): 129-136.
吕君昌, 金幸生, 高春玲, 杜天明, 丁明, 盛益明, 魏雪芳. 2013.空中之龙-中国翼龙化石研究最新进展[M]. 杭州: 浙江科学技术出版社: 127.
汪筱林, 周忠和. 2003. 辽西早白垩世九佛堂组两种新的翼手龙类化石[J]. 古脊椎动物学报, 41(1): 34-41.
徐莉, 吕君昌, 常华丽, 魏雪芳, 张兴辽, 贾松海, 张纪明. 2010.中国东北辽西中侏罗世达尔文翼龙新标本[J]. 地球学报, 31(S1): 82.
References:
BELS V L. 1990. The Mechanism of dewlap extension in Anolis carolinensis (Reptilia: Iguanidae) with histological analysis of the hyoid apparatus[J]. Journal of Morphology, 206: 225-244.
BONAPARTE J F. 1971. Descripcion del craneo y mandibulas de Pterodaustro guinazui (Pterodactyloidea-Pterodaustriidae nov.), de la formacion Lagarcito, San Luis, Argentina[J]. Publicaciones del Museo Municipal de Ciencias Naturales, Mar del Plata, 1: 263-272.
CHIAPPE LM, KELLNER W A D, RIVAROLA S D, FOX M. 2000. Cranial morphology of Pterodaustro guinazui (Pterosauria: Pterodactyloidea) from the Lower Cretaceous of Argentina[J]. Natural History Museum of Los Angeles County, Contributions in Science, 483: 1-19.
COLBERT E H. 1945. The hyoid bones in protoceratops and in Psittacosaurus[J]. American Museum Novitates, 1301: 1-10.
CODORNIÚ L, CHIAPPE L M. 2004. Early juvenile pterosaurs (Pterodactyloidea: Pterodaustro guinazui) from the Lower Cretaceous of central Argentina[J]. Canadian Journal of Earth Sciences, 41: 9-18.
CZERKAS S A, YUAN C. 2002. An arboreal maniraptoran from northeast China[J]. In Feathered dinosaurs and the origin of flight 1 (ed. S. J. Czerkas): 63-95. Blanding, UT: The Dinosaur Museum Journal.
DONG Z M. 1982. On a new pterosaurian (Huanhepterus quingyangensis gen. et sp. nov.) from Ordos, China[J]. Vertebrata PalAsiatica, 20: 115-121.
DONG ZM, LÜ J C. 2005. A new ctenochasmatid pterosaur from the Early Cretaceous of Liaoning Province[J]. Acta Geologica Sinica, 79(2): 164-167.
FREY E, MARTILL DM, BUCHY M. 2003. A new crested ornithochierid from the Lower Cretaceous of northeastern Brazil and the unusual death of unusual pterosaur [M]//Buffetaut, E., and Mazin, J-M. (eds), Evolution and Palaeobiology of Pterosaurs. Geological Society, London, Special Publication, 217: 55-63.
FÜRBRINGER M. 1922. Das Zungenbein der Wirbeltiere insbesondere der Reptilien und Vögel[J]. Abhandl. Heidelberg Akad. Wiss., div. B 11: 1-164.
JI S A, JI Q. 1998. A new fossil pterosaur (Rhamphorhynchoidea) from Liaoning[J]. Jiangsu Geology, 22(4): 199-206(in Chinese with English abstract).
JI S A, JI Q, PADIAN K. 1999. Biostratigraphy of new pterosaurs from China[J]. Nature, 398: 573-574.
KARDONG K V 1998. Vertebrates: comparative anatomy, function, evolution[M]. Washington State University, Boston, Massachusett: 747.
LANGSTON J W. 1981. Pterosaurs[J]. Scientific American, 244(2): 122-136.
LI J J, LÜ J C, ZHANG B K. 2003. A new Lower Cretaceous sinopterid pterosaur from the Western Liaoning, China[J]. Acta Paleontologica Sinica, 42(3): 442-447 (in Chinese with English ab-stract).
LAWSON D A. 1975. Pterosaur from the Latest Cretaceous of West Texas: Discovery of the largest flying creature[J]. Science, 185: 947-948.
LÜ J C. 2009. A new non-pterodactyloid pterosaur from Qinglong County. Hebei Province of China[J]. Acta Geologica Sinica, 83: 189-199.
LÜ J C. 2010a. The discovery of Darwinopterus and its significance[J]. Acta Geoscientica Sinica, 31(2): 129-136.
LÜ J C. 2010b. A new boreopterid Pterodactyloid pterosaur from the Early Cretaceous Yixian Formation of Liaoning Province, Northeastern China[J]. Acta Geologica Sinica, 84(2): 241-246.
LÜ J C, JI Q. 2005. A new ornithocheirid from the Early Cretaceous of Liaoning Province, China[J]. Acta Geologica Sinica, 79(2): 157-163.
LÜ J C, YUAN C X. 2005. New tapejarid pterosaur from western Liaoning, China[J]. Acta Geologica Sinica, 79(4): 453-458.
LÜ J C, JI S A, YUAN C X, JI Q. 2006. Pterosaurs from China[M]. Beijing: Geological Publishing House: 147.
LÜ J C, XU L, JI Q. 2008. Restudy of Liaoxipterus (Istiodactylidae: Pterosauria), with comments on the Chinese istiodactylid pterosaurs[J]. Zitteliana, B28: 229-241.
LÜ J C, UNWIN D M, JIN X S, LIU Y Q, JI Q. 2010a. Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull[J]. Proceedings of the Royal Society B, 227: 383-389.
LÜ J C, FUCHA X H, CHEN J M. 2010b. A New Scaphognathine Pterosaur from the Middle Jurassic of Western Liaoning, China[J]. Acta Geoscientica Sinica, 31(2): 263-266(in Chinese with English abstract).
LÜ J C, UNWIN D M, DEEMING D C, JIN X S, LIU Y Q, JI Q. 2011a. An egg-adult association, gender, and reproduction in pterosaurs[J]. Science, 331: 321-324.
LÜ J C, XU L, CHANG H L, ZHANG X L. 2011b. A new darwinopterid pterosaur from the Middle Jurassic of western Liaoning, Northeastern China and its ecological implications[J]. Acta Geologica Sinica, 85(3): 507-514.
LÜ Jun-chang, JIN Xing-sheng, GAO Chun-ling, DU Tian-ming, DING Ming, SHENG Yi-ming, WEI Xue-fang. 2013. Dragons of the skies-Recent advances on the study of pterosaurs from China[M]. Hangzhou: Zhejiang Science and Technology Press: 127(in Chinese).
MENG J, HU Y M, WANG Y Q, WANG X L, LI C K. 2006. A Mesozoic gliding mammal from northeastern China[J]. Nature, 444: 889-893.
MEYER H. 1834. Gnathosaurus subulatus, ein saurus aus dem lithographischen Schiefer von Solenhofen[J]. Museum Senckenbergianum, 1(3): 1-8.
MEYER H. 1851. Ctenochasma roemeri[J]. Palaeontographica, 2: 81-83.
ROMER A S. 1956. Osteology of the Reptiles[M]. Chicago·Illinois: The University of Chicago Press: 772.
WANG X L, ZHOU Z H. 2003. Two new pterodactyloid pterosaurs from the Early Cretaceous Jiufotang Formation of western Liaoning, China[J]. Vertebrata PalAsiatica, 41(1): 34-41(in Chinese with English abstract).
WANG X L, KELLNER A W A, ZHOU Z H, CAMPOS D A. 2005. Pterosaur diversity and faunal turnover in Cretaceous terrestrial ecosystems in China[J]. Nature, 437: 875-879.
WANG X L, KELLNER A W A, ZHOU Z H, CAMPOS D A. 2007. A new pterosaur (Ctenochasmatidae, Archaeopterodactyloidea) from the Lower Cretaceous Yixian Formation of China[J]. Cretaceous Research, 28: 245-260.
WANG X L, KELLNER A W A, JIANG S X, CHENG X, MENG X, RODRIGUES T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China Anais da[J]. Academia Brasileira de Ciências, 82(4): 1045-1062.
WELLNHOFER P. 1991.The illustrated encyclopedia of pterosaurs[M]. London: Salamander Books: 191.
WELLNHOFER P, KELLNER A W A. 1991. The skull of Tapejara wellnhoferi Kellner (Reptilia, Pterosauria) from the Lower Cretaceous Santana Formation of the Araripe Basin, Northeastern Brazil[J]. Mitt. Bayer. Staatsslg. Paläont. Hist. Geol., 31: 89-106.
WITTON M P, NAISH D. 2008. A Reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology[J]. Plos ONE, 3(5): doi:10.1371/journal.pone.0002271.
XU Li, LÜ Jun-chang, CHANG Hua-li, WEI Xue-fang, ZHANG Xing-liao, JIA Song-hai, ZHANG Ji-ming. 2010. A new specimen of Darwinopterus from the Middle Jurassic of western Liaoning, northeastern China[J]. Acta Geoscientica Sinica, 31(S1): 82-82(in Chinese with English abstract).
XU X, ZHAO Q, NORELL M, SULLIVAN C, HONE D, ERICKSON G, WANG X, HAN F, GUO Y. 2009. A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin[J]. Chinese Science Bulletin, 54: 430-435.
YOUNG C C. 1973. Pterosaurs from Wuerho[J]. Memoirs of the Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica, 11: 18-35(in Chinese).
ZHANG F C, ZHOU Z H, XU X, WANG X. 2002. A juvenile coelurosaurian theropod from China indicates arboreal habits[J]. Naturwissenschaften, 89: 394-398.
The Hyoid Apparatus of Liaoxipterus Brachycephalus (Pterosauria) and Its Implications for Food-catching Behavior
LÜ Jun-chang
Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037; Key Lab of Stratigraphy and Paleontology, Ministry of Land and Resources of China, Beijing 100037
The feeding behavior of pterosaurs was varied in different groups. Judging by the tooth morphologies and fossilized stomach contents of pterosaurs, the feeding habits included fish-eating, insect-eating, filtering small aquatic organisms, eating shelled crabs and snails, and fruit-eating. Because of the need for survival, they occupied different ecological niches, which determined their different food sources. Herein described is an almost complete, well-preserved hyoid apparatus of Liaoxipterus brachyognathus in comparison with the hyoid apparatus of the modern lizard Chameleon. The long processus lingualis (processus entoglossus) is similar to that of the modern lizard Chameleon, which captures prey by tongue, implying that Liaoxipterus might share a similar lingual feeding behavior. This phenomenon, plus its special tooth morphology, further suggests that it was an insect-eating rather than fish-eating pterosaur.
hyoid apparatus; Liaoxipterus; pterosauria; China
Q915.2; Q915.86
A
10.3975/cagsb.2015.03.13
本文由国家自然基金(编号: 41272022)和中国地质调查局地质调查项目(编号: 12120114026801)联合资助。
2015-03-09; 改回日期: 2015-04-01。责任编辑: 闫立娟。
吕君昌, 男, 1965年生。研究员, 博士生导师。主要从事中生代爬行动物(尤其是恐龙类和翼龙类)及其地层学研究。通讯地址: 100037, 北京市西城区百万庄大街26号。电话: 010-68999707。E-mail: Lujc2008@126.com。
