小鼠大脑缺氧后eIF4G在脑组织中的表达
2015-12-15高艳斌张林庆可李家宏强陈志明长春市公安局刑警支队吉林长春30000
高艳斌 张林庆 马 可李家宏 任 强陈志明(长春市公安局刑警支队,吉林 长春 30000)
小鼠大脑缺氧后eIF4G在脑组织中的表达
高艳斌张林庆马可1李家宏任强2陈志明2(长春市公安局刑警支队,吉林长春130000)
〔摘要〕目的探讨小鼠颅脑缺氧后eIF4G随时间变化基因转录及蛋白表达改变。方法将50只大鼠随机分为10组,采用手术结扎右侧颈总动脉,分别于术后2、8、24 h、1、3、6、9、12及16 d处死,应用RT-PCR检测eIF4G的表达变化。结果eIF4G在脑缺氧2 h后表达增高,且随时间延长表达持续增高,差异有统计学意义。结论eIF4G在小鼠脑缺氧后增高,说明蛋白翻译过程在脑缺氧后发挥重要作用。
〔关键词〕脑缺氧; eIF4G
1吉林大学第一医院儿科急诊2吉林大学基础医学院法医教研室
第一作者:高艳斌(1976-),男,副主任法医师,主要从事法医学研究。
翻译是细胞内蛋白质合成过程中的关键步骤,在细胞增殖、分化、程序性死亡及适应环境方面均分挥着重要作用〔1〕。对于翻译调节因子的表达调控可显著延缓衰老,而其中尤以对翻译起始因子4G(eIF4G)的调节所表现的生物学行为改变最为显著〔2〕。对eIF4G的调控为细胞启动蛋白合成的重要限速步骤,其在脑缺氧后的表达变化却鲜有报道。本研究对eIF4G表达与小鼠脑缺氧后不同时间段的关系进行研究。
1 材料与方法
1. 1动物分组及模型建立雄性昆明系小鼠50只,随机分为正常对照及脑缺氧组。脑缺氧组分别于术后2、8、24 h、1、3、6、9、12及16 d处死,共10组。处死后脑组织迅速置于液氮中,5 min后取出保存于-80℃冰箱中。采用手术结扎右侧颈总动脉,用10%水合氯醛(0. 35 g/kg体重)腹腔内注射麻醉,剪去颈部皮毛,消毒皮肤后,沿中线切开皮肤,暴露右侧颈总动脉后,结扎后缝合皮肤,置鼠笼喂养。正常对照组仅切开颈部皮肤,但并不结扎动脉。
1. 2方法
1. 2. 1 RNA提取称取100 mg脑组织用1 ml Trizol进行裂解;将Trizol裂解液转入EP管中,15℃~30℃放置5 min;加0. 2 ml氯仿后用力震荡15 s,室温下放置2~3 min,12 000 r/min(2℃~8℃)离心15 min。取上层水相置于新EP管中,加0. 5 ml异丙醇,15℃~30℃放置10 min,12 000 r/min(2℃~8℃)离心10 min。加1 ml 75%乙醇进行洗涤,涡旋混合,7 500 g(2℃~8℃)离心5 min,弃上清;让沉淀的RNA在室温下自然干燥; 用RNAse-free水溶解RNA沉淀。采用RT-PCR试剂盒进行逆转录PCR。
1. 2. 2 Real-time PCR检测取0. 2 ml薄壁PCR管,分别编号。向各管中加入2×qPCR TaqMix 12. 5 μl,10 μmol/L各基因正反向引物混合物0. 5 μl。管中不加模板用作阴性对照。各管补加水至25 μl。混匀置于荧光定量PCR仪中。本研究所用引物序列见表1。
1. 3统计学分析采用SPSS3. 0统计软件包进行统计,应用单因素方差分析和各组均数t检验处理。
2 结果
eIF4G在颅脑缺氧后表达持续增高,在正常颈动脉未结扎脑组织中,eIF4G表达量较稳定;在颅脑缺氧2 h后表达增高,差异有统计学意义,且随缺氧时间延长后持续增高,缺氧24 h后达峰值。见表2。
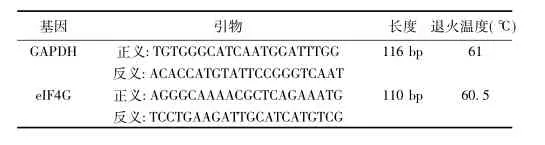
表1 Real-time PCR所用引物序列
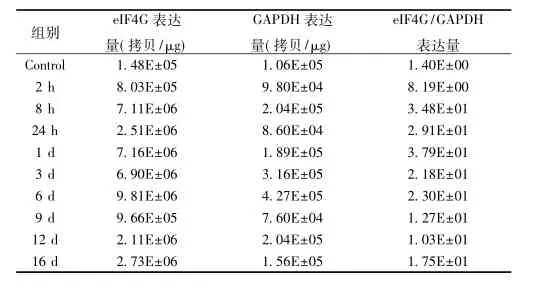
表2 eIF4G在小鼠大脑缺氧后脑组织转录表达变化(x±s)
3 讨论
蛋白翻译调节对于细胞生长及适应环境方面均具有重要意义〔3〕。翻译过程包括三个步骤,翻译起始、翻译延伸及翻译终止。尽管对?翻译各个阶段的调节均对细胞生物学行为具有重要影响,但对翻译起始的调节却是对翻译过程调节的重要限速步骤〔4〕。翻译起始过程既包括蛋白与RNA的相互作用,也包括蛋白与蛋白的相互作用,许多翻译起始因子参与上述过程,但eIF4G为此过程中最重要的调节蛋白,其可作为核糖体蛋白及mRNA的重要桥接蛋白〔5〕。尽管eIF4G为翻译过程中最重要的调节蛋白,其表达缺陷可导致细胞死亡,适当减少eIF4G的表达可延缓衰老,并延长生命。另外,其对于细胞适应应激过程亦发挥着重要作用,从而在细胞缺氧耐受及肿瘤发生过程中发挥重要作用〔6,7〕。
脑组织的长期慢性缺氧与老年患者神经退行性变的发生密切相关,而翻译过程的调节异常与其发生发展的关系却鲜有报道。在本研究中,本文颅脑缺氧后,翻译过程异常不仅出现在急性期,在脑组织缺氧后长期持续存在,提示对翻译异常的纠正可为临床治疗及预防脑神经退行性变提供新的理论基础。
4参考文献
1 El Kadmiri N,Slassi I,El Moutawakil B,et al.Glyceraldehyde-3-phosphate dehydrogenase(GAPDH)and Alzheimer's disease〔J〕.Pathol Biol(Paris),2014; 62(6): 333-6.
2 Alves da Costa C,Sunyach C,Pardossi-Piquard R,et al.Presenilin-dependent gamma-secretase-mediated control of p53-associated cell death in Alzheimer's disease〔J〕.J Neurosci,2006; 26: 6377-85.
3 Lujambio A,Akkari L,Simon J,et al.Non-cell-autonomous tumor suppression by p53〔J〕.Cell,2013; 153: 449-60.
4 Munoz-Espin D,Canamero M,Maraver A,et al.Programmed cell senescence during mammalian embryonic development〔J〕.Cell,2013; 155: 1104-18.
5 Rubbi CP,Milner J. Disruption of the nucleolus mediates stabilization of p53 in response to DNA damage and other stresses〔J〕.EMBO J,2003; 22: 6068-77.
6 Botchkarev VA,Flores ER. p53/p63/p73 in the epidermis in health and disease〔J〕.Cold Spring Harb Perspect Med,2014: 4(8): pii: a015248.
7 Tollini LA,Jin A,Park J,et al.Regulation of p53 by Mdm2 E3 ligase function is dispensable in embryogenesis and development,but essential in response to DNA damage〔J〕.Cancer Cell,2014; 26: 235-47.
〔2014-03-17修回〕
(编辑袁左鸣)
通讯作者:陈志明(1977-),男,讲师,主要从事法医病理学研究。
〔文章编号〕1005-9202(2015)20-5750-02;
doi:10. 3969/j. issn. 1005-9202. 2015. 20. 029
〔文献标识码〕A
〔中图分类号〕R74
