Combining Ability Analysis on Yield Traits of Recombinant Inbred Lines in Hybrid Rice
2015-12-14YingjiangCAOShumeiYOUJiakuiZHENGKaifengJIANGTaoZHANGLiYANGQianhuaYANGXianqiWANJingLUOZhaoxiangLILeiGAO
Yingjiang CAO, Shumei YOU, Jiakui ZHENG, Kaifeng JIANG, Tao ZHANG, Li YANG, Qianhua YANG, Xianqi WAN, Jing LUO, Zhaoxiang LI, Lei GAO
Rice and Sorghum Research Institute, Sichuan Academy of Agricultural Sciences /Luzhou Branch of National Rice Improvement Center/Key Laboratory of Southwest Rice Biology and Genetic Breeding, Ministry of Agriculture, Deyang 618000, China
As one of the most important grain crops around the world,rice is widely planted in 30%of the total area for food crops and about 2/3 population feeds on it in China[1].At present, improving yield per unit area plays a crucial role in effectively alleviating the conflict between the decreasing cultivated area caused by industry construction, natural damage,ecological restoration, and the increasing demand for rice due to population growth. Optimum cross combinations can be obtained through combining ability analysis of various culti vars[2],and the contributions of parental characters to F1generation can be determined by genetic analysis[3-6]. During the growth period, male parent often contributes more to the traits of F1than female parent, but in such traits,kernel length, kernel width and 1 000-grain weight, female parent has more contribution rate. Several important and widely planted materials from which many cultivars were derived were found in breeding practice, such as Minghui 63[7-10]. By 2010, a total of 34 cultivars derived from Minghui 63 as male parent had been approved by Provincial or National Crop Approval Committee,including four approved byNational Crop Approval Committee. In addition, 543 new restorer lines were bred using Minghui 63 as the core parent, and then 922 combinations derived from the 543 new restorer lines have already been examined and approved by Provincial or National Crop Approval Committee[10]. Then the questions were whether these materials contain high combining ability genes, and how to isolate them. To find the answers, Qi et al.[11], from Huazhong Agricultural University,constructed some combinations using Zhong 3 and HB 522, and then discovered 56 QTLs determining the general combining ability of maize agronomic traits and 21 QTLs controlling the specific combining ability in these combinations. Qu et al.[12]by crossing the chromosome segments substitution lines derived from 97 B and 9311, with three two-line GMS lines Hua 893S, Hua 88S and Aipei 64S, obtained a diallel cross population, and found that using the population could locate the QTLs controlling the combining ability of main agronomic traits of rice. In the present study, a permanent F2population was first obtained using a three-line restorer Luhui 8258 which had high combining ability as core parent, and then diallel cross scheme was made by testcrossing three-line CMS lines with the individuals of the population, to analyze the combining ability of main agronomic traits of rice, aiming at providing reference for discovering and locating gene loci for high combining ability.
Materials and Methods
Materials
A populatio n of 140 recombinant inbred lines at F8generation were obtained after seven successive generations of self-pollination using single seed descent (SSD)method from the F2hybrids of three-line restorers Luhui 8258×Yanghui 34. NC Ⅱdesign was adopted to conduct diallel cross through crossing three different types of cyto-plasmic male sterile (CMS)lines (Gang 46A,Ⅱ-32A and Lu 98A)with the 140 inbred lines obtained above and their parents Luhui 8258 and Yanghui 34,then the seeds of 426 F1materials were obtained and tested later. Eight cultivars, which were bred from Luhui 8258 with high combining ability, had already been examined and approved by Provincial or National Crop Approval Committee until March of 2014(Table 1).
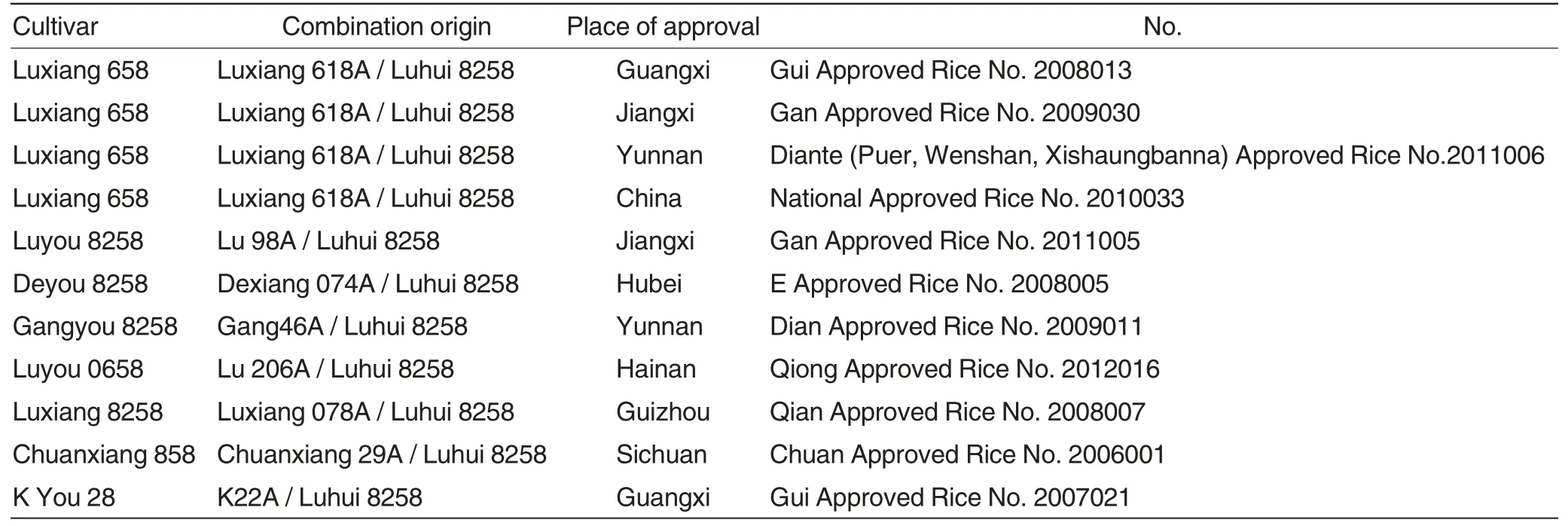
Table 1 Eight cultivars derived from the restorer line Luhui 8258
Methods
Randomized block design was adopted to conduct field experiments at Deyang Base of Rice and Sorghum Research Institute, Sichuan Academy of Agricultural Sciences and Suining Base of Dayin County, Suining City of Sichuan Province in 2012.Seeds were sown on March 25, and the seedlings were transplanted on May 15 at Deyang Base. At Suining Base seeds were sown on April 1, and the seedlings were transplanted on May 15.Three repetitions were set for each treatment. The rows were 33.3 cm long each and spaced 19.8 cm.The 21 plants for each material were planted in three rows, with 7 plants in each row,under conventional field management pest control.
Determination index
Three plants those grew well and were similar in size were selected from every plot at maturity to measure the yield per plant, effective panicle number per plant, seed setting rate, filled grain number per panicle and 1 000-grain weight of every material. In detail, the rice yield per plant was calculated by dividing the total seed weight of three plants by three. The effective panicle number per plant was calculated by dividing the number of panicles with more than five filled grains of three plants by three.Seed setting rate was equal to the total number of filled grains divided by the total number of glumous flowers. Filled grain number per panicle was equal to the total number of filled grains of three plants divided by the total number of effective panicles of the three plants. 1 000-grain weight was equal to the weight 1 500 filled grains (500 from each plant)multiplied by two thirds.
Analysis on combining ability
According to the studies of Liu et al.[13]and Mo[14],combining ability effect was estimated by the fixed model(model 1), and variance components and some related genetic parameters were estimated by the random model(model 2).
Normality test on 142 rice materials was performed using D’Agostino method[15]. Under the principle of D’Agostino test,the critical value of Y for a population of normal distribution increases as samples number increasing. According to the percentile points of statistic Y in D’Agostino test, the critical value of Y is -2.000 if n=150 and the test level is 0.05. Whether the combining ability of the five traits fit a normal distribution was judged depending on their Y values(Y≤-2.000).
Data analysis
Excel 2007 was used for dataprocessing.DPS 7.05 was adopted for combining ability analysis and normal distribution test.
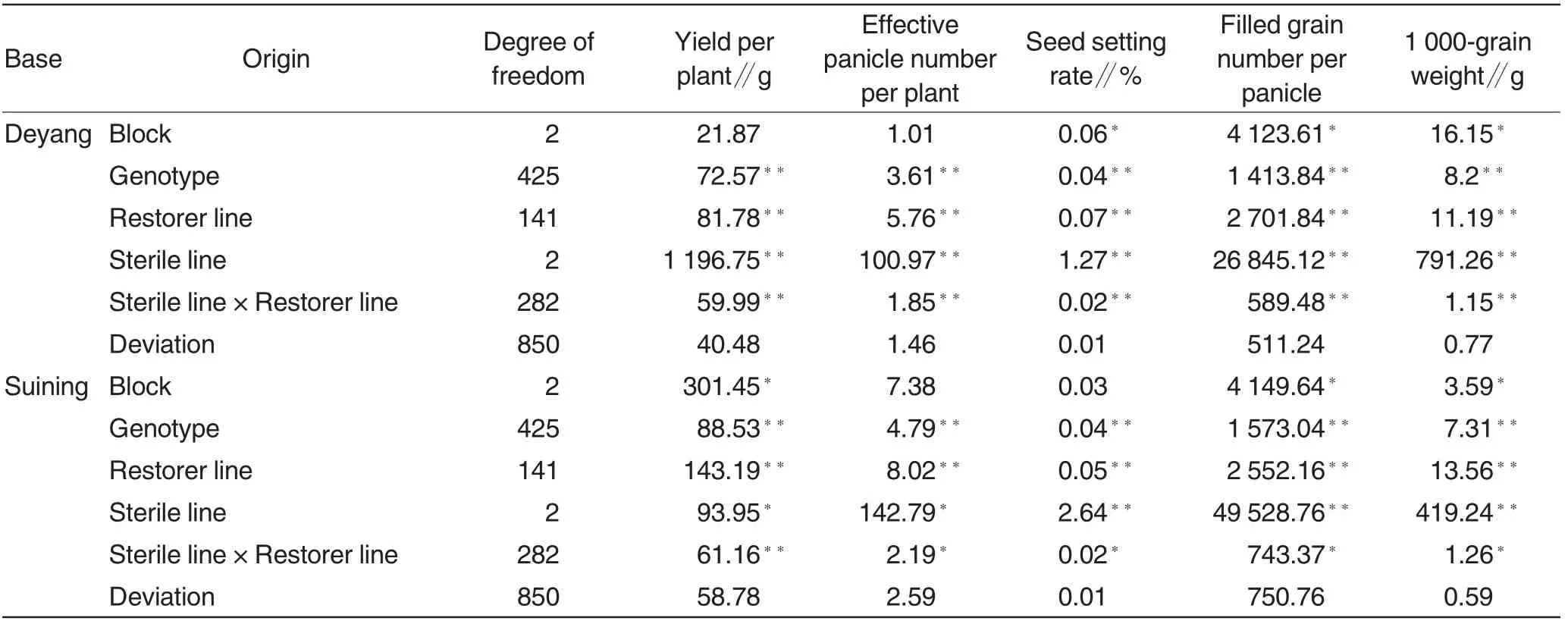
Table 2 Variance analysis on five yield traits at Deyang and Suining bases

Table 3 Genotype variance of combining ability and heritability of the 5 traits
Results and Analysis
Variances of yield traits and combing ability
The variance analysis on the five yield traits of 426 combinations revealed that extremely significant differences in the yield traits existed among these combinations at Deyang and Suining bases, indicating that the combinations were genetically different(Table 2). Further study on the variances of yield traits of sterile lines,restorer lines and sterile lines× restorer lines at Deyang and Suining bases indicated that these traits were affected by additive and non-additive effects of genes based on their extremely significant difference.
Genotype variance and heritability of combining ability of yield traits
Except for yield per plant at Deyang Base, the genotype variance for the general combining ability of every yield trait ranged from 73.65% to 100%, which demonstrated that the additive effects of parental genes weredominant during the formation of F1hybrids(Table 3).Judging from genotype variance of general combing ability,the contribution rates of the male parent in effective panicle number per plant and filled grain number per panicle at two sites were higher than 50%,indicating that the two traits were mainly controlled by the male parent.However, the female parent’s genes contributed more than male parent’genes to the seed setting rate of F1hybrids, because the contribution rate of the female parent was above 50%at two sites.
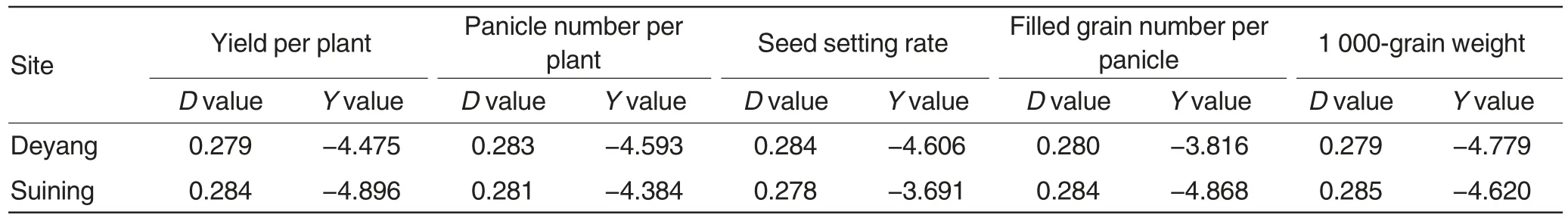
Table 4 Normality test on general combining ability of five yield traits
The results also revealed that 1 000-grain weight was little affected by environmental factors and thus could be stably inherited to later generations,as its broad and narrow heritability were both higher than 70% at two sites. In contrary, the other traits with low heritability could be easily affected by environment(Table 3).
Distribution of general combining ability of yield traits
Based on the value and variation range of combining ability, the 142 materials were divided into 13 groups.As shown from Fig.1 to Fig.5, the maximum distribution frequency of general combing ability of the yield traits ranged from 20 to 29. Meanwhile, at Deyang Base the yield per plant had the largest combining ability,while at Suining Base the effective panicle number per plant had the largest combining ability, and their class mid-values were 1.44 and -1.98 respectively.Only 20.42%of all tested materials, namely 29 lines varied in this range.
The ratios of recombinant inbred lines with GCA positive effect at each site were calculated and described as follows.In detail,47.18% (67 lines)of the recombinant inbred lines at Deyang Base and 52.11%(74 lines)of the recombinant inbred lines at Suining Base showed GCA positive effect in yield per plant (Fig.1); 46.48% (66 lines) of the recombinant inbred lines at Deyang Base and 47.18%(67 lines)of the recombinant inbred lines at Suining Base showed GCA positive effect in panicle number per plant (Fig.2); 43.66% (62 lines) of the recombinant inbred lines at Deyang Base and 52.11% (74 lines) of the recombinant inbred lines at Suining Base showed GCA positive effect in panicle number per plant (Fig.3); 50.70% (72 lines) of the recombinant inbred lines at Deyang Base and 52.82%(75 lines)of the recombinant inbred lines at Suining Base showed GCA positive effect in filled grain number per plant (Fig.4);49.30% (70 lines) of the recombinant inbred lines at Deyang Base and 59.43% (78 lines) of the recombinant inbred lines at Suining Base showed GCA positive effect in 1 000-grain weight(Fig.5).
Normality test on the general combining ability of five yield traits in permanent F2 population
As shown in Table 4, Y values of the five traits was between-4.896 and-3.691 at two sites, lower than the critical value-2.000,indicating that the general combining ability of five traits had a normal distribution in permanent F2population.
Correlation coefficient of combining ability of yield traits
The correlation coefficient of the general combining ability of the five traits at the two bases ranged from-0.65 to 0.46 (Table 5). Twelve pairs of the traits at the two bases were positively correlated in general combining ability, among which, four pairs at Deyang and five pairs at Suining shared extremely significant positive correlation. However, the general combining ability had negative correlations in eight pairs of the traits at the two bases, among which four pairs at Deyang and two pairs at Suining had extremely significant negative correlation.
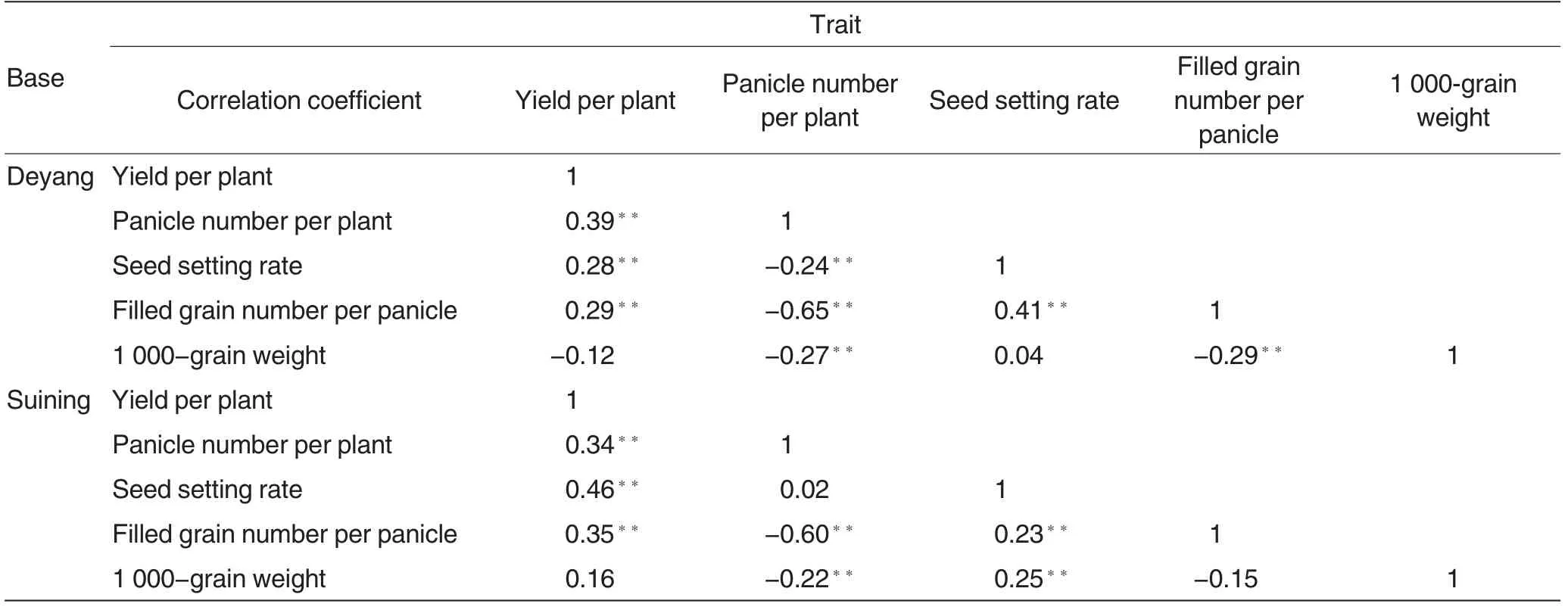
Table 5 Correlation coefficient among the general combining ability of five traits
Conclusion and Discussion
In 1980s,Zhou et al.[18]and Huang et al.[19]started breeding based on the theory of combining ability which was proposed by Sprague[16]and Griffing[17].In the present paper, the recombinant inbred lines were bred from Luhui 8258 and Yanghui 34 at Deyang and Suining bases to analyze the combining ability of rice. The results proved that significant or very significant correlation existed among the variances of general and specific combining abilities of five yield traits, revealing that the five traits were controlled by both additive and non-additive effects of genes. Meanwhile, the general combining ability variance of every parental trait was larger than their specific combining ability variance, indicating that their additive effects, not non-additive effects played a dominant role during the trait formation in their later generations.
In general, broad heritability reflects the effects of genetic variation and environment change.In this study,the broad and narrow heritability of four yield traits were below 50%, except that the broad and narrow heritability of 1 000-grain weight were both higher than 70%at two sites,suggesting all the traits except 1 000-grain weight, were vulnerable to environment impact, which agreed well with the findings of Jiang’s et al[6].
Correlation analysis proved that the combining abilities of yield traits(except 1 000-grain weight) and yield had an extremely significant positive correlation,with correlation coefficients ranging from 0.28 to 0.46. The results suggested that selecting parental materials with suitable traits is critical tobreeding target combinations with high combining ability.
Along with the development of molecular biotechnology, much progress has been made in locating the genes controlling target traits of crops by using molecular makers. In recent years, the combing ability of crops was even considered as a trait in some studies[11-12].This study proved that the general combining ability of five yield traits showed a normal distribution in recombinant inbred lines,right in line with inheritance of quantitative traits. So, the genes controlling rice general combining ability can be targeted by QTL mapping.
[1]ZHU DF (朱德峰),CHEN SH (程式华),ZHANG YP(张玉屏),et al.The production situation and restraint factors of rice around the world (全球水稻生产现状与制约因素分析)[J]. Sientia Agricultura Sinca (中国农业科学), 2010,(3): 474-479.
[2]TANG WB(唐文帮), XIAO YH(肖应辉),DENG HB (邓化冰), et al. Combining ability and utilization of C815S male sterile lines (水稻两用核不育系C815S的配合力及利用价值)[J]. Journal of Hunan Agricultural University (Natural Sciences)(湖南农业大学学报:自然科学版),2010,36(4):367-372.
[3]SONG Y (宋宇),ZOU XY (邹小云),HE HH (贺浩华), et al. Combining ability analysis of yield and related characters in three-line Indica hybrid rice(籼型三系杂交水稻产量及其相关性状的配合力分析)[J]. Acta Agriculturae Universitatis Jiangxiensis(江西农业大学学报),2004,26(5):719-725.
[4]YU SW(余守武),YI JH(尹建华),LIU YB(刘宜柏), et al.Study on breeding of three-way hybrid rice (Oryza sativa L.)III.Analysis on the combining ability and heritability of main agronomic traits of three-way hybrid rice in mid-late season cropping(三交水稻的育种研究Ⅲ.三交中晚稻主要农艺性状的配合力和遗传力分析)[J].Acta Agronomic Sinica(作物学报),2005,31(6):784-789.
[5]CAI JG (蔡巨广), FANG SR (方珊茹),ZHENG YM (郑燕梅),et al.Review on combing ability of hybrid rice (杂交水稻配合力的研究概况)[J]. Fujian Science and Technology of Rice and Wheat (福建稻麦科技),2006,24(3):44-47.
[6]JIANG KF(蒋开锋),ZHENG JK(郑家奎),ZENG DC (曾德初),et al.Primary study on combining ability of yield stability of hybrid rice(杂交水稻稳产性配合力初步研究)[J]. Chinese Journal of Rice Science(中国水稻科学),1998,12(3):134-138.
[7]TANG SY(汤圣祥),WANG XD(王秀东),LIU X(刘旭).Study on the renewed tendency and key backbone-parents of inbred rice cultivars(O.sativa L.)in China(中国常规水稻品种的更替趋势和核心骨干亲本研究)[J]. Scientia Agricultura Sinica (中国农业科学), 2012, 45(8):1455-1464.
[8]SI HM(斯华敏),FU YP(付亚萍),LIU WZ(刘文真),et al.Pedigree analysis of photoperiod-thermo sensitive genetic male sterile rice (水稻光温敏雄性核不育系的系谱分析)[J]. Acta Agronomica Sinica(作物学报),2012,38(3):394-407.
[9]LIU HN (刘怀年),WANG SQ (王世全),DENG QM (邓其明), et al.Critical section analysis of related characters of backbone parent Shuhui 527 in rice (水稻骨干亲本蜀恢527 产量相关性状关键区段分析)[J]. Journal of Agricultural Biotechnology (农业生物技术学报),2011,19(3):393-406.
[10]WU FX (吴方喜), CAI QH (蔡秋华),ZHU YS (朱永生), et al. Application and innovation of Indica hybrid restoring-line Minghui 63(籼型杂交稻恢复系明恢63 的利用与创新)[J]. Fujian Journal of Agricultural Sciences (福建农业学报),2011,26(6):1101-1112.
[11]QI HH, HUANG J, ZHENG Q, et al. Identification of combining ability loci for five yield-related traits in maize using a set of testcrosses with introgression lines [J].Theor Appl Gene,2013,126:369-377.
[12]QU Z,LI LZ,LUO JY, et al.QTL mapping of combining ability and heterosis of agronomic traits in rice backcross recombinant inbred lines and hybrid crosses[J].Plos One,2012,7(1):1-10.
[13]LIU LF (刘来福), MAO SX (毛盛贤),HUANG YZ(黄远樟).Crop quantitative genetics(作物数量遗传学)[M].Beijing:China Agriculture Press(北京:中国农业出版社),1984:250-259.
[14]MAO HD (莫惠栋). Combining ability analysis of mating pattern p×q (p×q 交配模式的配合力分析)[J].Jiangsu Agricultural Research (江苏农学院学报),1982,(3):51-57.
[15]LIANG XJ(梁小筠).Normality test(正态性检验)[M].Beijing: China Statistics Press (北京: 中国统计出版社),1997:105-114.
[16]SPRAGUE BA, TATUM LA. General vs specific combining ability in single crosses [J]. Agron J, 1942,(34): 923-934.
[17]GRIFFING JB.Concept of general and specific combining ability in relation to dialle crossing systems[J].Australian J Biol Sci,1956(9):463-493.
[18]ZHOU KD(周开达),NI HY(黎汉云),LI RR (李仁端), et al. Preliminary study on the heritability and combining ability of major characteristics in hybrid rice(杂交水稻主要性状配合力 遗传力的初步研究)[J].Acta Agronomica Sinica(作物学报),1982,(3):145-152.
[19]HUANG YX(黄耀祥),DONG QK(董群铠),ZHANG JY (张俊英),et al.Combining ability analysis of yield in different strains of rice parents cultivars (水稻不同株型亲本品种产量的配合力分析) [J]. Guangdong Agricultural Sciences(广东农业科学),1985,(6):1-5.
猜你喜欢
杂志排行

Agricultural Science & Technology的其它文章
- Evaluation on Suitability of Camellia sinensis Planting Based on GIS
- Light Quality-controlled Phytochemicals Biosynthesis in Vegetables and Fruits
- Cloning and Characterization of Phytochrome A Gene FaPHYA from Tall Fescue
- Genetic Analysis of Embryo Production Frequency in Wheat×Maize Cross
- The Application Effects of Truly Biodegradable Mulch in Potato Farmlands
- The Analysis and Prospect of Development of Fresh Cut Flower Industry Based on the Patent Analysis