Genetic Analysis of Embryo Production Frequency in Wheat×Maize Cross
2015-12-14KunLIUBoNINGHongshengLIShaoxiangLIJianGUHongZHAOMujunYANG
Kun LIU, Bo NING, Hongsheng LI, Shaoxiang LI, Jian GU, Hong ZHAO, Mujun YANG
Institute of Food Crops, Yunnan Academy of Agricultural Sciences, Kunming 650205, China
Zenkteler and Laurie developed a technology to induce the wheat haploid embryos with maize pollen[1-2], which has three following advantages: first, since maize is insensitive to mating suspension gene Kr in wheat,the embryo production frequency (or embryo formation frequency, which refers to the number of obtained wheat haploid embryos per 100 pollinated florets) of wheat × maize cross is higher than that of wheat × Hordeum bulbosum L.cross by tens and even hundreds of times[3-4]. Second, the production of haploid embryos has no strict selectivity for wheat genotypes, all genotypes of wheat can be used to produce haploid embryos[5-8]. In particular, for genotypes that cannot produce callus or plants by anther culture,higher embryo and plant production frequencies are shown in wheat × maize crosses[9]than those in wheat anther culture.Third, no albino seedlings or somatic mutation are produced. This technology makes it easier to produce stable homozygous haploids, which is of great value for the application and genetic research of breeding. Currently,there are great differences in haploid embryo production frequency among different genotypes of wheat[7,10-13].The lower EPF is less than 10%, while the higher EPF is more than 50%,seriously restricting the application of the technology. Since the formation of wheat haploid embryos is the premise to produce haploid and doubled haploid seedlings, the overall stabilization and improvement of haploid embryo production frequency of wheat materials has become a hotspot in domestic and foreign researches. The haploid embryo production frequency in wheat × maize cross is affected by many factors. Previous studies mainly focused on maize genotype[10-11,14],pollination method[15], wheat floret po sition and development stage during pollination[16-17], treatment method of 2, 4-D after pollination[16,18]and culture conditions and culture nutrition formulation of cut spikes[12,18]. But there are rare reports on effects of genetic factors on haploid embryo production fre-quency. In this study, a DH population derived from wheat materials with different EPFs was used as the tested material, the genetic characteristics of haploid embryo production frequency in wheat × maize cross were studied with mixed major gene and polygene inheritance model so as to understand the influence of genetic factors on haploid embryo production frequency,thereby providing a scientific basis for the improvement of haploid embryo production frequency in wheat×maize cross.
Materials and Methods
Materials
Wheat DH population The wheat DH population derived from C49S-87/01Y1-1069 was constructed using the haploid embryo induction technology by the Institute of Food Crops,Yunnan Academy of Agricultural Sciences in 2006. The multiplication, screening and identification were performed in summer, 2007. The DH plants with morphological variation among different years were eliminated, and finally total 250 DH lines were retained. No segregation distortion was found in the DH population when examined with the known powdery mildew-resistant gene Pm21 and yellow rust-resistant gene Yr26 from 01Y1-1069, suggesting that the DH population had good genetic representation. The female parent C49S-87 was a photoperiodsensitive genetic male sterile line bred by the Institute of Special Crops,Chongqing Academy of Agricultural Sciences in 1994,and the male parent 01Y1-1069 was a wheat cultivar bred by the Institute of Food Crops,Yunnan Academy of Agricultural Sciences. In the haploid embryo induction tests by wheat × maize cross in 2007-2009,the haploid embryo production frequencies of C49S-87 ranged from 10% to 15% , and of 01Y1-1069 ranged from 40%to 45%.
Maize material The maize cultivar Baitiannuo, with good ability in inducing haploid embryo was bred by the maize breeding center of Institute of Food Crops, Yunnan Academy of Agricultural Sciences through years of tests. It was used as the pollen donor for wheat×maize cross in this study.
Methods
Haploid embryos induced by wheat × maize cross The 250 DH lines and their parents were planted in the net house of Yunnan Academy of Agricultural Sciences on May 18th,2008. The wheat and maize materials were sowed dividedly in summer as described by Gu et al[19].The wheat DH materials and their parents were sowed once, and there were three rows for each of the wheat DH materials and their parents; while the maize materials were sowed at three different times respectively. After heading the wheat materials were emasculated artificially before anthesis. Among the 250 lines, total 138 wheat DH lines with heading time less than 5 d were selected. For each of the selected lines,total 6 spikes were emasculated,and the kept florets were not less than 150 to get more than 100 caryopses.The spikes were pollinated with maize pollen 2-3 d after emasculation[19],and cut down 24 h after pollination. And then, spikes were sprayed with 2, 4-D solution (100 mg/L) and cultured in vitro. The embryos were observed under a dissecting microscope after a 14-d culture. For each of the DH lines, the pollinated florets and obtained haploid embryos numbers were counted. The haploid embryo production frequency was calculated according to the following formula:
EPF(%)=Number of haploid embryos/Number of pollinated florets ×100.
Data statistics and analysis The EPFs of the DH population were analyzed using DPS.The genetic characteristics of EPFs were analyzed using the mixed major gene and polygene inheritance model proposed by Wang and Gai[20-21].Among the 46 mixed major gene and polygene models in the DH population,the distribution parameters of the relevant components in the mixed distribution were estimated by Likelihood and IECM algorithms.The optimum inheritance model was finally selected based on the AIC(Akaike’s Information Criterion)values and a set of fitness tests. The effect values of major gene, polygene and variance were calculated according to the following formulas:
Wherein, σ2prepresents the phenotypic variance of the population, σ2mgrepresents the genetic variance of major genes, σ2represents the distribution variance of components, and h2mg(%) represents the heritability of major gene.
Results and Analysis
Embryo production frequency of the DH population
The distribution of EPFs of the DH population derived from C49S-87/01Y1-1069 was arranged according to the continuous variable data (Fig.1).As shown in Fig.1, the distribution of EPFs was characterized as genetic effect of major genes with two apparent peaks.The mean of the EPFs was 27.19 with standard deviation of 5.67 and chi-square value of 16.25 (P =0.13).
Genetic model analysis
The EPFs of the DH population were analyzed with mixed major gene and polygene inheritance model using P1, P2 and DH populations[21-26]. Total 7 kinds of 46 inheritance models wereused,including one-major-gene model(A),two-major-gene model (B),polygene model (C), one-major-gene and polygene model (D), two-major-gene and polygene model (E),three-majorgene model (F)and three-major-gene and polygene model (G). Their maximum likelihood values and AIC values were shown in Table 1. According to the principle of minimum AIC value and results of fitness test (Table 2),E-1-4 was concluded as the best model for analyzing the inheritance of EPF in this DH population.
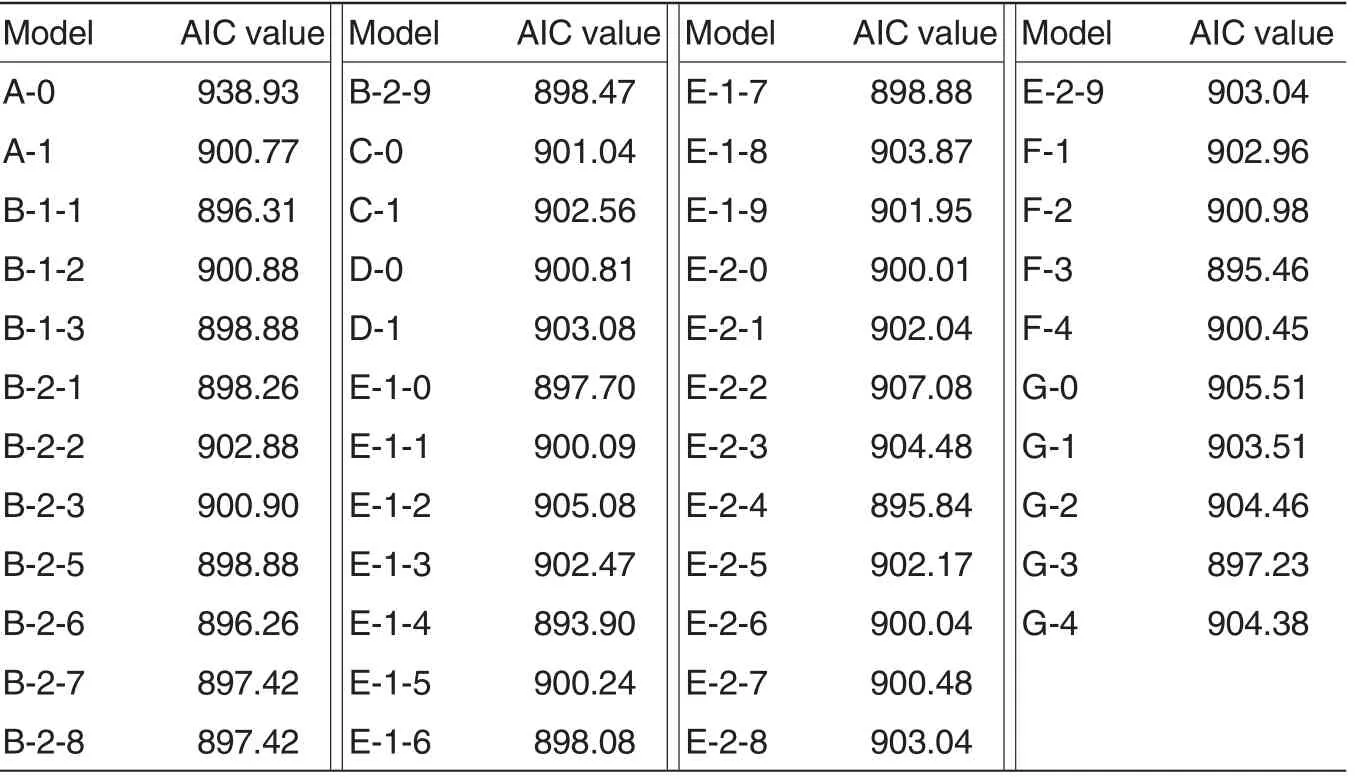
Table 1 AIC values under the 46 inheritance models
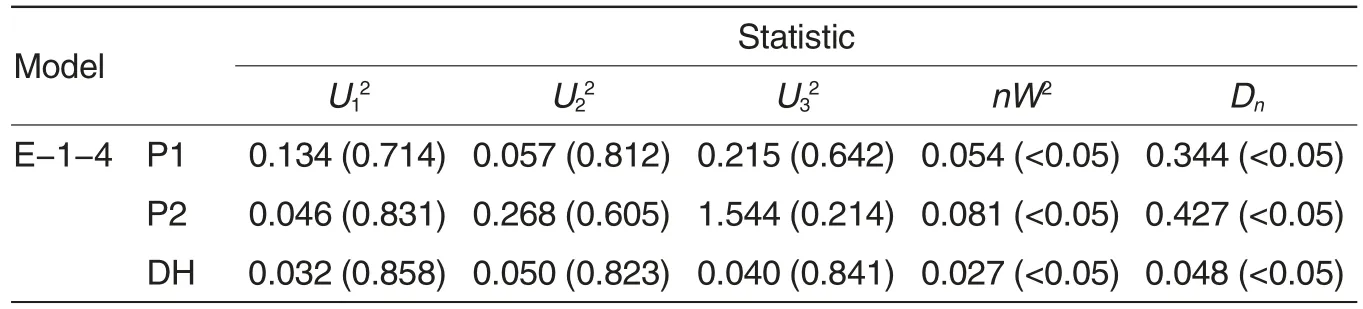
Table 2 Fitness tests of models for the DH population derived from C49S-87/01Y1-1069
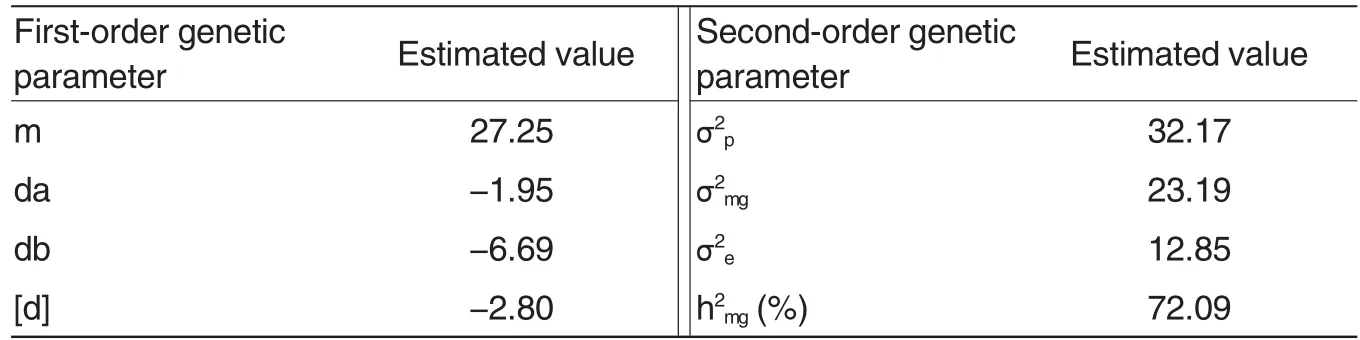
Table 3 Genetic parameters estimation of EPFs of the DH population derived from C49S-87/01Y1-1069
Estimation of genetic parameters
Based on the model analysis and fitness tests for the EPFs of the DH population, it could be concluded that the inheritance of EPFs in the DH population was fitted to the two dominant epistatic major genes plus polygene model (E-1-4). And the maximum likelihood estimates of various genetic parameters were calculated from the component distribution of the optimum model, and then the first-order and second-order genetic parameters were further estimated(Table 3).As shown in Table 3,the inheritance of EPFs in the DH population derived from C49S-87/01Y1-1069 was controlled by two dominant epistasis major genes. The genetic effects of the first and second major genes were 1.95 and 6.69, respectively. The population was composed of three normal distributions(0.503 246 ∶0.252 494 ∶0.244 260 ≈2∶1∶1). The inheritability of major gene was as high as 72.09% , while the polygenic effect was 2.80, suggesting that the EPF of wheat material is mainly impacted by the genotype[25].
Discussion
The expression of quantitative traits is greatly influenced by the environment[26].The adoption of genetically stable population is an effective way to improve the efficiency of detecting the quantitative trait loci and the accuracy of estimating the genetic effect, such as doubled haploid (DH), recombinant inbred lines (RIL) and backcross inbred population(BIL)[22,27].In this study,the DH population and its P1 and P2 were analyzed jointly. The results show that the inheritance of EPF in wheat × maize cross is controlled by two dominant epistatic genes, and the inheritability of the major gene reaches 72.09%. In the previous study on F2population of the same combination,the inheritance of EPF is found to be controlled by two allelic dominant major genes, and the inheritability of the major gene is 43.46%[28]. The results from F2and DH populations all showed that the inheritance of EPF is controlled by two dominant major genes,but there are great differences in the mode of gene action and inheritability of major gene, which might be due to the differences in genetic composition between the two populations. Within the F2population, the genotype is different among different florets in a same plant, so the EPF of each plant is actually the average of different genotypes. However, within the DH population,the genotype is theoretically the same among different florets in a same plant, so the study results are more accurate and reliable. Considering the higher inheritability of major gene in EPF, it is expected that using materials with higher EPFs as cross parents, their offspring will get higher EPFs in practical DH breeding. In this study, although the EPF of female parent C49S-87 is only 10%-15%,the EPF of male parent 01Y1-1069 is higher than 40%, so the average EPF of the 138 DH lines is 27.19%, and about half of the DH lines exceed 30%.In contrast, if the EPFs of double parents are lower,it is needed to increase the pollinated wheat spikes during the crossing between wheat and maize so as to generate enough doubled haploid plants for breeding selection.
In addition, although the EPF of haploid embryos is mainly influenced by genotype, the polygenic effect(2.80) even exceeded the genetic effect of the first major gene (1.95), so the non-genetic factors cannot be ignored. In practice, in order to further stabilize and improve the overall EPF of wheat materials, the protocols for inducing haploid embryos by wheat ×maize system should be improved and standardized, such as improving the quality of emasculation (to avoid injuring stigma), screening high-inductivity maize varieties, adopting high-viability maize pollen for pollination and improving culture conditions (nutrition,temperature,light,etc.)of cut spikes.
[1]ZENKTELER M, NITZSCHE W. Wide hybridization experiments in cereals[J].Theor Appl Genet,1984,68:311-315.
[2]LAURIE DA, BENNETT MD. Wheat ×maize hybridization[J]. Can J Genet Cytol,1986,28:313-316.
[3]LAURIE DA, BENNETT MD. The effect of the crossability loci Kr1 and Kr2 on fertilization frequency in hexaploid wheat × maize crosses [J]. Theor Appl Genet,1987,73:403-409.
[4]INAGAKI M, TAHIR M. Comparison of haploid production frequency in wheat varieties crossed with Hordeum bulbosum L. and maize [J]. Japan J Breed,1990,40:209-216.
[5]SUENAGA K, NAKAJIMA J. Efficient production of haploid wheat (Triticum aestivum) through crosses between Japanese wheat and maize(Zea mays)[J].Plant Cell Rep,1989,8:263-266.
[6]MJUEEB-KAZIA. Polyhaploidy in the Triticeae madited by crosses of Triticum species with Zea mays and Tripsacum datyloides. 8thInt[M]. Wheat Genet Sympo Beijing,1993:1119-1123.
[7]CHEN XM (陈新民), LI GX (赖桂贤),CHEN X (陈孝),et al. Different haploid production in crosses between different wheat F1and maize (不同小麦组合与玉米杂交产生单倍体的差异)[J].Acta Agronomica Sinica (作物学报),1996,22(4):437-441.
[8]LAURIE DA, REYMONDIE S. High frequencies of fertilization and haploid seeding production in crosses between commercial hexaploid wheat varieties and maize[J].Plant Breeding,1991,106:182-189.
[9]KISAN NS, NKONGOLO KK, QUICK JS, et al. Production of haploid wheat plants from wheat × maize crosses[J].Plant Breed,1993,110:96-102.
[10]CHRISTINA B, SABINE G, TAMAS L.Effects of parental genotypes on haploid embryo and plantlet formation in wheat×maize crosses [J].Euphytica,1998,103:319-323.
[11]CAI H(蔡华),MA CX(马传喜),LU WZ(陆维忠). Study on wheat haploid production induced by distant hybridization between different wheat combinations and different-genotype maize (不同小麦组合与不同玉米基因型远缘杂交诱导小麦单倍体的研究)[J]. Crops(作物杂志),2005,3:67-69.
[12]KISAN NS. Production of haploid wheat plants from wheat × maize crosses[J]. Plant Breeding, 1993, 110:96-102.
[13]CHERKAOUT S,LAMSAOURI O,CHLYAH A, et al. Durnm wheat × maize crosses for haploid wheat production:influence of parental genotypes and various experimental factors[J]. Plant Breeding,2000,119:31-36.
[14]VINESH VERMA, BAINS NS, NANDA GS, et al. Maize genotypes show striking differences for induction and regeneration of haploid wheat embryos in wheat × maize system [J].Crop Science,1999,39:1722-1727.
[15]CHEN XM (陈新民), LI XY (李学渊),CHEN X(陈孝),et al.Effect of different crossing techniques on haploid production in wheat×maize crosses(不同杂交技术对小麦×玉米产生单倍体的影响)[J].Acta Agronomica Sinica(作物学报),1998,24(6):743-746.
[16]MARTINS-LOPES PF, GUEDES-PINTO H, PINTO-CARNIDE O, et al. The effect of spikelet position on the success frequencies of wheat haploid production using the maize cross system[J]. Euphytica, 2001, 121: 265 -271.
[17]LAURIE DA. Factors affecting fertilization frequency in crosses of Triticum aestivum cv. ‘Highbury’ × Zea mays cv. ‘Seneca 60’[J]. Plant Breeding,1989,103(2):133-140.
[18]BALLESTEROS J, GARCIA-LLAMAS C, RAMIREZ MC, et al. Low relative humidity increase haploid production in durum wheat × maize crosses[J].Plant Breeding,2003,122:276-278.
[19]GU J(顾坚),LIU K(刘琨),LI SX(李绍祥), et al. Preliminary study on using wheat×maize to induce wheat haploid plant in natural condition of Kunming(昆明自然条件下利用小麦×玉米诱导小麦单倍体的研究初报)[J].Journal of Triticeae Crops(麦类作物学报),2006,26(4):23-26.
[20]WANG JK(王建康),GAI JY (盖钧镒).Identification of major gene and polygene mixed inheritance model and estimation of genetic parameters of a quantitative trait from F2progeny(利用杂种F2世代鉴定数量性状主基因-多基因的混合遗传模型并估计其遗传效应)[J]. Journal of Genetics and Genomics(遗传学报),1997,24(5):432-440.
[21]GAI JY(盖钧镒),ZHANG YM(章元明),WANG JK(王建康).Genetic system of quantitative traits of plants (植物数量性状遗传体系)[M]. Beijing: Science Press(北京: 科学出版社),2003.
[22]ZHANG YM(章元明),GAI JY(盖钧镒),WANG YJ (王永军).An expansion of joint segregation analysis of quantitative trait for using P1, P2 and DH or RIL populations(利用P1、P2 和DH 或RIL 群体联合分离分析的拓展)[J].Hereditas(Beijing)(遗传),2001,23(5):467-470.
[23]GAI JY(盖钧镒).Segregation analysis of genetic system of quantitative traits in plants (植物数量性状遗传体系的分离分析方法研究)[J]. Hereditas (Beijing)(遗传),2005,27(1):130-136.
[24]REN LJ (任丽娟), ZHANG X (张旭),ZHOU MP(周淼平),et al.Heritability of resistance to wheat spindle streak mosaic virus(小麦梭条花叶病抗性遗传分析)[J].Journal of Triticeae Crops(麦类作物学报),2008,28(1):154-159.
[25]LI JQ(李璟琦),MA L(马伶),WEI C(魏翠). Study of dedifferentiation characteristics of immature embryos from twelve kinds of wheat (Triticum aestivum L.) varieties in the middle of Guanzhong irrigated district of Shaanxi Province(陕西关中中部灌区12 个小麦品种幼胚脱分化特性研究)[J]. Journal of Anhui Agricultural Sciences (安徽农业科学),2012,18:9615-9616,9687.
[26]YAN J (严俊), YANG RZ (杨荣志),XUE WT(薛文韬),et al.QTL analysis of 1 000-grain weight under different ecological environment in tetraploid wheat (不同生态环境下四倍体小麦千粒重的QTL 分析)[J]. Guizhou Agricultural Sciences (贵州农业科学),2011,11:6-10.
[27]ZHANG YM(章元明),GAI GY(盖钩镒),QI CK (戚存扣).The precision of segregating analysis of quantitative trait and its improving methods(数量性状分离分析的精确度及其改善途径)[J].Acta Agronomica Sinica (作物学报),2001,27(6):788-793.
[28]LI SX(李绍祥),TIAN YX(田玉仙),GU J(顾坚), et al. Mixed major gene plus polygene inheritance analysis of haploid embryo production frequency in wheat×maize cross (小麦×玉米杂交单倍体胚成胚率的主基因+多基因混合遗传分析)[J].Southwest China Journal of Agricultural Sciences (西南农业学报),2012,25(1):15-18.
猜你喜欢
杂志排行

Agricultural Science & Technology的其它文章
- Evaluation on Suitability of Camellia sinensis Planting Based on GIS
- Light Quality-controlled Phytochemicals Biosynthesis in Vegetables and Fruits
- Cloning and Characterization of Phytochrome A Gene FaPHYA from Tall Fescue
- The Application Effects of Truly Biodegradable Mulch in Potato Farmlands
- The Analysis and Prospect of Development of Fresh Cut Flower Industry Based on the Patent Analysis
- A Comparative Study on Pattern of Urban Forest Patch and Its Ecological Benefit Evaluation in the Park of Hefei Based on the GIS Technology