基于三七连作障碍的土壤理化性状及酶活性研究
2015-12-06孙雪婷龙光强张广辉孟珍贵陈中坚杨生超陈军文
孙雪婷,龙光强,张广辉,孟珍贵,陈中坚,杨生超,陈军文*
1.云南农业大学农学与生物技术学院,云南 昆明 650201;2. 云南农业大学 云南省优势中药材规范化种植工程研究中心,云南 昆明 650201;3. 文山学院三七研究所,云南 文山 663000;
基于三七连作障碍的土壤理化性状及酶活性研究
孙雪婷1,2,龙光强2,张广辉2,孟珍贵2,陈中坚3,杨生超2,陈军文1,2*
1.云南农业大学农学与生物技术学院,云南 昆明 650201;2. 云南农业大学 云南省优势中药材规范化种植工程研究中心,云南 昆明 650201;3. 文山学院三七研究所,云南 文山 663000;
为评价三七(Panax notoginseng (Burk) F. H. Chen)连作对土壤理化性状及土壤酶活性的影响,分析了三七不同种植年限及连作土不同治理方法的土壤理化性状和酶活性,从而为部分解析三七连作障碍形成机制及为缓解并克服连作障碍提供理论依据。结果表明,随着三七种植年限的增加,土壤pH呈显著下降趋势,土壤酸化现象加重;种植三七1年、2年和3年的土壤与未种植过三七的土壤相比有机质含量分别下降73%、87%和83%,并伴随着土壤的板结。与生土和健康土相比,大量元素氮(N)和磷(P)含量在种植三七的土壤及发病土壤中显著下降。铁(Fe)、硼(B)、铝(Al)等元素在种植三七的土壤中却显著升高,种植三七3年的土壤与生土相比,Fe、B、Al分别上升29%、31%、32%。土壤蛋白酶、过氧化氢酶、蔗糖酶、磷酸酶及脲酶活性随着三七种植年限的增加均显著降低,种植三七3年的土壤中,其土壤蛋白酶、过氧化氢酶、蔗糖酶、脲酶及磷酸酶较未种植过三七的土壤分别下降91%、56%、52%、92%、92%和69%;而发病植株根际土壤的过氧化氢酶、蔗糖酶、磷酸酶活性高于健康植株根际土壤的酶活性,其中发病土壤的蔗糖酶活性较健康土高出 42%;连作植株根际土壤与经消毒治理后的植株根际土壤相比,脲酶、蛋白酶、多酚氧化酶及过氧化氢酶活性显著较高。研究结果还显示,土壤酶活性与土壤pH、有机质含量、N、P、K及一些微量元素(如:钼、铜)之间存在显著相关关系。本研究结果暗示,三七连作后导致了土壤酸化和板结现象加重及土壤矿质元素组成失衡和某些元素的富集,这可能是诱导三七连作障碍发生的原因之一;另外,三七种植年限增加明显抑制土壤酶活性,这将阻碍三七对某些营养元素的吸收及间接加剧土壤自毒物质的积累,从而也可能诱导连作障碍的发生。
三七;连作障碍;土壤酸化;矿质元素;土壤酶
连作障碍(Continuous cropping obstacles)是指在同一块土地中连续栽培同种或同科作物时,即使在正常的栽培管理条件下,也会出现生长势变弱、产量降低、品质下降、病虫害严重的现象(张重义和林文雄,2009)。前人通过对粮食和牧草作物(Kong等,2004;Meriles等,2009;Chona等,2002),如水稻(Oryza.sativa L)、大豆(Giycine max(L)Merrill)、苜蓿(Medicago sativa Linn),园艺作物(Wu等,2009;Hao等,2007;Mondal等,2013),如黄瓜(Cucumis sativus Linn)、西瓜(Citrullus lanatus(Thunb)Mansfeld)、草莓(Fragaria ananassa Duchesne)的研究发现,导致作物发生连作障碍的主要因素有(寻路路, 2013):(1)根际正常微生物群落结构发生变化,病原微生物增加,有益微生物减少;(2)作物分泌的有机物对其自身产生毒害,即化感自毒作用;(3)土壤理化性状的恶变,导致土壤肥力下降及土壤盐渍化和酸化。目前传统大田作物的连作障碍研究受到了广泛关注,然而对药用作物连作障碍的研究则相对较少。
近年来,大多研究主要集中从土壤微生物群落的变化(Ling等,2013;Cao等,2011;Haggag等,2008;Lang等,2011)及土壤中自毒物质的提取和鉴定(He等,2009;Canals等,2005;Jefferson和 Pennacchio,2003;Liu等,2008)等方面解析连作障碍的成因,而土壤理化性状变化对连作障碍影响的研究则相对受到忽视。研究发现,作物连作后土壤粘性增大,土壤交换性盐基总量及盐基饱和度降低,土壤通气空隙降低,土壤容重增大,土壤酸化及盐渍化现象加重(Lithourgidis等,2006)。同时,作物连作后土壤养分失衡,有机质含量下降,磷(P)含量过剩,而钾(K)含量不足(Lithourgidis等,2006;France,1989)。另一方面,土壤酶作为土壤体系中一切生化反应的“催化剂”,对土壤养分转化及土壤生物活性起监测作用(Asmer等,1992;Dick等,1998;Frankenberger和Dick,1983)。棉花(Gossypium hirsutum Linn.)连作后棉田土壤脲酶的活性随着连作年限的增加,呈先升后降趋势,过氧化氢酶活性呈先降后升趋势(张伟等,2011)。虽然有研究证实,连作会导致土壤酶活性发生变化,但土壤酶在作物产生连作障碍过程中扮演的角色依然不是很清楚。
三七为五加科人参属多年生草本植物,以根及根状茎入药,具化瘀止血、消肿定痛等功效,是我国名贵中药材(国家药典委员会,2005)。三七为典型的宿根植物,忌地性极强。据前期调查,种植过三七的农田需要轮作玉米等作物至少 10年以上才可再种植三七。有研究表明,种植三七导致土壤蔗糖酶、淀粉酶、过氧化氢酶、多酚氧化酶和磷酸酶的活性降低,也导致土壤有机质、有效K、有效Ni、有效Cu、有效Zn、有效Fe含量下降(简在友等,2009)。但相关的研究还缺乏对三七连作后植株根际土壤、健康植株根际土壤、发病植株根际土壤及经消毒治理的植株根际土壤的理化性状及酶活性的系统评估。本文通过在三七道地产区文山州采集三种类型的土样:(1)种植三七1年、2年、3年的根际土样及未种植三七的生土;(2)同一地块种植三七两年后未发生严重病害的畦面的健康土与发生严重病害的畦面的发病土;(3)不能重茬的连作土和相应进行过消毒处理的治理土;然后对比分析这些系列土样的理化性状和土壤酶活性及其两者的相关性,以期为部分解析三七连作障碍形成机制提供基础数据。
1 材料与方法
1.1 试验地概况
试验所用土样均采自于三七传统道地产区云南省文山州砚山县。砚山县位于云南省东南部,东经 103°35~104°45、北纬 23°18~23°59之间,海拔1539 m,属低纬北亚热带高原季风气候,年平均气温16.1 ℃,正常年降雨量1008 mm,全年无霜期250~320 d,是三七的主产区之一。
1.2 土样的采集
于2013年12月,在云南省文山州砚山县的文山苗乡三七科技园及其附近区域选点取土样。样品采集地条件:年平均气温 17.5 ℃,最高气温不高于33 ℃;降水量(年)1080 mm;海拔:1557 m;年平均湿度:75%~85%;土壤类型:红壤。
选择未种植过三七的地块及种植三七 1年、2年、3年的地块,取自未种植过三七地块的土壤视为生土,种植三七的地块取三七根际土壤。种植三七的地块与未种植三七的地块相距约50 m。据走访调查,两者整理为农用地之前都为多年的弃荒地,且之前都未种植过三七。
三七播种之前,结合深耕,将钙镁磷肥均匀施入土壤,使其与土层混合。三七种植过程中,于 4月中旬左右,按8.1 g·m-2施磷酸二铵((NH4)2HPO4)和普钙,6月中旬左右按 10.8 g·m-2施硝铵(NH4NO3)、硫酸钾、按6.8 g·m-2施磷酸二铵、按13.5 g·m-2施复合肥。3月份喷施杀虫药(多抗霉素、链霉素等),4月针对疫病喷施噁霉灵等;后期(9月以后)针对病情在需要的时候选择不同的农药进行防治病虫害。晴天检查水分状况,并进行补水;雨天根据植株发病情况,对有病的种苗及时清除,防止病情扩散,并实时监管,使三七免受病、虫、草害。
在本研究中,健康土指的是种植三七两年但并未发生严重病害且符合三七生产要求的地块的土壤,发病土指的是来自同一地块的、但发病特别厉害、几乎无健康植株存在的不同畦面的土壤。健康土和发病土都取自三七根际土壤。
连作土为来自三七连作两年以上、在生产上被认为是在相当长的时间内不能再种植三七的地块,治理土为上述连作土经过人为治理消毒(主要对土壤病害微生物进行消毒)后再种植三七的地块的土样。其中连作土1和连作土2为来自三七不同种植区域的土壤,相应的治理土1和治理土2为采用不同方法消毒治理的土样。其中,治理土1为三七采收后(12月),将氯化苦(CCl3NO2)配成一定浓度,用抽提剂注射到土壤中,薄膜覆盖,每隔3个月做一次重复处理,连续处理3次,治理土2为相同处理不同试验地采集的土地,治理土1和治理土2采集地相距50 m左右。前期试验结果表明,在连作土 1和连作土 2的两种连作土上种植三七一年后,存苗率都不足10%;而在两种治理土上种植三七一年后,存苗率在70%以上。在一年试验周期结束后,取连作土和治理土上种植三七的根际土壤。
除未种植三七的弃荒地所采集的生土外,所有待测土样种植管理措施、施肥量、施肥种类均相同。以上土样均采用交叉五点法进行采集,取三七植株根际0~30 cm土壤。取回的三七植株根际土壤,带回室内,风干,除杂,过1~2 mm筛,测定土壤理化性状及土壤酶活性指标。
1.3 测定项目与方法
1.3.1 土壤理化性状的测定
土壤理化性状指标的测定与分析参照《土壤农化分析》和《土壤理化分析》提供的方法(南京农业大学主编,1986;中国科学院南京土壤研究所主编,1997)。土壤pH采用土∶水=1∶5酸度计法,土壤有机质采用 K2Cr2O7-H2SO4消煮、FeSO4容量法测定;土壤全氮用重铬酸钾、硫酸消化-蒸馏法,有效氮用碱解扩散法测定;土壤全磷用碱熔-钼锑抗比色法测定,有效磷用Olsen法测定;土壤全钾及有效钾分别用火焰光度法和中性 NH4Ac浸提法测定;土壤全铝及有效铝均用氟化钾取代-EDTA滴定法测定;全硫测定用燃烧碘量法,有效硫测定用硫酸钡比浊法;土壤中全硼用碳酸钠熔融-甲亚胺法测定,有效硼用热水回流浸提法测定;土壤中全量铜、锌的测定用原子吸收分光光度计测定,有效铜、锌用DTPA-TEA浸提-AAS法测定;全锰含量用原子吸收分光光度法测定,有效锰用 KMnO4比色法测定;土壤中全钼含量用HClO4-HNO3消煮-催化极谱法测定,有效钼用草酸-草酸铵浸提,催化极谱法测定;土壤中全量钙、镁的测定用EDTA滴定法,有效钙、镁的测定用原子吸收分光光度法测定;土壤中全铁测定用原子吸收光谱法,有效铁的测定用DTPA溶液浸提-原子吸收光谱法测定;土壤钠的测定用火焰光度法;生土及种植土的土壤理化性状分析由中国科学院西双版纳热带植物园生物地球化学实验室完成,其余土壤样品的理化性状分析由云南省农业科学院环境资源研究所完成。
1.3.2 土壤酶活性的测定
土壤酶活性测定方法参照关松荫编著的《土壤酶及其研究法》(关松荫,1986)。
脲酶采用苯酚钠-次氯酸钠比色法测定,以24 h后1 g土壤中NH3-N的毫克数表示酶活性。土壤蔗糖酶活性采用3,5-二硝基水杨酸比色法测定,蔗糖酶的活性以24 h后1 g土壤生成葡萄糖的毫克数表示。用邻苯三酚比色法测定土壤多酚氧化酶的活性,多酚氧化酶活性以2 h后1 g土壤中生成的紫色没食子素的毫克数表示。用滴定法测定过氧化氢酶的活性,其活性以每克土所消耗的 0.1 mol·L-1KMnO4的毫升数表示。蛋白酶采用铜盐比色法测定,土壤蛋白酶活性以每克土壤在24 h内酶解蛋白质释放的NH3-N的毫克数(mg)表示;磷酸酶采用磷酸苯二钠比色法测定,酶活性以24 h后1 g土壤中释放出的中P2O5毫克数表示。
1.4 数据统计分析
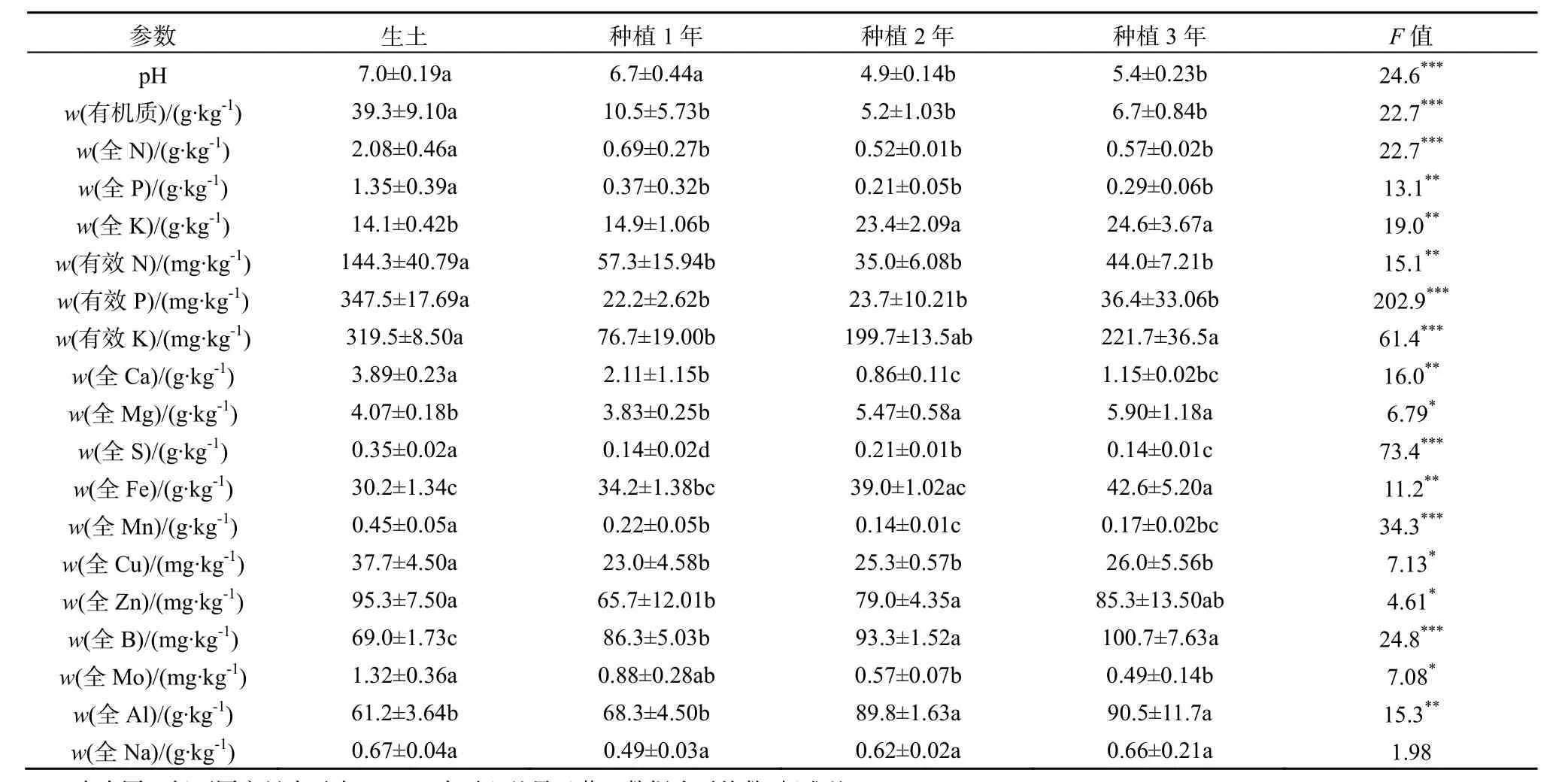
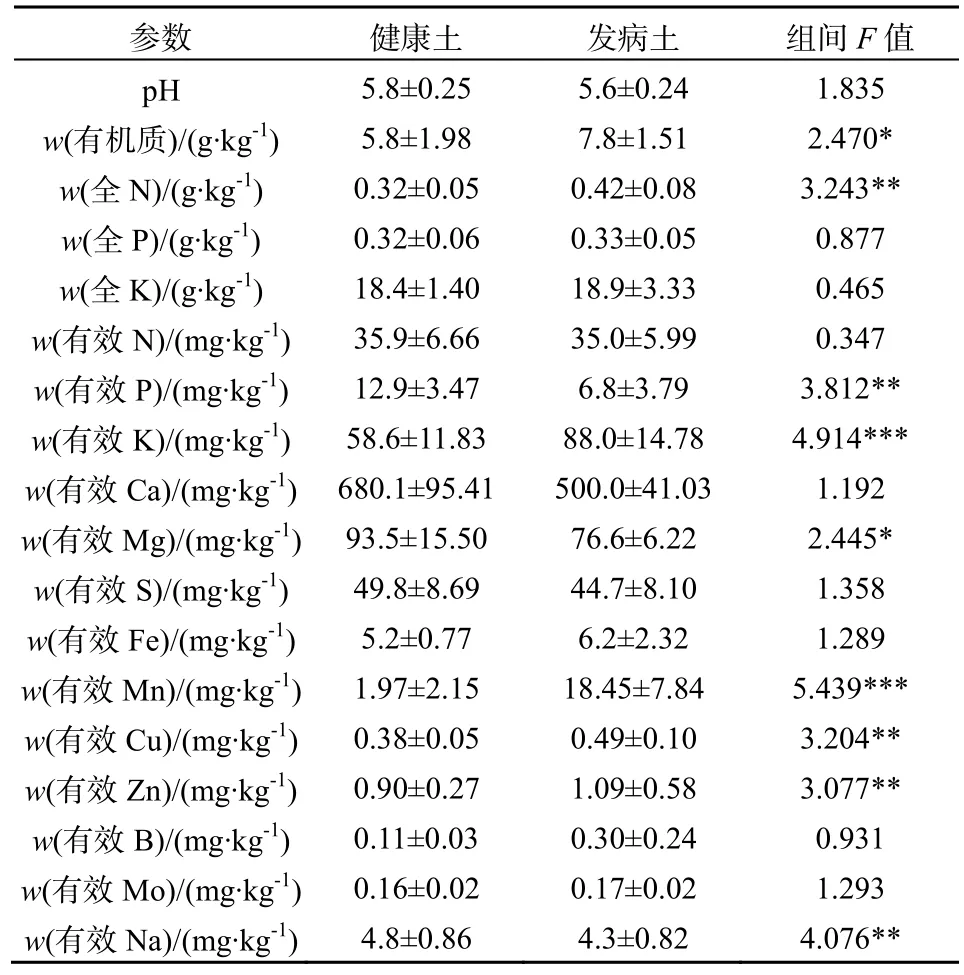
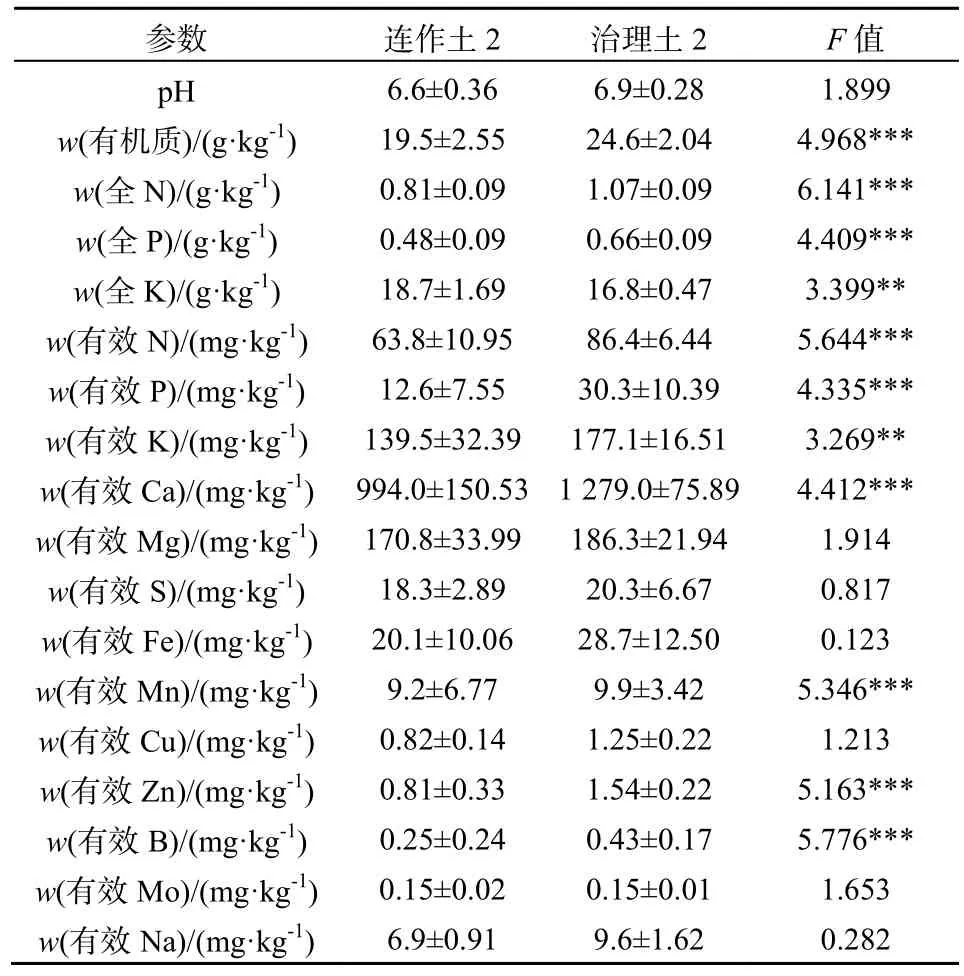
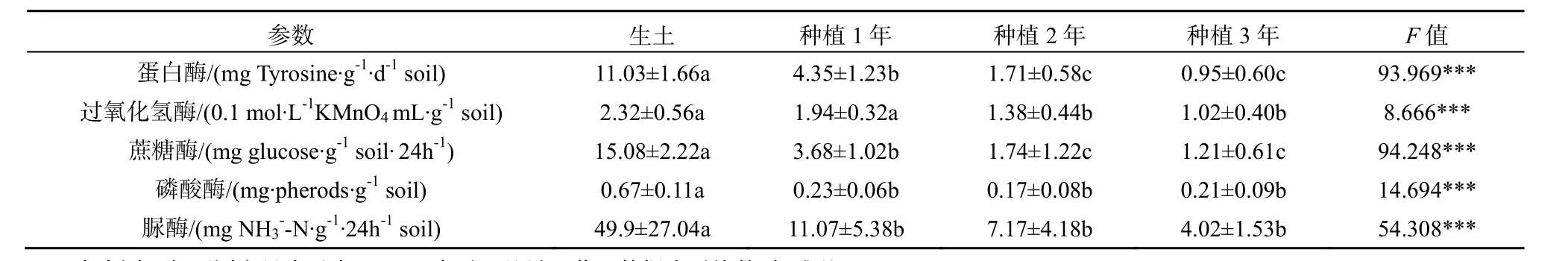
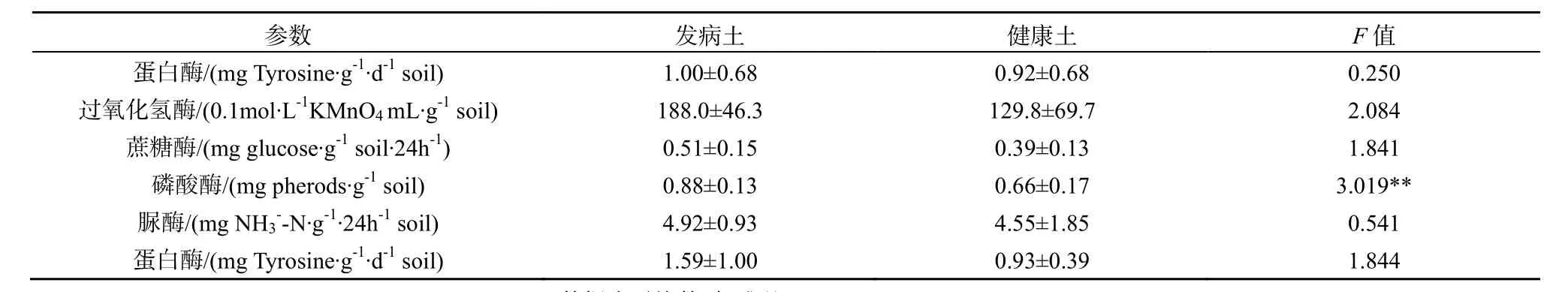
所有数据均使用Microsoft Excel 和SPSS 17.0软件进行整理和分析。其中种植三七1、2、3年地块的土样及生土,其土壤理化性状及酶活性实验结果用单因素方差分析(one way ANOVA)检验组间差异;其余土样的理化性状及酶活性数据采用独立样本t检验(Independent Sample T test)分析其显著性差异;用Pearson相关系数分析土壤酶活性与土壤理化性状间的相关关系。取自一个试验小区的土壤为一个重复。*0.01 2.1 连作对三七种植土壤理化性状的影响 2.1.1 三七不同种植年限土壤的理化性状比较 三七不同种植年限对土壤理化性状有明显影响(表1)。种植三七3年后的土壤较未种植过三七的土壤pH呈下降趋势;且总体上看,种植三七后土壤pH呈下降趋势。种植三七1年、2年、3年后土壤有机质含量比未种植过三七的生土有机质含量分别下降73%、87%和83%。 种植三七3年后土壤中全N、全P、有效N、有效P较生土分别下降73%、79%、70%和90%,且差异显著;而随着种植年限的增加,有效K含量总体呈现上升趋势,种植三七3年后土壤全K含量增加43%。中量元素全Ca、全S的含量随种植年限增加呈下降趋势,而全Mg含量呈现显著上升趋势。相比生土,种植三七3年后土壤中全量Fe、B、Al含量分别上升29%、31%、32%;而微量元素全量Mn、Cu、Zn、Mo的含量呈下降趋势;随着种植年限的增加,全Na的含量虽然呈现小幅度下降,但变化不显著。 2.1.2 三七根际健康土和发病土理化性状比较 由表2可知,健康土与发病土的pH并无明显差异。发病土有机质含量显著高于健康土。发病土有效K比健康土中高33%,而有效P含量却比健康土低48%。全N含量在发病土中显著较高,而有效N、全P、全K两者差异不显著。发病土中有效Ca、有效Mg、有效S、有效Na的含量均低于健康土。而有效Fe、有效Mn、有效Cu、有效Zn、有效B、有效Mo的含量均是发病土中较高。 2.1.3 三七连作土与治理土理化性状差异 如表3所示,连作土1的pH显著高于治理土1;连作土1中大量元素P、K及中量元素有效Ca、有效Mg含量均比治理土1高;连作土1中的有效Cu、有效Mo含量较治理土1高,并达到极显著水平;连作土1中大量元素全N含量低于治理土1,但差异不显著;中量元素S含量显著低于治理土1;微量元素有效Fe、Mn含量均较治理土1低,达到极显著水平;连作土1中有效Na含量显著低于治理土1。 表1 不同种植年限三七根际土壤的理化性状Table 1 Physical-chemical properties of rhizosphere soil of Panax notoginseng cultivated by different years 表中同一行不同字母表示在P≤0.05水平上差异显著,数据为平均数±标准差(n=3) 表2 三七种植园健康土和发病土的土壤理化性状Table 2 Physical-chemical properties of rhizosphere soil collected from healthy and diseased Panax notoginseng 表3 三七连作土1与治理土1土壤理化性状Table 3 Physical-chemical properties of soil collected from continuous cropping system 1 and correspondingly treated system 1 for Panax notoginseng 由表4可知,治理土2中有机质含量显著高于连作土2;治理土2中大量元素N、P及有效K含量均显著高于连作土 2,达到极显著水平(P<0.001);治理土2中的中量元素有效Ca含量高出连作土 222%,达到极显著水平(P<0.001);治理土2中微量元素有效Mn、有效Zn、有效B含量高于连作土 2,达到极显著水平(P<0.001);治理土2中全K含量比连作土2低。而pH在治理土2和连作土2中的差异不显著,两者呈微酸性。 表4 三七连作土2与治理土2土壤理化性状Table 4 Physical-chemical properties of soil collected from continuous cropping system 2 and correspondingly treated system 2 for Panax notoginseng 2.2 三七连作对土壤酶活性的影响 2.2.1 三七不同种植年限土壤酶活性比较 随着三七种植年限的增加,土壤蛋白酶、过氧化氢酶、蔗糖酶、磷酸酶和脲酶的活性均呈显著下降趋势(表5)。种植三七1年、2年、3年后土壤蛋白酶活性较未种植过三七的生土其活性分别下降61%、84%和91%;在种植三七3年后,过氧化氢酶活性下降了56%,蔗糖酶和脲酶活性均下降了92%,磷酸酶下降了69%。 表5 不同种植年限三七根际土壤酶活性Table 5 Enzyme activities of soil collected from rhizosphere of Panax notoginseng cultivated by different years 2.2.2 三七根际健康土和发病土的土壤酶活性比较 三七种植园健康土和发病土的土壤酶活性分析发现,发病土中各测定的酶活性均高于健康土(表 6)。其中发病土中磷酸酶活性较健康土高出25%,与健康土相比显著较高;发病土的多酚氧化酶的活性高于健康土30%;发病土中过氧化氢酶及蔗糖酶活性较健康土分别高出24%和42%;发病土的蛋白酶活性和脲酶活性均高于健康土,但差异不显著。 表6 三七种植园健康土和发病土酶活性比较Table 6 Enzyme activities of soil collected from rhizosphere of healthy and diseased Panax notoginseng 2.2.3 三七连作土与治理土土壤酶活性比较 由表7可知,连作土1中蛋白酶、多酚氧化酶、脲酶、及蔗糖酶活性均高于治理土 1,过氧化氢酶和磷酸酶活性却低于治理土1。其中连作土1蛋白酶活性高于治理土 1达 38%;多酚氧化酶活性高15%,达到显著水平;脲酶活性高达49%,达到极显著水平;蔗糖酶活性高达32%;治理土1中磷酸酶活性高于连作土32%,达到极显著水平。 连作土2中多酚氧化酶、过氧化氢酶及磷酸酶活性均高于治理土 2,蛋白酶活性、脲酶活性及蔗糖酶活性均低于治理土2(表8)。连作土2中多酚氧化酶活性高于治理土2的34%,达到显著水平;过氧化氢酶活性在连作土2中较高,但差异不显著;连作土2中磷酸酶活性较治理土2高48%,达到极显著水平;治理土 2中蛋白酶活性较连作土 2高51%;连作土2中脲酶及蔗糖酶活性均显著高于治理土2,其活性分别是治理土2的47%和40%。 2.3 土壤酶活性与土壤理化性状的相关性 由表9可知,所测土壤酶活性与土壤理化具有一定的相关性。其中pH、有机质、全P、有效P与蛋白酶活性呈显著正相关,S、Mn与蛋白酶活性呈负相关;过氧化氢酶活性与Mo呈显著正相关,与Mn呈极显著负相关,与Ca、Mg、S呈负相关,与全K及Na也呈现负相关关系;磷酸酶活性与Na、Ca、Mg呈显著正相关关系,而与B、Mo、Cu、Zn、Fe呈负相关关系;蔗糖酶活性及脲酶活性与有机质、全N、全P、有效N、有效P以及Mo、Cu含量呈现显著正相关关系,而Na、S、Ca、Mg、Mn与其呈负相关关系。 表7 三七连作土1与治理土1酶活性比较Table 7 Enzyme activities of soil collected from continuous cropping system 1 and correspondingly treated system 1 for Panax notoginseng 表8 三七连作土2与治理土2酶活性比较Table 8 Enzyme activities of soil collected from continuous cropping system 2 and correspondingly treated system 2 for Panax notoginseng 表9 土壤酶活性与土壤理化性状的相关性Table 9 Correlations between activities of soil enzyme and properties of soil physic-chemistry 3.1 三七连作驱动的土壤理化性状变化 目前普遍认为,土壤理化性状恶变是造成作物连作障碍发生的主要因素之一。有研究表明,随着三七连作年限的增加,土壤有效K及有效S含量呈下降趋势,而P、Cu、Mg等元素却逐年升高,土壤酸化及盐渍化水平加重(刘莉等,2013)。本研究结果也表明,随着三七种植年限的增加,土壤pH呈显著下降趋势。土壤pH下降说明土壤有酸化的趋势,土壤酸化加重将为病原真菌滋长提供条件,从而可能加重病虫害的发生(Moharana等,2012)。种植三七后,土壤有机质含量和大量元素氮(N)、磷(P)含量显著下降,而全Fe、全B、全Al等却显著升高,K含量却随种植年限的增加而呈上升趋势(表1)。三七是一种喜钾作物,K含量增加可能与生产上人为追求产量施加硫酸钾过量有关。K+能与土壤胶体吸附的Ca2+、Mg2+等阳离子发生交换,使土壤板结,导致土壤中有机质含量下降,并影响植物呼吸(MOHARANA,2012)。土壤中微生物活动与有机质含量密切相关,K含量增高而有机质含量降低,可能会使土壤中微生物群落的多样性遭到破坏,同时导致土壤板结、透气性降低、肥力下降,这可能会加剧作物连作障碍发生(Wei等,2006)。发病土中有效Fe、有效Mn、有效Cu、有效Zn等元素含量比健康土高,连作土2也表现为有效Fe、有效Cu等元素含量较高。有研究表明,土壤中Fe、Al含量过高,会抑制植物质膜ATP的活性,阻碍根系生长,影响植物对营养物质的吸收和利用,对植株造成伤害,使作物产量及质量下降(Nicola等,2003)。 土壤理化性状变化是植物内在因素及人为管理措施等外在因素共同作用的结果。植物在生长过程中对某些营养元素的选择性吸收会导致这些元素的缺乏,而另外一些元素却发生富集,这将致使土壤营养物质的平衡遭到破坏。有研究表明,由于人参(Panax ginseng C. A. Mey.)对土壤矿质元素的吸收偏好,人参重茬三年土与生茬土相比 Fe、Mn、Zn、Cu、K、B的含量明显降低,而Na、Ca含量却恰恰相反(Diekow等,2005)。同时也有研究证实,在连作小麦(Triticum aestivum Linn.)和玉米(Zea mays)的土壤中,添施肥料(如磷肥)后,土壤中Fe、Mn、Zn的含量显著增加,Cu含量未受到影响(Gao等,2000)。Na、Al、Mn、Fe及Cu、Zn等矿质元素高浓度时对植物有毒害作用。在三七生产过程中,多用遮荫棚进行设施栽培,土壤中的无机元素容易富集于土壤表层,再加之化学肥料的过度施用,更容易导致土壤表层某些元素的集聚或缺素,这些因素都会对植株生长产生不良影响。同时,营养元素富集还会滋长病原真菌积累(Asmer等,1992),导致土壤病害加重,进一步加剧连作障碍的发生。 3.2 三七连作对土壤酶活性的影响 土壤酶主要来自于植物根系分泌物及土壤微生物的活动,并参与土壤各种生物化学过程,是土壤环境质量监测的敏感指标(苏洁琼等,2014;赵玉涛等,2008;Bandick和Dick,1999)。寻路路(寻路路,2013)研究发现,栽种过三七的土壤蔗糖酶、过氧化氢酶、多酚氧化酶及磷酸酶活性均小于未种植三七的土壤。本研究结果也显示,随着三七种植年限的增加,蛋白酶、过氧化氢酶、蔗糖酶、磷酸酶及脲酶活性均显著降低。而发病土的过氧化氢酶、蔗糖酶、磷酸酶活性高于健康土(表6)。土壤酶在遭遇土壤环境变化的情况下,会产生一些抗逆机制来维持自身稳定性,在初期表现为活性下降,而随着环境干扰作用加强,其生物适应性也会随之加强,出现升高现象(Liu等,2002)。 蔗糖酶作为促进蔗糖分解的重要酶,能够提高土壤的生物活性,是表征土壤氮素循环及肥力水平的重要指标(刘恩科等,2008)。连作土 1的蔗糖酶活性高于治理土1,而连作土2的蔗糖酶活性却显著低于治理土 2(表 7、8),这可能是两种连作土样治理方法不同所致。同时,微生物的代谢产物(酚酸及蔗糖类碳水化合物)也会导致蔗糖酶活性的变化。脲酶活性与蛋白酶活性在连作土1中显著高于治理土 1,发病土与健康土中两种酶活性差异不显著,治理土2中脲酶及蛋白酶活性显著升高(表7、8)。脲酶和蛋白酶都是参与土壤氮素循环的重要酶,对土壤中氮素的含量变化表现敏感(苏洁琼等,2014)。连作土1中全氮(N)含量是生土中的全氮(N)含量的近四倍(表1、表3)。三七连作过程中,为提高产量会加施氮肥,导致三七根际土壤中氮素含量升高,氮素循环加快,这可能是导致连作土1中两种酶活性升高的原因之一。多酚氧化酶是植物根际分泌物、植物腐解及土壤微生物代谢等分解释放的酶,能促使邻苯三酚氧化成醌,使其与土壤中大分子物质如蛋白质、糖类等反应生成有机质及色素,对土壤环境变化极其敏感(王延平等, 2013)。本研究结果表明,多酚氧化酶活性在连作土1与连作土2中基本一致,经治理后其活性降低,原因可能是植物残茬降解及根际分泌的酚酸类物质在土壤中积聚,诱发多酚氧化酶活性上升;而治理后,土壤中微生物被大量消除,土壤分解代谢能力降低,多酚氧化酶活性降低(王延平等,2013;Tian等,2013)。过氧化氢酶作为植物抗氧化系统中的关键酶,它对植物抵御逆境胁迫起到至关重要的作用(廖海兵等,2011)。而发病土及连作土 2中过氧化氢酶活性较低,这与廖海兵等(2011)对连作条件下浙贝母土壤酶活性的研究结果相关类似。这说明三七连作后土壤的抗逆性减弱,这有可能诱发病害的发生。 苏洁琼等人的研究发现,土壤酶(如:蛋白酶、脲酶、磷酸酶、蔗糖酶、过氧化氢酶、多酚氧化酶)活性与土壤全N、NO3-、NH4+-N及有效N均呈极显著负相关,与土壤pH呈正相关关系(王延平等,2013)。本研究结果表明,所测的土壤酶(蛋白酶、脲酶、磷酸酶、蔗糖酶、过氧化氢酶、多酚氧化酶)活性与土壤有机质、全N、全P、有效N、有效P、有效K呈正相关,与全K呈负相关关系。土壤酶通常来自于土壤中微生物的代谢活动、植物根系分泌和动植物残体腐解,所以土壤酶活性对土壤理化性状变化响应灵敏。土壤pH变化、有机物污染等都会引起土壤酶活性发生改变(杨红飞等,2007)。 土壤酶对土壤环境的自我修护及调节起到重要作用,酶活性紊乱会导致植物根际和根系的微域环境发生改变(寻路路,2013)。蔗糖酶活性降低会导致土壤易溶性营养物质减少,阻碍植物对土壤中营养物质的摄取(Liu等,2002)。蛋白酶、磷酸酶和脲酶与土壤氮素及磷素水平息息相关,作物连作导致的蛋白酶及磷酸酶活性降低,会很大程度削弱土壤养分供应能力(刘恩科,2008)。同时,土壤酶活性变化会影响土壤解毒能力,连作导致的土壤酶活性失调,这可能将间接加剧土壤中自毒物质的积累,抑制植物生长,诱导连作障碍的发生。 本研究结果表明,随着三七种植年限的增加,土壤pH呈显著下降趋势,土壤有酸化趋势,土壤酸化会促进病原真菌的生长,可能诱发三七连作障碍;土壤有机质含量和N、P含量在种植三七的土壤及发病土中显著下降,而Fe、B、Al等元素却显著升高,因此,种植三七后土壤有板结的趋势,并且某些元素富集会导致植物中毒及土壤养分比例失调;同时,随着种植年限的增加,土壤蛋白酶、过氧化氢酶、蔗糖酶、磷酸酶和脲酶的活性均呈显著下降趋势,三七连作后土壤中关键酶活性的下降,将影响三七对某些营养元素的吸收,并削弱土壤对自毒物质的代谢,可能诱导连作障碍发生。 ASMER E, EILAND E, LEISEN N E. 1992. Interrelationship between extracellular enzyme activities, ATP content, total counts of bacteria and CO2 evolution[J]. Biology and Fertility of Soils, 14(4): 288-292. BANDICK A K, DICK R P. 1999. Field management effects on soil enzyme Activities[J]. Soil Biology and Biochemistry, 31: 1471-1479. CANALS R M, EMETERIO L S, PERALTA J. 2005. Autotoxicity in Lolium rigidum: analyzing the role of chemically mediated interactions in annual plant populations[J]. Journal of Theoretical Biology, 235(3): 402-407. CAO Y, ZHANG Z, LING N, et al. 2011. Bacillus subtilis SQR 9 cancontrol Fusarium wilt in cucumber by colonizing plant roots[J]. Biology and Fertility of Soils, 47(5): 495-506. CHON S UK, CHOI S K, JUNG S, et al. 2002. Effects of alfalfa leaf extracts and phenolic allelochemicals on early seedling growth and root morphology of alfalfa and barnyard grass[J]. Crop Protection, 21(10): 1077-1082. DICK R E, MYROLD D D, KERLE E A. 1998. Microbial biomass and soil enzyme activities in compacted and rehabilitated skid trail soils[J]. Soil Science Society of America Journal, 2: 512-516. DIEKOW J J, MIELNICZUK J, KNICKER H, et al. 2005. Soil C and N stocks as affected by cropping systems and nitrogen fertilisation in a southern Brazil Acrisol managed under no-tillage for 17 years[J]. Soil and Tillage Research, 81(1): 87-95. FRANCE E A. 1989. Soil chemistry changes after 27 years under four tree species in southern Ontario Canadian[J]. Journal of Forest Research, 19(12): 1648-1650. FRANKENBERGER W T, DICK W A. 1983. Relationships between enzyme activities and microbial growth and activity Indices in soil[J]. Soil Science Society of America Journal, 47(5): 945-951. GAO M, CHE F C, WEI C F, et al. 2000. Effect of long-term application of manures on forms of Fe, Mn, Cu and Zn in purple paddy soil[J]. Journal of Plant Nutrition and Soil Science, 6: 11-17. HAGGAG W M, TIMMUSK S. 2008. Colonization of peanut roots by biofilm-forming Paenibacillus polymyxa initiates biocontrol against crown rot disease[J]. Applied and Environmental Microbiology, 104(4): 961-969. HAO Z P, WANG Q, CHRISTIE P, et al. 2007. Allelopathic potential of watermelon tissues and root exudates[J]. Scientia Horticulturae, 112(3): 315-320. HE C N, GAO W W, YANG J X. 2009. Identification of autotoxic compounds from fibrous roots of Panax quinquefolium L.[J]. Plant and Soil, 318(1-2): 63-72. JEFFERSON L V, PENNACCHIO M. 2003. Allelopathic effects of foliage extracts from four Chenopodiaceae species on seed germination[J]. Journal of Arid Environments, 55(2): 275-285. KONG C H, XU X H, ZHOU B, et al. 2004. Two compounds from allelopathic rice accession and their inhibitory activity on weeds and fungal pathogens[J]. Phytochemistry, 65(8): 1123-1128. LANG J J, HU J, RAN W, et al. 2011. Control of cotton Verticillium wilt andfungal diversity of rhizosphere soils by bio-organic fertilizer[J]. Biology and Fertility of Soils, 48(2): 191-203. LING N, DENG KY, SONG Y, et al. 2014. Variation of rhizosphere bacterial community in watermeloncontinuous mono-cropping soil by long-term application of a novel bioorganic fertilizer[J]. Microbiological Research, 169(7-8): 570-578. LITHOURGIDIS A S, DAMALAS C A, GAGIANAS A A. 2006. Long-term yield patterns for continuous winter wheat cropping in northern Greece[J]. European Journal of Agronomy, 25: 208-214. LIU J G, LI Y B, JIANG G Y, et al. 2008. Allelopathic effects of cotton in continuous cropping[J]. Allelopathy Journal, 21(2): 299-306. LIU X J, LIAO X Y, ZHANG Y Z, et al. 2002. Effects of rice-based cropping system on the distribution of manganese in the profile of paddy soil derived from red earth[J]. Acta ecologica sinica, 22(9): 1440-1445. MERILES J M, VARGAS G B, CONFORTO C, et al. 2009. Soil microbial communities under different soybean cropping systems: Characterization of microbial population dynamics, soil microbial activity, microbial biomass, and fatty acid profiles[J]. Soil and Tillage Research, 103: 271-281. MOHARANA P C, SHARMA B M, BISWAS D R, et al. 2012. Long-term effect of nutrient management on soil fertility and soil organic carbon pools under a 6-year-old pearl millet-wheat cropping system in an Inceptisol of subtropical India[J]. Field Crops Research, 136(20): 32-41. MONDAL M F, ASADUZZAMAN M D, KOBAYASHI K Y, et al. 2013. Recovery from autotoxicity in strawberry by supplementationof amino acids[J]. Scientia Horticulturae, 164: 137-144. NICOLA RW, JAMES LY, TRAQUAIRB A, et al. 2003. Ginsenosides stimulate the growth of soil-borne pathogens of American ginseng[J]. Phytochemistry, 64(1): 257-264. TIAN YQ, ZHANG X Y, WANG J G. 2013. Soil microbial communities associated with the rhizosphere of cucumber under different summer cover crops and residue management: A 4-year field experiment[J]. Sciential Horticulturae, 150(4): 101-109. WEI X R, HAO M D, SHAO M G, et al. 2006. Changes in soil properties and the availability of soil micronutrients after 18 years of cropping and fertilization[J]. Soil and Tillage Research, 91(1-2): 120-130. WU F Z, WANG X Z, XUE C Y. 2009. Effect of cinnamic acid on soil microbial characteristics in the cucumber rhizosphere[J]. European Journal of Soil Biology, 45(4): 356-362. 张重义, 林文雄. 2009. 药用植物的化感自毒作用与连作障碍[J]. 中国生态农业学报, 17(1): 189-196. 关松荫. 1986. 土壤酶及其研究法[M]. 北京: 农业出版社. 国家药典委员会. 2005. 中华人民共和国药典[M]. 2005年版. 北京: 化学工业出版社. 简在友, 王文全, 游佩进. 2009. 三七连作土壤元素含量分析[J]. 中国现代中药, 11(4): 10-17. 廖海兵, 李云霞, 邵晶晶, 等. 2011. 连作对浙贝母生长及土壤性质的影响[J]. 生态学杂志, 30(10): 2203-2208. 刘恩科, 赵秉强, 李秀英, 等. 2008. 长期施肥对土壤微生物量及土壤酶活性的影响[J]. 植物生态学报, 32(1): 176-182. 刘莉, 王华田, 许坛, 等. 2013. 三七不同间隔年限种植土壤的理化性状比较分析[J]. 西南农业学报, 26(5): 1946-1952. 南京农业大学主编. 1986. 土壤农化分析[M]. 北京: 农业出版社. 苏洁琼, 李新荣, 鲍婧婷. 2014. 施氮对荒漠化草原土壤理化性质及酶活性的影响[J]. 应用生态学报, 25(3): 664-670. 王延平, 王华田, 许坛, 等. 2013. 酚酸对杨树人工林土壤养分有效性及酶活性的影响[J]. 应用生态学报, 24(3): 667-674. 寻路路. 2013. 三七种植不同土壤微生态研究[D]. 杨凌: 西北农林科技大学: 3-15. 杨红飞, 严密, 姚婧, 等. 2007. 铜、锌污染对油菜生长和土壤酶活性的影响[J]. 应用生态学报, 18(7): 1484-1490. 张伟, 刘建国, 龚久平. 2011. 秸秆还田对连作棉田土壤酶活性的影响[J]. 生态环境学报, 20(5): 881-885. 赵玉涛, 李雪峰, 韩士杰, 等. 2008. 不同氮沉降水平下两种林型的主要土壤酶活性[J]. 应用生态学报, 19(12): 2769-2773. 中国科学院南京土壤研究所主编, 1997. 土壤理化分析[M]. 上海: 上海科学技术出版社: 142-150, 196-231. Properties of Soil Physical-Chemistry and Activities of Soil Enzymes in Context of Continuous Cropping Obstacles for Panax notoginseng SUN Xueting1,2, LONG Guangqiang2, ZHANG Guanghui2, MENG Zhengui2, CHEN Zhongjian3, YANG Shengchao2, CHEN Junwen1,2* In order to evaluate the effect of continuous cropping of Panax notoginseng on soil physical-chemical and activities of soil enzymes, in this paper, physical-chemical properties and enzyme activities were analyzed in rhizosphere soil collected from plots where P. notoginseng has been cropped for different years and in soil that has been managed with different remedies. The objectives of the present study were to explain the underlying mechanism of continuous cropping obstacle and to provide a theoretical basis for a method to relieve and overcome the continuous cropping obstacle of P. notoginseng. The result shows that soil pH decreased and soil acidification increased as the cropping time increased. When compared with the uncultivated soil of organic matter, 1-, 2-, and 3-year-cultivated soil of organic matter were decreased by 73%, 87% and 83% , and the cultivated soil had a tendency for hardening. The content of N and P were significantly decreased in the cultivated soil and diseased soil when compared with the uncultivated soil and healthy soil. While the contents of Fe, B and Al were significantly increased by 29%, 31%, 32% respectively, when compared with the uncultivated soil. Acvitivies of protease, catalase, sucrase, phosphatase, and urase in soil significantly decreased as the cropping time proceeded. Activities of protease, catalase, invertase, urease and phosphatase were decreased by 91%, 56%, 52%, 92%, 92% and 69% in the 3-year-cultivated soil of P. notoginseng than in the uncultivated soil of ones. Activities of catalase, sucrase and phosphatase in diseased soil were higher than those in healthy soil, and the activity of invertase in the former was higher than in the latter even by 42%. In comparison with the continuous cropping soil, the disinfection-treated soil showed the significantly higher activities of urease, protease, polyphenol oxidase and catalase. In addition, the activities of enzyme in soil were significantly correlated with soil pH and contents of organic matter, N, P, K, and some microelements (e.g. Mo and Cu). The results obtained here indicated that continuous cropping of P. notognseng aggravate soil acidification and hardening and result in the imbalance of soil mineral elements and the enrichment of some elements, this may induce the continuous cropping obstacle of P. notoginseng. On the other hand, the activities of enzymes in soil were inhibited by the continuous cropping, this would hinder the absorption efficiency of P. notoginseng for some elements and increase the accumulation of soil autotoxic compounds, this may also induce the continuous cropping obstacle. Panax notoginseng; continuous cropping obstacle; soil acidification; mineral element; soil enzymes 10.16258/j.cnki.1674-5906.2015.03.007 S153.6, S154.2 A 1674-5906(2015)03-0409-09 孙雪婷,龙光强,张广辉,孟珍贵,陈中坚,杨生超,陈军文. 基于三七连作障碍的土壤理化性状及酶活性研究[J]. 生态环境学报, 2015, 24(3): 409-417. SUN Xueting, LONG Guangqiang, ZHANG Guanghui, MENG Zhengui, CHEN Zhongjian, YANG Shengchao, CHEN Junwen. Properties of Soil Physical-Chemistry and Activities of Soil Enzymes in Context of Continuous Cropping Obstacles for Panax notoginseng [J]. Ecology and Environmental Sciences, 2015, 24(3): 409-417. 国家自然科学基金项目(81360609);云南省中青年学术技术带头人后备人才培养项目(2014HB011);云南省科技计划项目(2012CG024) 孙雪婷(1989年生),女,硕士研究生,主要从事三七自毒物质与连作障碍的研究。E-mail: 15368783223@163.com *通信作者:陈军文(1978年生),男,副教授,主要从事作物栽培生理生态学研究。E-mail:cjw31412@163.com 2014-12-032 结果与分析




3 讨论
4 结论
1. College of Agriculture and Biotechnology, Yunnan Agriculture University, Kunming 650201, China; 2. Yunnan Agriculture University, Yunnan Provincial Research Center on Good Agricultural Practice for Dominant Chinese Medicinal Materials, Kunming 650201, China; 3. Institute of Sanqi, Wenshan University, Wenshan 663000, China
