人LC3B基因的真核表达及鉴定
2015-11-29董倩徐小洁纪贝贝范忠义梁迎春王涛叶棋浓胡毅
董倩 ,徐小洁,纪贝贝,范忠义,梁迎春,王涛,叶棋浓,胡毅
1.解放军总医院 肿瘤内一科,北京 100853;2.军事医学科学院 生物工程研究所,北京 100850
自噬是真核生物细胞内特有的一种“自食”的现象。目前普遍认为自噬是一种应激调控和防御机制,其实质是细胞通过胞内降解来维持细胞稳态的过程[1],即双层膜包裹部分胞质、细胞内需要降解的大分子物质(如错误折叠的蛋白质)、细胞器(如内质网、线粒体)等形成自噬体,它再与附近的溶酶体融合形成自噬溶酶体,通过水解酶来降解所包裹的内容物,产生氨基酸、ATP、核酸、脂肪酸、糖等来维持细胞生存[2-3]。研究发现自噬对肿瘤的发生、发展及治疗等具有多方面的影响[4],但其分子机制仍然在不断探索中。
自噬发生的重要生物学标志是微管相关蛋白LC3Ⅱ或LC3B。微管相关蛋白1 轻链3(microtubule associated protein 1 light chain 3,简称LC3)是酵母Atg8 的同源蛋白[5],可作为自噬发生的重要检测标志之一[6]。已证实哺乳动物LC3 有LC3A、LC3B 和LC3C 共3 种亚型,各具有不同的功能[7],其中LC3B与自噬发生相关,常被作为标记物。一般情况下,通过蛋白质印迹及免疫荧光等实验观察LC3B蛋白水平的变化,可间接判断自噬是否发生。在此,我们拟构建LC3B 的真核表达载体,同时验证LC3B的生物活性,为进一步探讨细胞自噬奠定基础。
1 材料和方法
1.1 材料
人胚肾细胞293T、大肠杆菌DH5α、pcDNA3.0载体(本实验室保存);HA Beads、HRP 标记的抗HA标签鼠单克隆抗体(Sigma公司);VigoFect(威格拉斯生物技术有限公司);限制性内切酶、DNA 连接酶、PCR 试剂(TaKaRa 公司);PCR 回收试剂盒、质粒提取、胶回收试剂盒(Promega 公司);DMEM、小牛血清(Gibco公司);引物由北京赛百盛生物技术有限公司合成;测序由北京奥科生物技术有限责任公司完成。
1.2 人LC3B基因编码序列的扩增
以人乳腺文库为模板,根据文献报道的LC3编码序列设计上游引物(5'-CGGGATCCATGCCGTCGGAGA AGACCTTCA-3')和下游引物(5'-CGGAATTCTTACAC TGACAATTTCATCC-3'),并在上、下游引物中分别引入BamHⅠ、EcoRⅠ酶切位点,按以下条件PCR扩增人LC3B的编码序列:预变性(95℃,5 min);变性(95℃,30 s)、退火(56℃,30 s)、延伸(72℃,30 s),30个循环;延伸(72℃,7 min)。扩增产物于4℃保存,经10 g/L琼脂糖凝胶电泳后,用DNA胶回收试剂盒回收目的片段。
1.3 HA-LC3B重组质粒的构建和鉴定
回收扩增产物,用BamHⅠ和EcoRⅠ双酶切,暴露出带有粘性的末端,用T4DNA 连接酶于16℃将PCR 回收片段与载体pcDNA3.0-HA 进行连接过夜,连接产物转化大肠杆菌DH5α,于选择性LB 固体培养基(含氨苄西林)上生长,随机挑克隆,37℃培养,取微量菌液保存,剩余菌液提取质粒,并用BamHⅠ和EcoRⅠ双酶切鉴定,酶切产物于20 g/L 琼脂糖凝胶中电泳,约400 bp 处有条带者表明目的片段插入成功,构建成功的克隆由北京奥科生物公司测序。
1.4 质粒转染和Western印迹检测
按本实验室方法进行转染。293T 细胞于含1‰双抗、10%胎牛血清的DMEM 培养液中培养,将其接种于6 cm 皿中,于37℃、5% CO2细胞培养箱中培养24 h 后进行转染。转染时以细胞密度达到80%~90%为宜,转染前1 h 换液。将重组质粒(5 μg)与NaCl(200 μL)混合,VigoFect 与NaCl 混合5 min,再将以上2种溶液混匀,室温静置15 min,把混合物加到已接种293T 细胞的6 cm 皿中,同时转染空载体pcDNA3.0-HA 作为对照组,于细胞培养箱中培养,4~6 h 后更换为正常DMEM 培养液,24 h 后收集细胞,用1×PBS 洗1 次,室温3000 r/min 离心5 min,吸弃上清,将上样缓冲液(2×SDS)加入沉淀中,沸水浴15 min,室温12 000 r/min 离心2 min,上清液经SDS-PAGE 后电转移至硝酸纤维素膜上,用脱脂奶粉(5%)封闭1 h,加入HRP 标记的抗HA 标签鼠单克隆抗体,室温孵育1 h,TBST 洗膜3 次,每次5 min,化学发光法显色3 min,暗室压片显影。
1.5 细胞免疫共沉淀实验
293T 细胞培养于含1‰双抗、10%胎牛血清的DMEM 培养液中,接种于6 cm 皿,分别将空载体HA+Flag-Atg4B、重组质粒HA-LC3B 与Flag-Atg4B共转染其中,于37℃、5% CO2细胞培养箱中,4~6 h后更换为正常DMEM 培养液,24 h 后收集细胞,用1×PBS洗1次,加入用0.5 mL 1×PBS刮下的细胞,收集入EP 管,4℃、3000 r/min 离心5 min,弃上清,加入0.5 mL 中盐IP 缓冲液混匀细胞,置冰上30 min,于冰上行超声波裂解,4℃、12 000 r/min 离心10 min,取上清与HA Beads 于4℃旋转结合4 h 以上,4℃、3000 r/min 离心5 min,弃上清,再次用中盐IP缓冲液漂洗,加入与HA Beads 等量的上样缓冲液(2×SDS),煮沸15 min,12 000 r/min 离心2 min,上清液经SDS-PAGE 后电转移至硝酸纤维素膜上,用脱脂奶粉(5%)封闭1 h,加入HRP 标记的抗HA 标签鼠单克隆抗体,室温孵育1 h,TBST 洗膜3 次,每次5 min,化学发光法显色3 min,暗室压片显影。
2 结果
2.1 人LC3B编码序列的克隆
以人乳腺文库为模板PCR 扩增LC3B 的编码序列,获得目的片段约400 bp,与预期相符(图1)。
2.2 重组质粒HA-LC3B的构建与鉴定
用BamHⅠ和EcoRⅠ酶切重组质粒HA-LC3B,经10 g/L 琼脂糖电泳后显示2 条片段,其中一条约400 bp,而酶切空载体只出现1 条片段,与预期结果相符(图2)。重组质粒的测序显示插入片段的DNA序列与人LC3B基因的编码序列完全一致,从而证明重组质粒HA-LC3B构建成功(序列略)。
2.3 Western印迹检测融合蛋白的表达
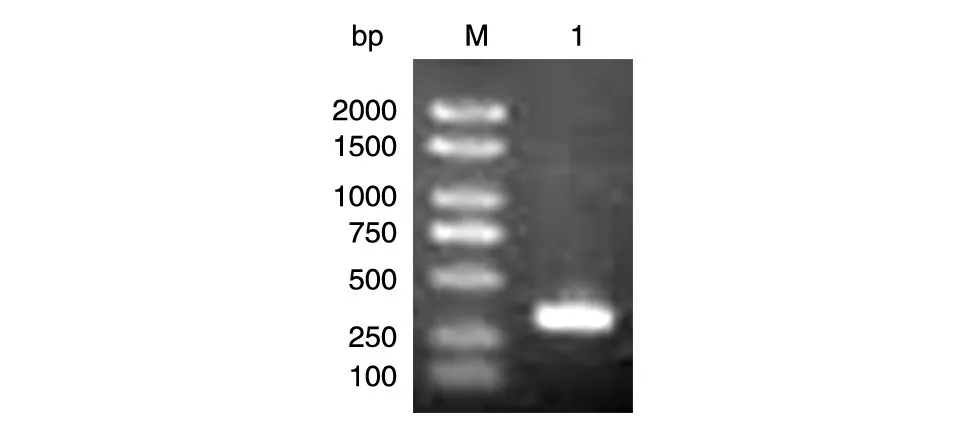
图1 目的基因LC3B的PCR扩增
将构建的重组质粒HA-LC3B 和空载体分别转染293T 细胞,4~6 h 后更换为正常DMEM 培养液,24 h后提取细胞蛋白,行SDS-PAGE 检测HA-LC3B蛋白的表达。结果显示,对293T 细胞转染重组质粒HA-LC3B 后,用HA-HRP 抗体,在相对分子质量约16×103处可见明显的特异性条带,表明重组蛋白HA-LC3B能在293T细胞中正确表达(图3)。
2.4 细胞免疫共沉淀实验结果
据报道LC3B 可与Atg4B 蛋白相互作用[8],据此可检测HA-LC3B 融合蛋白的生物学活性。细胞免疫共沉淀实验结果表明,在符合Atg4B 蛋白(相对分子质量约47 000)的位置显示特异性条带(图4),而HA 空载体蛋白在同一位置无条带,说明HA-LC3B蛋白与Atg4B 蛋白能特异地相互作用,且其结构及生物学功能并不受HA标签的影响。
3 讨论
上世纪50 年代,de Duve 通过电镜观察到自噬体结构[9]。自噬现象对于细胞生长是柄双刃剑,过多或过少都会影响细胞生长。细胞自噬是一个动态发展的过程,由一系列自噬相关蛋白(Atg 蛋白)介导完成[10],其中Atg8 泛素样蛋白修饰过程对自噬体的形成必不可少[11]。LC3 是在哺乳动物细胞中发现的酵母Atg8 的同源体,分为Ⅰ型和Ⅱ型,Ⅰ型位于胞质中,为可溶性蛋白,Ⅱ型则位于自噬体膜上。在自噬过程中,LC3Ⅰ与磷脂酰乙醇胺结合后形成LC3Ⅱ即LC3B,被聚集招募结合于自噬体膜上。自噬体与溶酶体融合形成自噬溶酶体,LC3B 在自噬溶酶体内将被降解。LC3B 的含量与自噬体的数量成正相关,因此,细胞中LC3B的含量变化可以直接反映自噬的发生过程[12]。已知人类LC3的3种亚型中LC3B与自噬密切相关[7],利用生化技术如Western 印迹检测LC3B 水平常作为监测自噬水平的有效方法[10,13-14]。同时,在自噬过程中Atg4B的作用也不容忽视[15]。
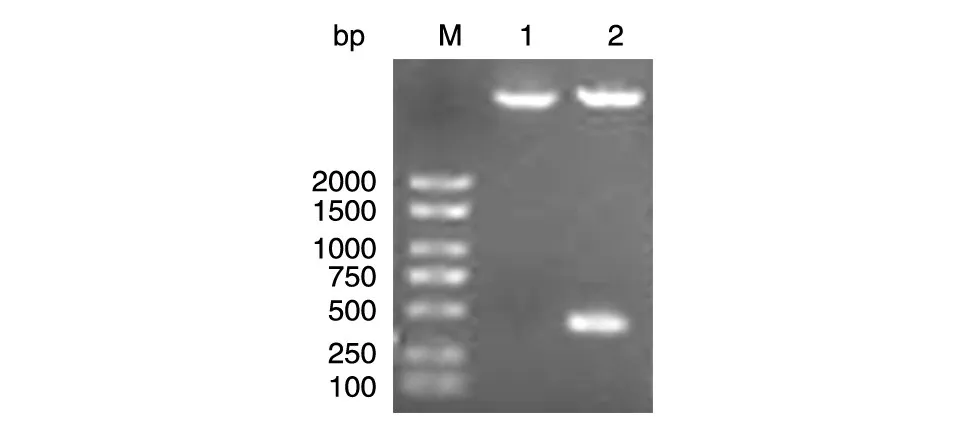
图2 重组质粒HA-LC3B的BamHⅠ/EcoRⅠ双酶切电泳图谱
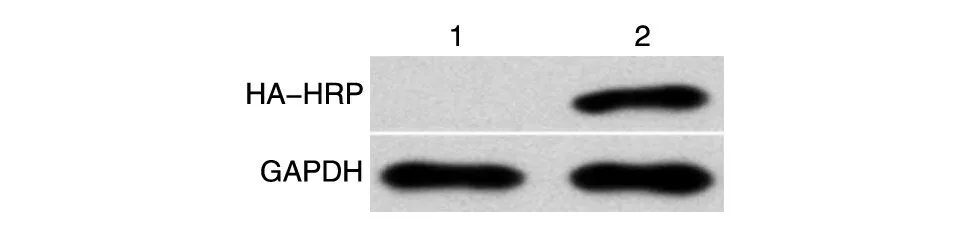
图3 Western印迹检测融合蛋白HA-LC3B的表达
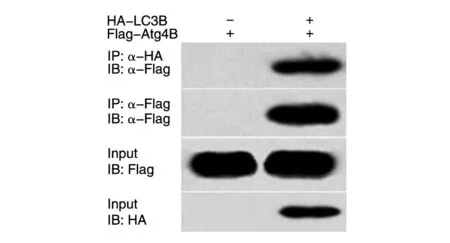
图4 细胞免疫共沉淀实验检测HA-LC3B和Atg4B的相互作用
综上,我们构建了HA-LC3B 真核表达载体,并证实Atg4B 可与LC3B 相互作用。LC3B 在真核细胞中的成功表达,为今后研究细胞自噬奠定了基础。
[1]Tooze S A,Yoshimori T.The origin of the autophagosomal membrane[J].Nat Cell Biol,2010,12(9):831-835.
[2]Komatsu M,Ichimura Y.Selective autophagy regulates various cellular functions[J].Genes Cells,2010,15(9):923-933.
[3]Rabinowitz J D,White E.Autophagy and metabolism[J].Science,2010,330(6009):1344-1348.
[4]Choi A M,Ryter S W,Levine B.Autophagy in human health and disease[J].N Engl J Med,2013,368(19):1845-1846.
[5]Marino G,Niso-Santano M,Baehrecke E H,et al.Self-consumption:the interplay of autophagy and apoptosis[J].Nat Rev Mol Cell Biol,2014,15(2):81-94.
[6]李玲,徐小洁,叶棋浓.细胞自噬与肿瘤[J].中国生物化学与分子生物学报,2013,11(2):995-1001.
[7]He H,Dang Y,Dai F,et al.Post-translational modifications of three members of the human MAP1LC3 family and detection of a novel type of modification for MAP1LC3B[J].Biol Chem,2003,278(31):29278-29287.
[8]Satoo K,Noda N N,Kumeta H,et al.The structure of Atg4B-LC3 complex reveals the mechanism of LC3 processing and delip-idation during autophagy[J].EMBO J,2009,28(9):1341-1350.
[9]Eskelinen E L,Reggiori F,Baba M,et al.Seeing is believing:the impact of electron microscopy on autophagy research.Autophagy,2011,7(9):935-956.
[10]Barth S,Glick D,Macleod K F.Autophagy:assays and artifacts[J].J Pathol,2010,221(2):117-124.
[11]Sou Y S,Waguri S,Iwata J,et al.The Atg8 conjugation system is indispensable for proper development of autophagic isolation membranes in mice[J].Mol Biol Cell,2008,19(11):4762-4775.
[12]Tanida I,Ueno T,Kominami E.LC3 and autophagy[J].Methods Mol Biol,2008,445:77-88.
[13]Kabeya Y,Mizushima N,Ueno T,et al.LC3,a mammalian homologue of yeast Apg8p,is localized in autophagosome membranes after processing[J].EMBO J,2000,19(21):5720-5728.
[14]Hansen T E,Johansen T.Following autophagy step by step[J].BMC Biol,2011,9:1-4.
[15]黄蓉,杜楠,叶棋浓,等.人自噬相关基因LC3B 原核表达载体的构建及活性检测[J].军事医学,2014,11(38):867-870.
