SIRT2的亚细胞定位及其对293T细胞增殖的影响
2015-11-29王旋仲念念赵盼雄季少平
王旋,仲念念,赵盼雄,季少平
河南大学 医学院,河南 开封 475004
Sirtuins 是NAD+依赖的蛋白质去乙酰化酶[1-2],在哺乳动物中由7个成员组成,即SIRT1~SIRT7[3],包含一个约275 个氨基酸的保守催化核心区[4-5]。它们调节许多细胞功能,例如基因组维护、寿命和新陈代谢。已明确SIRT1 和SIRT6 和DNA 修复或DNA 损伤反应信号通路相连[6]。除了SIRT4,其他成员已经被认为具有去乙酰化作用[7]。据报道,SIRT2 去乙酰化非组蛋白蛋白[8],如α微管蛋白[9]、p53[10]、钙黏着蛋白1(Cdh1)[11]、细胞分裂周期蛋白20(CDC20)等,参与调节细胞稳态。SIRT2 在肿瘤发生过程中也扮演了重要角色,有时促进肿瘤的生成,有时抑制肿瘤的生成,这取决于细胞环境或肿瘤种类[8]。SIRT2 还是抑制细胞分化的少突神经胶质蛋白[12],在大脑中高度表达[13]。蛋白质功能的发挥通常与其亚细胞定位密切相关[13],它们常通过在不同亚细胞环境里的运动发挥作用。故而,进行细胞亚定位研究,将为揭示SIRT2蛋白的功能提供实验依据。
1 材料与方法
1.1 材料
人胚肾细胞株293T 由本实验室保存;大肠杆菌Top10、带有目的基因的克隆载体为本实验室保存;DMEM 来自GIBCO 公司;胎牛血清来自浙江天杭生物科技有限公司;蛋白定量BCA 试剂盒购自康为世纪生物科技有限公司;出核转运抑制剂细霉素B(leptomycin B)购自碧云天生物技术研究所;anti-SIRT2 抗体购自Sigma 公司;anti-eGFP 抗体、anti-LMNB1 抗体购自武汉三鹰生物技术有限公司;antiβ-actin 抗体、羊抗兔二抗购自武汉博士德生物工程有限公司;质粒小量抽提试剂盒及凝胶回收试剂盒购自康为世纪生物科技有限公司;pMD19-T 载体,限制性核酸内切酶EcoRⅠ、KpnⅠ,T4DNA 连接酶,DNA 分子量标准DL2000 均购自大连宝生物工程有限公司;DNA分子量标准Marker Ⅳ购自上海莱枫生物科技有限公司;阳离子脂质体LipofectAMINE 2000 购自Invitrogen 公司;核浆分离试剂(PBS,Tris,氯化镁,NP-40)为国产分析纯产品。
1.2 PCR扩增目的基因及克隆
根据GenBank 中的基因序列(NM_001008368),应用软件Primer Premier 5.0设计用于克隆SIRT2基因的上游引物(5'-GAATTCGAGATGGACTTCCTAC GG-3')和下游引物(5'-GGTACCCTAGTGTTCCTCT TTCTCTTTG-3'),上、下游引物中分别引入EcoRⅠ、KpnⅠ酶切位点,扩增片段为1052 bp。引物由金唯智生物科技有限公司合成。
以本实验室的SIRT2-pIRES为模板进行PCR反应。反应体系:模板1 μL,10×PCR 缓冲液5 μL,dNTP(2.5 mmol/L)1 μL,灭菌蒸馏水40.5 μL,rTaq(5 U/μL)0.5 μL,上、下游引物(10 μmol/L)各1 μL。反应条件:94℃5 min;94℃40 s,56℃40 s,72℃2 min,扩增32 个循环。PCR 产物经琼脂糖凝胶电泳分析鉴定,切胶回收后纯化。用pMD19-T 载体连接试剂盒(TaKaRa 公司)将回收纯化的PCR 产物与pMD19-T 载体连接(SolutionⅠ5 μL,pMD19-T DNA 0.5 μL,PCR产物4.5 μL,16℃反应2 h),取连接产物3 μL 转化感受态大肠杆菌Top10,经蓝白斑筛选出阳性重组子,小量提取质粒DNA,EcoRⅠ、KpnⅠ双酶切鉴定有无插入片段,并取10 μL质粒送生工公司测序。
1.3 重组质粒SIRT2-pEGFP-C2的构建及鉴定
用EcoRⅠ、KpnⅠ分别双酶切重组pMD19-T/SIRT2质粒及pEGFP-C2空载体,凝胶电泳后回收目的基因片段,用T4DNA 连接酶将pEGFP-C2 和SIRT2 目的片段在16℃反应过夜,连接产物转化大肠杆菌Top10,挑选重组质粒,用EcoRⅠ和KpnⅠ双酶切鉴定。
1.4 重组质粒在293T细胞中的表达
293T 细胞系在含10%胎牛血清的DMEM 培养基中,于37℃、5% CO2饱和湿度的孵箱中培养。
按照LipofectAMINE 2000 的说明书分别转染SIRT2-pEGFP-C2 和pEGFP-C2 至293T 细胞,并设立空白对照,24 h 后在荧光显微镜下观察,48 h 后收集细胞进行Western印迹。
1.5 SIRT2的亚细胞定位
将细胞接种在60 mm 的细胞培养皿中,分为7组;待各培养皿中的细胞长到50%~60%时,用LipofectAMINE 2000 转染SIRT2-pEGFP-C2,同时设置pEGFP-C2 的空载体对照,24 h 后在荧光显微镜下观察;加入细霉素B后5 h,在荧光显微镜下观察;将培养基换成含10%胎牛血清的DMEM 后培养19 h,在荧光显微镜下观察;收集细胞,用核浆分离试剂分离浆蛋白和核蛋白,并进行Western印迹。
分组如下:①转染SIRT2-pEGFP-C2;②转染pEGFP-C2;③未转染的细胞;④转染SIRT2-pEGFP-C2后加细霉素B;⑤转染pEGFP-C2后加细霉素B;⑥转染SIRT2-pEGFP-C2,加细霉素B 5 h 后更换为完全培养基;⑦转染pEGFP-C2,加细霉素B 5 h后更换为完全培养基。
1.6 MTT法检测SIRT2对293T细胞增殖的影响
将293T 细胞接种于6 孔板中,分别转染SIRT2-pEGFP-C2 和pEGFP-C2,并设立空白对照组。转染18 h 后消化细胞,分别按6000/孔重新接种96 孔板,24 h 后加细霉素B(终浓度为20 ng/mL),分组为a(转染SIRT2-pEGFP-C2 未加细霉素B)、b(转染pEGFP-C2 未加细霉素B)、c(空白对照未加细霉素B)、d(转染SIRT2-pEGFP-C2 并加细霉素B)、e(转染pEGFP-C2 并加细霉素B)、f(空白对照并加细霉素B),每组4 个平行孔,继续培养24 h,每孔加入200 μL 新鲜的DMEM 及20 μL 浓度为5 mg/mL 的MTT,避光温育4 h。弃去孔中培养基和MTT,所有孔中加入150 μL DMSO,振荡10 min,测定D490nm值并计算细胞增殖抑制率,用不含细胞的孔调零。实验重复4次。
细胞增殖抑制率=(1-实验组D490nm/空白对照组D490nm)×100%
1.7 统计学处理
用SPSS 统计软件进行数据处理,采用单因素方差分析和LSD-t检验及两独立样本的t检验。实验数据用x±s的形式表示。
2 结果
2.1 目的基因的获得
PCR 扩增获得SIRT2 编码基因,经琼脂糖凝胶电泳鉴定,产物约1000 bp,与预期大小一致(图1)。
2.2 T载体克隆的鉴定
目的基因与T 载体连接后,质粒双酶切产物经琼脂糖凝胶电泳鉴定与目的基因大小相符。质粒对应的测序结果显示,装入T 载体的序列与GenBank上登录的序列也相符(图2)。
2.3 真核表达载体的构建与鉴定
用EcoRⅠ和XhoⅠ双酶切重组质粒得到的目的基因插入pEGFP-C2 空质粒,获得真核表达质粒,该质粒再经EcoRⅠ和XhoⅠ双酶切鉴定,片段大小正确,表明构建成功(图3)。
2.4 重组质粒在293T细胞中的表达
将重组质粒转染293T细胞,48 h后可见绿色荧光主要分布在细胞质中,而转染空载体的绿色荧光基本均匀分布(图4)。收集细胞进行Western 印迹,发现转染重组质粒的细胞高表达SIRT2 蛋白,转染空载体和没有转染的细胞几乎不表达SIRT2 蛋白(图5)。
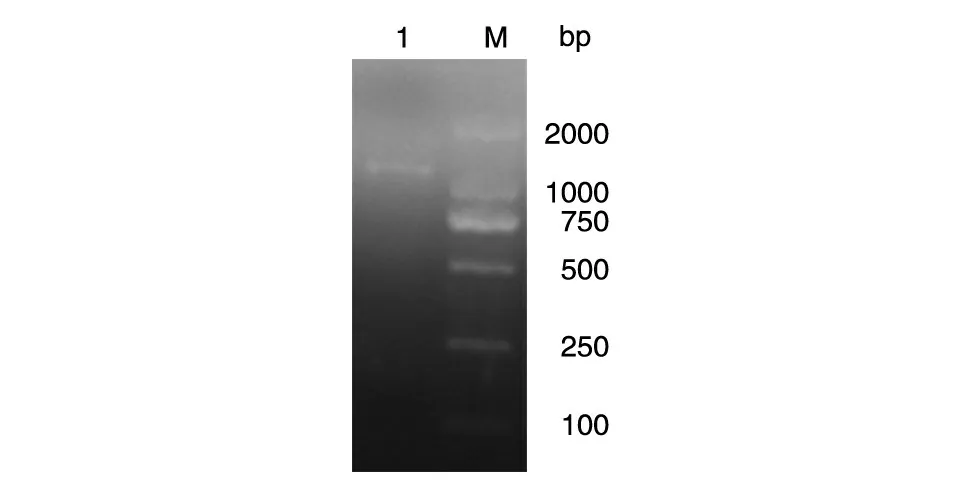
图1 SIRT2目的基因的PCR产物
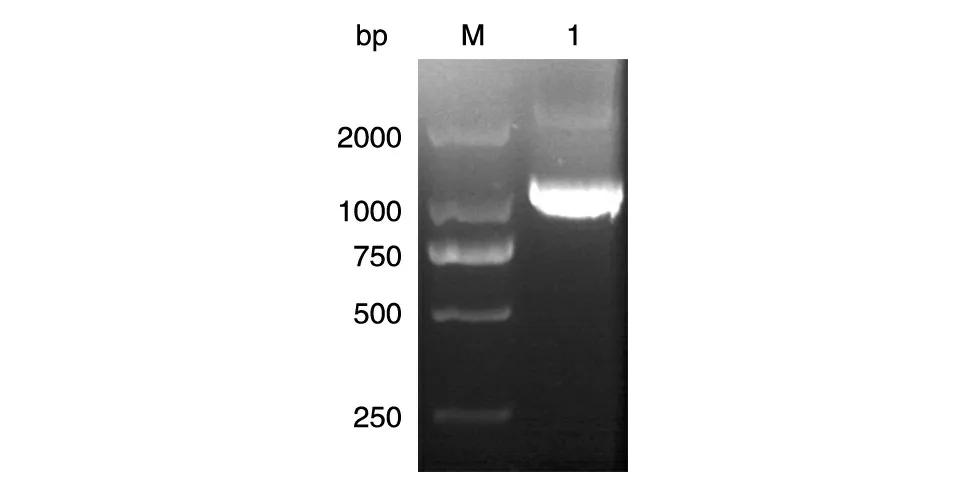
图2 T-SIRT2载体的鉴定
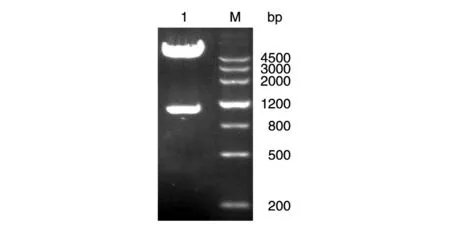
图3 SIRT2-pEGFP-C2载体的鉴定
2.5 SIRT2的亚细胞定位
在显微镜下观察发现,293T 细胞中转染SIRT2-pEGFP-C2 后,SIRT2 主要分布在细胞质中,加入细霉素B 后,SIRT2 在核内有明显聚集,撤去细霉素B后,SIRT2 还是主要分布在细胞质中(图6);进一步通过Western 印迹检测发现,转染SIRT2-pEGFP-C2后SIRT2 主要在细胞质中表达,细胞核内几乎没有,加入细霉素B 后,SIRT2 在细胞质中依旧表达很高,在核内的表达也明显升高,撤去细霉素B 后,和加细霉素B之前的情况基本一致(图7)。
2.6 SIRT2对293T细胞增殖的影响
组a 和组b 间差别显著(P<0.05,n=4),组a 和组c 间差别显著(P<0.05,n=4),组b 和组c 间无显著差别,说明转染SIRT2后细胞增殖受到抑制。组d和组e 间差别显著(P<0.05,n=4),组d 和组f 间差别显著(P<0.05,n=4),组e 和组f 间无显著差别,说明转染SIRT2 并加入细霉素B 后,细胞增殖仍然受到抑制。组a 和组c 间差别显著(P<0.05,n=4),组b 和组d 间无显著差别,说明SIRT2 进入核后对细胞增殖的抑制作用减弱。
3 讨论
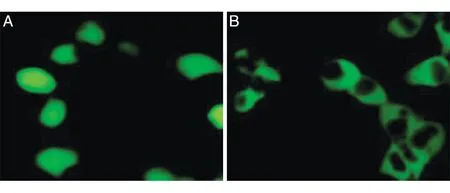
图4 荧光显微镜下观察SIRT2在293T细胞中的表达(400×)
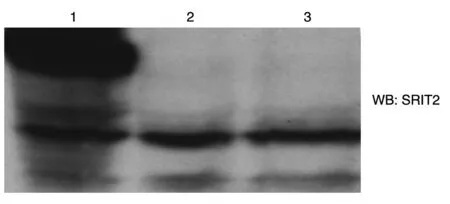
图5 Western印迹检测SIRT2在293T细胞中的表达
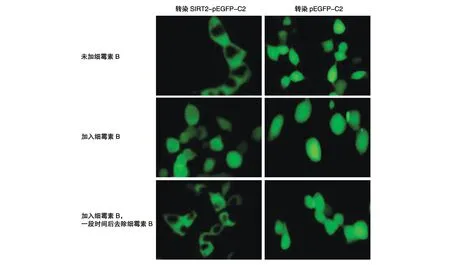
图6 荧光显微镜下观察细霉素B对SIRT2亚细胞定位的影响(400×)
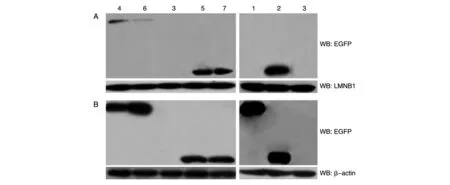
图7 Western印迹分析细霉素B对SIRT2亚细胞定位的影响
蛋白质是生命活动的基础,对生物体来说具有至关重要的作用。由于细胞各部分都有特定的蛋白质组分,合成的蛋白质必须定向地、准确无误地进行运送,才能保证细胞活动的正常进行。不同的蛋白质通常分布在细胞的不同部位,具有不同的功能。即使同一个蛋白在不同的组织细胞中及在不同的生长发育阶段,也可能具有不同的功能[14]。因此,要想真正了解蛋白质的功能,通常还需要知道蛋白质所处的空间位置。并且,了解蛋白质的亚细胞定位信息,可对蛋白质的其他研究如相互作用、进化等也能提供必要的信息[15]。研究表明,蛋白质的功能与其亚细胞定位密切相关,因此阐明蛋白质的定位情况将为其功能研究提供重要的理论依据。例如,有研究指出,随着p21 由细胞核到细胞质移位,肝癌肿瘤细胞的恶性程度增加,肿瘤转移加快。可见,了解SIRT2 的亚细胞定位情况,对于进一步研究其功能具有重大的指导意义。
蛋白和mRNA 在细胞核和细胞质之间的运输称为入核和出核,它是维持细胞动态稳定的重要因素。这种运输需要一些其他蛋白因子的辅助,并要求有一特定的可识别定位序列。蛋白和mRNA 的运输则需要信号介导并消耗能量。CRM1(染色体区维持蛋白1)是主要的出核介导物。细霉素B是一种抗真菌化合物,通过抑制NES(核输出信号)和CRM1的相互作用,强效地、特异性地阻止NES相关的核蛋白输出[16]。
GFP 能自发荧光,不必外加底物或破碎组织,不需要目的蛋白的抗体或原位杂交技术,通过荧光显微镜即可观察与荧光蛋白融合的蛋白在活细胞的空间和时间定位。GFP 已被广泛应用于基因表达调控、蛋白定位、迁移变化、转基因动物等基因功能方面的研究[17]。EGFP 是GFP 的突变型,其荧光强度比GFP 高4~35 倍。通过构建目的基因与绿色荧光蛋白的融合表达载体对蛋白进行亚细胞定位研究,灵敏度高,易操作,实验周期短,很适于活体实时定位和动态研究。
为明确并验证SIRT2 的亚细胞定位情况,我们以EGFP 为标记物,分析了SIRT2 在293T 细胞中的表达、定位情况。荧光显微镜观察结果及Western印迹结果显示,转染SIRT2-pEGFP-C2 后,SIRT2 主要分布在细胞质中,加入细霉素B 后,SIRT2 在细胞质中依旧表达很高,在核内的表达也明显升高,撤去细霉素B 后,SIRT2 主要分布在细胞质中。这表明SIRT2 具有核质穿梭功能,它的亚细胞定位是动态变化过程,为进一步研究SIRT2的功能奠定了基础。
MTT 实验检测细胞活力的结果表明SIRT2 对细胞增殖起抑制作用,其亚细胞定位影响抑制作用的强弱。然而,这个结论还需要多方面的验证,并且SIRT2 的亚细胞定位对其他功能的影响也有待于进一步研究。
[1]Haigis M C,Sinclair D A.Mammalian sirtuins:biological insights and disease relevance[J].Annu Rev Pathol,2010,5:253-295.
[2]de Oliveira R M,Sarkander J,Kazantsev A G,et al.SIRT2 as a therapeutic target for age-related disorders[J].Front Pharmacol,2012,3:82.
[3]Ji S,Doucette J R,Nazarali A J.SIRT2 is a novel in vivo downstream target of Nkx2.2 and enhances oligodendroglialcell differentiation[J].J Mol Cell Biol,2011,3(6):351-359.
[4]Saunders L R,Verdin E.Sirtuins:critical regulators at the crossroads between cancer and aging[J].Oncogene,2007,26(37):5489-5504.
[5]Michan S,Sinclair D.Sirtuins in mammals:insights into their biological function[J].Biochem J,2007,404(1):1-13.
[6]Zhang H,Park S H,Pantazides B G,et al.SIRT2 directs the replication stress response through CDK9 deacetylation[J].Proc Natl Acad Sci USA,2013,110(33):13546-13551.
[7]Rothgiesser K M,Erener S,Waibel S,et al.SIRT2 regulates NF-kappa B dependent gene expression through deacetylation of p65 Lys310[J].J Cell Sci,2010,123(Pt24):4251-4258.
[8]Cha Y I,Kim H S.Emerging role of sirtuins on tumorigenesis:possible link between aging and cancer[J].BMB Rep,2013,46(9):429-438.
[9]North B J,Marshall B L,Borra M T,et al.The human Sir2 ortholog,SIRT2,is an NAD+-dependent tubulin deacetylase[J].Mol Cell,2003,11(2):437-444.
[10]Jin Y H,Kim Y J,Kim D W,et al.SIRT2 interacts with 14-3-3 beta/gamma and down-regulates the activity of p53[J].Biochem Biophys Res Commun,2008,368(3):690-695.
[11]Kim H S,Vassilopoulos A,Wang R H,et al.SIRT2 maintains genome integrity and suppresses tumorigenesis through regulating APC/C activity[J].Cancer Cell,2011,20(4):487-499.
[12]Li W,Zhang B,Tang J,et al.Sirtuin 2,a mammalian homolog of yeast silent information regulator-2 longevity regulator,is an oligodendroglial protein that decelerates cell differentiation through deacetylating alpha-tubulin[J].J Neurosci,2007,27(10):2606-2616.
[13]Donmez G,Outeiro T F.SIRT1 and SIRT2:emerging targets in neurodegeneration[J].EMBO Mol Med,2013,5(3):344-352.
[14]Simpson J C,Pepperkok R.The subcellular localization of the mammalian proteome comes a fraction closer[J].Genome Biol,2006,7(6):222.
[15]张菁,丁小凤,罗畅,等.Dnd1 的蛋白亚细胞定位及其对HeLa细胞增殖的抑制作用[J].中国生物化学与分子生物学报,2009,25(1):37-43.
[16]Shi Y,Zhang L,Yang T.Nuclear export of L-periaxin,mediated by its nuclear export signal in the PDZ domain[J].PLoS One,2014,9(3):e91953.
[17]Valdez B C,Perlaky L,Cai Z J,et al.Green fluorescent protein tag for studies of drug-induced translocation of nucleolar protein RH-II/Gu[J].Biotechniques,1998,24(6):1032-1036.
