小鼠黑色素瘤相关抗原MART1的原核表达及抗体制备
2015-11-29李倩孙鹏杨建林曹春雨王艳林
李倩,孙鹏,杨建林,曹春雨,王艳林
三峡大学 医学院 肿瘤微环境与免疫治疗湖北省重点实验室,湖北 宜昌 443002
鼠黑色素瘤相关抗原(melanoma antigen recognized by T-cells 1,MART1)又称Melan-A,主要存在于正常黑色素细胞或恶性黑色素肿瘤细胞中。作为一种特征性黑素瘤肿瘤抗原,MART1 的表达异常与恶性黑色素瘤发生与发展密切相关[1-2]。最新研究表明,MART1 中的抗原肽能够被T 淋巴细胞识别并激发强烈的CTL 反应,提示MART1在黑色素瘤特异性免疫治疗中潜在的应用价值[3-6]。我们拟在大肠杆菌中原核表达和纯化MART1蛋白,并以此为抗原制备高效价的兔抗鼠多克隆抗体,以为MART1在抗肿瘤免疫治疗中的应用研究提供技术支撑。
1 材料与方法
1.1 材料
日本大耳白兔(雄性,2 kg)购自湖北省实验动物中心;鼠黑色素瘤B16-F1 细胞株、大肠杆菌Rosseta(DE3)、质粒载体pET32a 为本实验室保存;TRIzol 总RNA 纯化试剂由Introvogen 公司提供;TaqDNA 聚合酶、限制性内切酶由Thermo 公司提供,Ni-NTA蛋白纯化树脂购于Qiagen公司;弗氏完全佐剂、弗氏不完全佐剂、抗β-actin 和抗His 标签单抗购自Sigma 公司;Rodamine 标记的山羊抗小鼠IgG 为北京中杉金桥公司产品;HRP 标记的山羊抗小鼠IgG 多克隆抗体购自武汉博士德公司;PCR 引物合成及DNA测序由上海生工生物工程技术有限公司完成。
1.2 RT-PCR克隆小鼠MART1 cDNA
用TRIzol 试剂提取小鼠B16-F1 黑素瘤细胞总RNA,以该RNA 为模板、Oligo(dT)为引物,经逆转录酶M-MLV 催化,将mRNA 逆转录成cDNA 第一链,再以此cDNA 第一链为模板,用PCR 技术扩增出MART1 双链DNA 编码序列。PCR 上下游引物序列分别为5'-CGGAATTCATGCCCCAAGAAGACACAT TCA-3'和5'-AGCAAGCTTTCTCAGGGTGAATAAG GTGG-3',在上下游引物的5'端分别引入酶切位点EcoRⅠ和HindⅢ(引物序列中下划线部分)以便随后的克隆操作。PCR 产物经EcoRⅠ和HindⅢ双酶切后克隆入pET32a 原核表达载体,用酶切和DNA测序鉴定所得重组质粒pET32a-MART1 中插入DNA序列和方向的正确性。
1.3 MART1的原核表达、纯化及鉴定
用质粒pET32a-MART1 转化大肠杆菌Rosseta(DE3)感受态细胞,培养至细菌处于对数生长期时,加入IPTG 至终浓度为0.3 mmol/L,诱导细菌表达重组蛋白5 h;离心收集诱导后的菌体,用裂解缓冲液(10 μmol/L 咪 唑,300 mmol/L NaCl,50 mmol/L NaH2PO4)重悬沉淀,超声波裂解后取上清,上清中的MART1 重组蛋白用Ni-NTA 树脂亲和层析分离,咪唑浓度梯度洗脱获得初步纯化的MART1重组蛋白,再用制备性PAGE 进一步纯化。用Western 印迹鉴定MART1 重组蛋白:样品经SDS-PAGE 分离后电转至PVDF膜,膜用5%脱脂牛奶封闭1 h,与1∶1000稀释的鼠源His 标签抗体共孵育,低速摇床上4℃过夜,再与1∶5000 稀释的HRP 标记的羊抗鼠IgG 室温共孵育1 h,ECL显影指示目标蛋白。
1.4 MART1多克隆抗体的制备
用纯化的MART1 蛋白免疫雄性日本大耳白兔。首次免疫剂量500 μg/只,与等量弗氏完全佐剂混合后于脊柱两旁皮下注射;间隔10 d 后进行第二次免疫,注射剂量为100 μg/只,与等量弗氏不完全佐剂混合后皮下注射;10 d 后同样方法进行第三次免疫;第三次免疫1周后颈静脉取血,分离血清。
1.5 ELISA检测抗体效价
以纯化的MART1 蛋白(2 μg/孔)为包被抗原,一抗为本实验中制备的梯度稀释的兔抗血清,二抗为HRP标记的羊抗兔IgG(1∶5000稀释),TMB显色,全波长酶标仪检测光密度(D450nm)。
1.6 Western印迹检测抗体的特异性
蛋白样品包括IPTG 诱导前、后的大肠杆菌菌液,纯化的MART1 蛋白和B16-F1 细胞裂解液。上述样品经SDS-PAGE 分离后电转至PVDF 膜,膜用5%脱脂牛奶封闭后,先后与本实验自制的兔抗血清(1∶3000 稀释)和HRP 标记的羊抗兔IgG(1∶5000 稀释)共孵育,ECL显影指示目标蛋白。
1.7 制备抗体用于免疫荧光染色
将B16-F1 细胞接种于事先放置有盖玻片的6孔板中制备细胞爬片,培养24 h 后用40 g/L 多聚甲醛于4℃固定15 min,经5 mL/L TxitonX-100 透化处理15 min,10 mL/L山羊血清37℃封闭30 min,然后分别与本实验制备的MART1抗血清(1∶500稀释)和阴性对照(1×PBS 取代一抗)37℃共孵育2 h,加入罗丹明标记的羊抗兔IgG(1∶100 稀释),37℃避光孵育30 min,荧光倒置显微镜下观测并记录结果。
2 结果
2.1 鼠MART1 编码cDNA 的克隆与原核表达载体构建
如1.2所述方法构建重组质粒pET32a-MART1,重组质粒经EcoRⅠ和HindⅢ双酶切,获得的限制性片段长度与理论预期一致(图1),DNA 测序证实pET32a-MART1 中插入的DNA 序列、方向及读框之间的对接均无误。
2.2 MART1重组蛋白的原核表达、纯化及鉴定
将重组质粒pET32a-MART1转化大肠杆菌Rosseta(DE3)感受态细胞并用IPTG 诱导5 h 后,SDSPAGE 分析显示诱导后的细菌中有一新生蛋白条带出现,其分子大小与MART1重组蛋白预期相对分子质量一致(图2)。该蛋白可经Ni-NTA 树脂亲和层析及切胶回收方法有效纯化。Western 印迹分析证实该融合蛋白能与His标签抗体特异性结合(图3)。
2.3 抗MART1抗体的制备及效价鉴定
用上述实验纯化所得重组MART1 蛋白作为抗原免疫日本大耳白兔,经3 次皮下注射后,获取免疫后的兔血清并用ELISA 法分析制备抗体的滴度和特异性。
用上述纯化的同一批次的MART1 蛋白包被ELISA 检测板,将抗体梯度稀释(每个稀释度设3 个复孔)后进行抗原-抗体结合反应,以高出对照血清D450nm值2.1 倍的制备血清抗体的最高稀释度为抗体滴度。结果如图4,本实验所制备抗体的滴度为1∶1 024 000。
2.4 Western印迹鉴定制备抗体的特异性
Western 印迹结果显示,制备的抗体能有效识别和结合原核表达的MART1 蛋白,在相对分子质量23×103处出现明显的特异性反应条带,其分子大小与预期一致(图5)。另外,用16-F1细胞裂解液作为样品进行的分析也能检测出特异性条带,表明本实验制备的抗体能与真核细胞表达的MART1 抗原特异性结合(图5)。
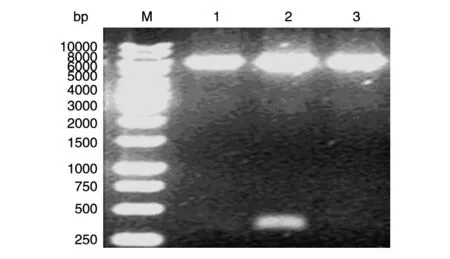
图1 pET32a-MART1质粒的酶切鉴定
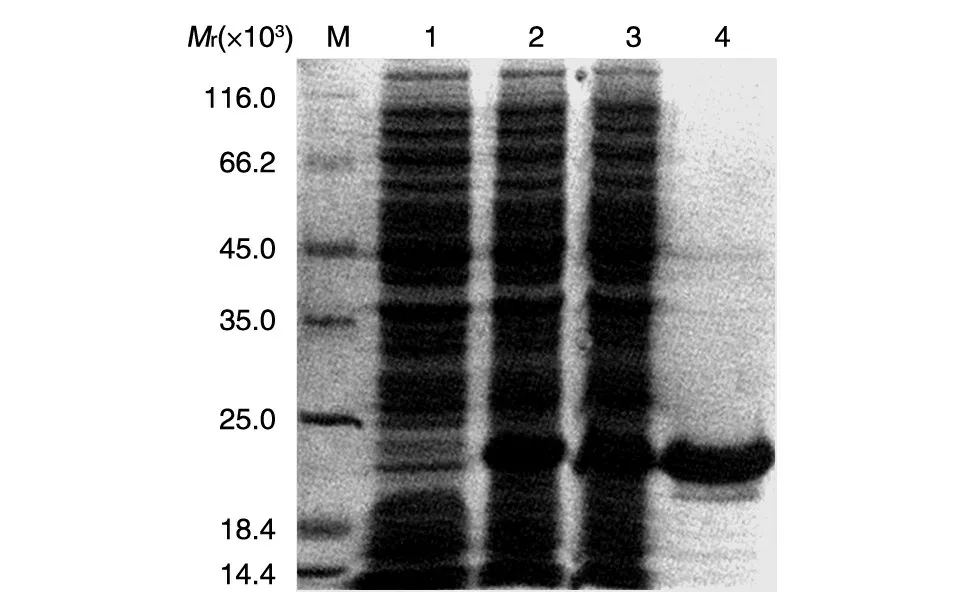
图2 SDS-PAGE分析原核表达及纯化的重组MART1
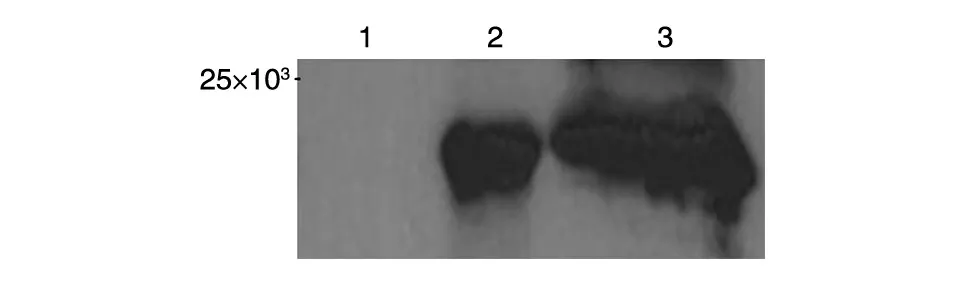
图3 Western印迹分析原核表达和纯化的MART1重组蛋白
同时,还用免疫荧光染色法检测了制备抗体的特异性。制备B16-F1细胞爬片后,先后与制备抗体(一抗)和罗丹明标记的羊抗兔IgG(二抗)共孵育,然后在荧光显微镜下分析结果。结果显示,制备抗体可以有效识别B16-F1细胞中的MART1蛋白(图6)。
3 讨论
为获得大量可用作免疫抗原的小鼠MART1 蛋白,我们利用基因克隆技术将小鼠MART1基因全长编码序列克隆到pET32a载体中,DNA 双向测序证实构建的pET32a-MART1 重组质粒中外源DNA 片段插入方向及其与质粒中6×His 标签编码序列框架对接无误,无移码突变、碱基缺失等。在大肠杆菌原核表达系统中,IPTG 能高效诱导MART1 重组蛋白表达。由于重组蛋白N 端带有6×His 标签,我们用Ni-NTA 树脂亲和层析法纯化MART1 重组蛋白。为得到更高纯度的MART1 重组蛋白,我们将Ni-NTA 树脂纯化所得的蛋白样品用制备性聚丙烯酰胺凝胶电泳进一步纯化。
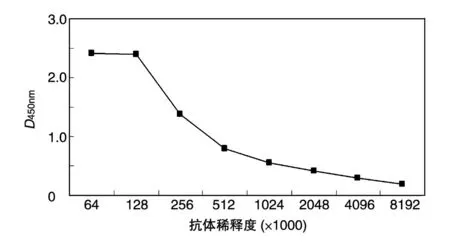
图4 ELISA检测抗体效价
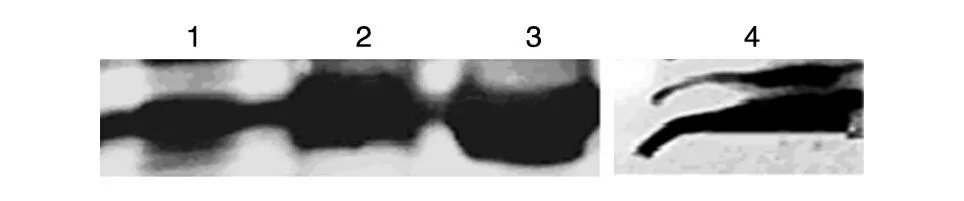
图5 Western印迹检测抗体的特异性
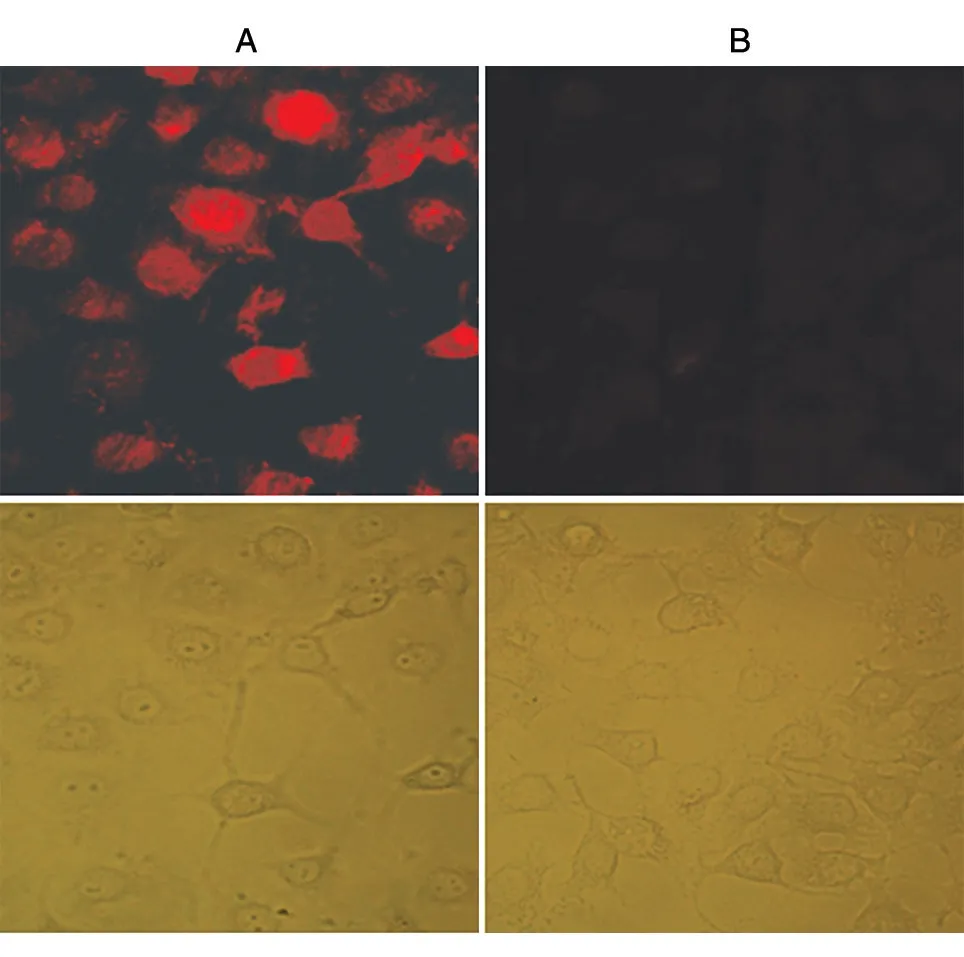
图6 细胞免疫荧光分析制备抗体的特异性(×400)
为获得抗MART1的多克隆抗体,我们利用上述纯化的MART1 重组蛋白作为抗原免疫日本大耳白兔,经1 次启动免疫和2 次强化免疫后,在实验动物体内获得高效价的抗MART1 多克隆抗体。经免疫印迹和细胞免疫荧光法分析证实,该抗体能有效识别和结合原核及真核细胞中表达的MART1蛋白,具有良好的特异性。
综上所述,我们建立了小鼠MART1蛋白原核表达和纯化技术,制备出的高效价和特异性抗MART1多克隆抗体将为黑色素瘤的防治研究提供技术支撑。
[1]Weinstein D,Leininger J,Hamby C,et al.Diagnostic andprognostic biomarkers in melanoma[J].J Clin Aesthet Dermatol,2014,7(6):13-24.
[2]Wang X,Gillies E,Hassell L A,et al.Comparative genomic hybridization in a case of melanoma that loses expression of S100,HMB45,Melan A and tyrosinase in metastasis[J].Int J Clin Exp Pathol,2014,7(1):468-473.
[3]Chauvin J M,Larrieu P,Sarrabayrouse G,et al.HLA anchor optimization of the melan-A-HLA-A2 epitope within a long peptide is required for efficient cross-riming of human tumorreactive T cells[J].J Immunol,2012,188(5):2102-2110.
[4]Dadaglio G,Morel S,Bauche C,et al.Recombinant adenylate cyclase toxin of bordetella pertussis induces cytotoxic T lymphocyte responses against HLA*0201-restricted melanoma epitopes[J].Int Immunol,2003,15(12):1423-1430.
[5]Buhl T,Braun A,Forkel S,et al.Internalization routes of cell-penetrating melanoma antigen peptides into human dendritic cells[J].Exp Dermatol,2014,23(1):20-26.
[6]Zelba H,Weide B,Martens A,et al.Circulating CD4+T cells that produce IL4 or IL17 when stimulated by melan-A but not by NY-ESO-1 have negative impacts on survival of patients with stage IV melanoma[J].Clin Cancer Res,2014,20(16):4390-4399.
