Pb、Cd胁迫对紫茎泽兰N、P、K吸收的影响
2015-11-28刘小文齐成媚欧阳灿斌颜冬冬王秋霞郭美霞曹坳程
刘小文, 齐成媚, 欧阳灿斌, 李 园,颜冬冬, 王秋霞, 郭美霞, 曹坳程*
(1. 湘南优势植物资源综合利用湖南省重点实验室, 永州 425199; 2. 湖南科技学院生命科学与化学工程系, 永州 425199; 3.中国农业科学院植物保护研究所, 北京 100193)
Pb、Cd胁迫对紫茎泽兰N、P、K吸收的影响
刘小文1,2,3, 齐成媚2, 欧阳灿斌3, 李 园3,颜冬冬3, 王秋霞3, 郭美霞3, 曹坳程3*
(1. 湘南优势植物资源综合利用湖南省重点实验室, 永州 425199; 2. 湖南科技学院生命科学与化学工程系, 永州 425199; 3.中国农业科学院植物保护研究所, 北京 100193)
采用模拟Pb、Cd污染土壤培养法研究了紫茎泽兰在重金属Pb、Cd胁迫下植株不同部位对3种营养元素(N、P、K)吸收的影响。结果表明,紫茎泽兰各组织在Pb胁迫下对N、P的吸收总体上均保持一定的促进作用,对K则表现出一定的抑制效应;Cd胁迫下对N、P有良好的吸收,低浓度Cd促进K的吸收,高浓度则表现出抑制作用。Pb-Cd复合污染胁迫下根茎叶均对N的吸收有一定的促进作用,而对K的吸收有较强的抑制效应。这表明,紫茎泽兰在重金属Pb、Cd胁迫下可通过大量吸收N、P等元素来有效缓解重金属的毒害作用,这可能也是紫茎泽兰能适应高Pb、Cd胁迫的一种耐性机制。
紫茎泽兰; Pb; Cd; 营养元素
紫茎泽兰(AgeratinaadenophorumSprengel)是一种生长迅速,繁殖能力强、抗逆性强、传播推进快、群体自然演替能力强的植物,极易在裸地、稀疏植被生境和其他恶劣条件下存活和繁殖的世界性恶性杂草[1],被列入《外来有害生物的防治和国际生防公约》中四大恶性杂草之一[2]。作为外来入侵物种,紫茎泽兰在我国西南地区广泛分布、繁殖能力强、传播范围广、生物量大、抗逆性强、生长迅速,已成为恶性杂草并发展到难以控制的程度[3],在国家环保总局首批公布的16种外来入侵物种中,名列首位,是我国面临的重大生态灾害物种之一[4]。紫茎泽兰在新的生境能迅速扩展,这与其在新的环境压力选择下产生的形态、生理的可塑性变化密切相关,使其更能适应和充分利用当地的环境条件,最终在竞争中占据主导地位而得以扩散[5]。
N、P、K是植物必需的营养元素,在体内物质的组成和代谢过程中有重要的作用[6-8],在植物体内蛋白质、核酸等重要物质的合成和代谢过程中都是必不可少的[9]。植物体内N、P、K缺乏或含量较低会导致植物体内物质组成和代谢的紊乱[10]。植物根系吸收离子过程中不同离子之间往往互相影响和制约,重金属的胁迫有时会引起大量营养元素的缺乏,介质中较高浓度的重金属会引起植物对大量营养元素的吸收和转运能力下降[11-12],导致体内缺乏相应的营养元素,进而引起内部生理代谢紊乱,外部呈现相应的缺素症状[13]。同时,重金属胁迫也可能对一些营养元素的吸收有促进作用[14-17]。Pb、Cd是植物生长的非必需元素,两者均可对植物的叶绿素合成和抗氧化酶产生不利影响,当超过一定剂量则严重影响植物的生理代谢活动,阻碍植物生长发育甚至导致植物死亡[18]。然而,笔者前期研究也表明,无论在自然状态下还是盆栽试验均证明紫茎泽兰对Pb、Cd具有较强耐性,盆栽试验表明1 000 mg/kg Pb或者100 mg/kg Cd胁迫下,紫茎泽兰均具有较好的耐受性,且紫茎泽兰对Pb、Cd具有一定的吸收转移能力[19-20]。而有关紫茎泽兰体内Pb、Cd的分布对其他离子的吸收有何影响,目前尚未见文献报道。本文研究了Pb、Cd胁迫下紫茎泽兰体内N、P、K等3种大量元素的积累与分布特征,这对于揭示紫茎泽兰的耐Pb、Cd特性和防控紫茎泽兰都有重要意义。
1 材料与方法
1.1 材料
供试材料为西昌学院校内试验地表层土壤,理化性质:pH 6.30(水)/7.71(CaCl2),总P为441 mg/kg,总N为853 mg/kg,总K为2 313 mg/kg,CEC(阳离子交换量)为11.23 meq/100 g,有机质含量为25.1 mg/kg,重金属Pb、Cd含量分别为20、0.2 mg/kg。
1.2 方法
1.2.1 植株培养方法
试验在温室大棚里进行,白天温度为(25±2)℃,夜间温度为(15±2)℃,相对湿度55%±8%。Pb胁迫处理水平为:CK(对照),200、500、1 000 mg/kg,Pb以Pb(NO3)2的形式加入;Cd胁迫处理水平为:CK(对照),20、50、100 mg/kg,Cd以CdCl2·2.5H2O形式加入;交互处理水平为:500 mg/kg Pb+50 mg/kg Cd,土壤处理后,拌匀,每盆装土壤2 kg,稳定两周后移植紫茎泽兰幼苗。选取重量和高度一致约8~10 cm的植株,随机分配到每个浓度处理中,每个处理栽种16盆,每盆1株。移植后,用称重法补充水分,保持土壤含水量为田间持水量的60%左右,重金属胁迫处理60 d后,收获供试植物和土壤,将根、茎、叶分开,105 ℃下杀青30 min,并在65~70 ℃烘干至恒重,干样经粉碎后经过浓HNO3∶HF∶HClO4(2∶1∶2)消化,用北京瑞利WFX-120A /石墨炉原子吸收分光光度计测定N、P、K的含量,参照隋方功[21]的方法测定。
1.2.2 数据处理与分析
上述试验均重复3次,计算平均值,并采用SAS 8.2软件和Excel进行统计分析和作图。并用Duncan氏检验法对显著性差异(P<0.05)进行多重比较。
2 结果与分析
2.1 Pb胁迫对紫茎泽兰N、P、K吸收的影响
由表1可得,Pb胁迫后紫茎泽兰根累积的N随Pb浓度的增加呈不同程度的变化, 1 000 mg/kg处理组显著高于对照,而200、500 mg/kg处理组却显著低于对照(P<0.05);P的累积随Pb浓度的增加呈现先升后降的趋势,各处理均显著高于对照(P<0.05),以500 mg/kg处理组为最大,增幅达41.08%;K的累积量随Pb浓度的增加也是先升高后降低,200 mg/kg处理组达到最大,为对照的1.4倍,除1 000 mg/kg处理组外,其他处理组均与对照达到显著性差异(P<0.05),说明Pb胁迫后能显著增加紫茎泽兰根中P的累积量,低浓度的Pb能促进根对K的吸收,高浓度的Pb则对N的吸收十分有利。
经Pb胁迫后,紫茎泽兰茎中N的累积量随Pb浓度的增加先升后降,200 mg/kg处理组显著高于对照,其他各处理则明显低于对照,1 000 mg/kg处理时仅为对照的57.52%;P的吸收则随Pb浓度的增加持续增加,1 000 mg/kg处理时茎中P的累积量大大增加,达到对照的1.94倍;K的累积量随Pb浓度的增加显著降低,各处理与对照差异显著(P<0.05),1 000 mg/kg处理组仅为对照的40.49%,此时K的吸收受到了较强的抑制。这表明Pb胁迫后有助于紫茎泽兰茎对P的吸收,低浓度的Pb能促进茎对N的吸收,对K的吸收有一定的排斥作用。
紫茎泽兰叶中N的累积量随Pb胁迫浓度的增加出现显著上升的变化趋势(P<0.05), 1 000 mg/kg处理时比对照增幅超过100%;P的吸收则随Pb浓度的增加出现折线型变化,以200 mg/kg处理时达到最大,500 mg/kg处理时为最小;K的累积量随Pb浓度的增加呈先降后升的变化趋势,各处理均显著低于对照,K的吸收一定程度上受到了抑制。Pb胁迫下紫茎泽兰叶表现出对N的强烈吸收,低浓度的Pb能促进叶对P的吸收,不利于对K的吸收。
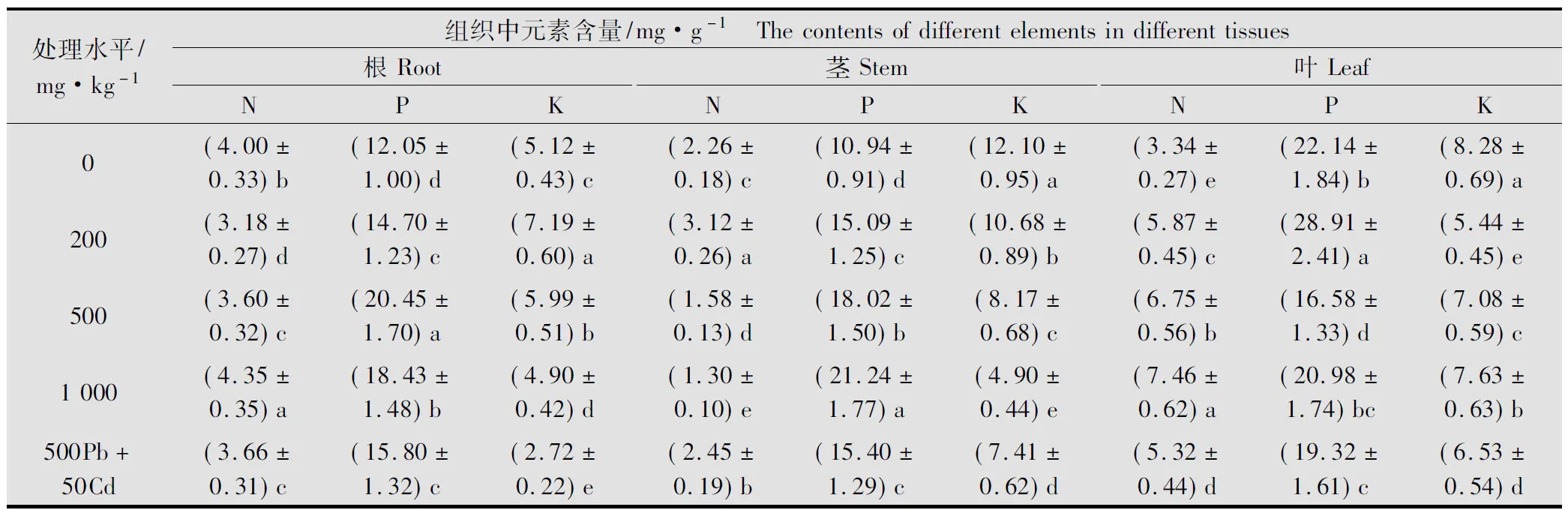
表1 Pb胁迫对紫茎泽兰N、P、K积累的影响1)
1) 表中数据代表平均值±标准差;同列不同小写字母代表差异显著(P<0.05)。下同。
The data in the table represent the mean±SD; different letters in the same column indicate significant difference at 0.05 level; the same below.
2.2 Cd胁迫对紫茎泽兰N、P、K吸收的影响
从表2可以看出,Cd胁迫后紫茎泽兰根累积的N随Cd浓度的增加出现不同程度变化,低浓度处理组显著低于对照(P<0.05),100 mg/kg处理时又显著高于对照,为对照的1.64倍;P的吸收则随Cd浓度的增加出现先上升后下降的变化趋势,各处理均显著高于对照(P<0.05),50 mg/kg处理时根中P的累积量达到最大,为对照的132.9%;K的累积量类似于P,但其以20 mg/kg处理组最大,其他处理组均明显低于对照(P<0.05);表明Cd胁迫后能有效增加紫茎泽兰根中P的累积量,低浓度的Cd有助于根对K的吸收,高浓度的Cd则对N的吸收十分有利。
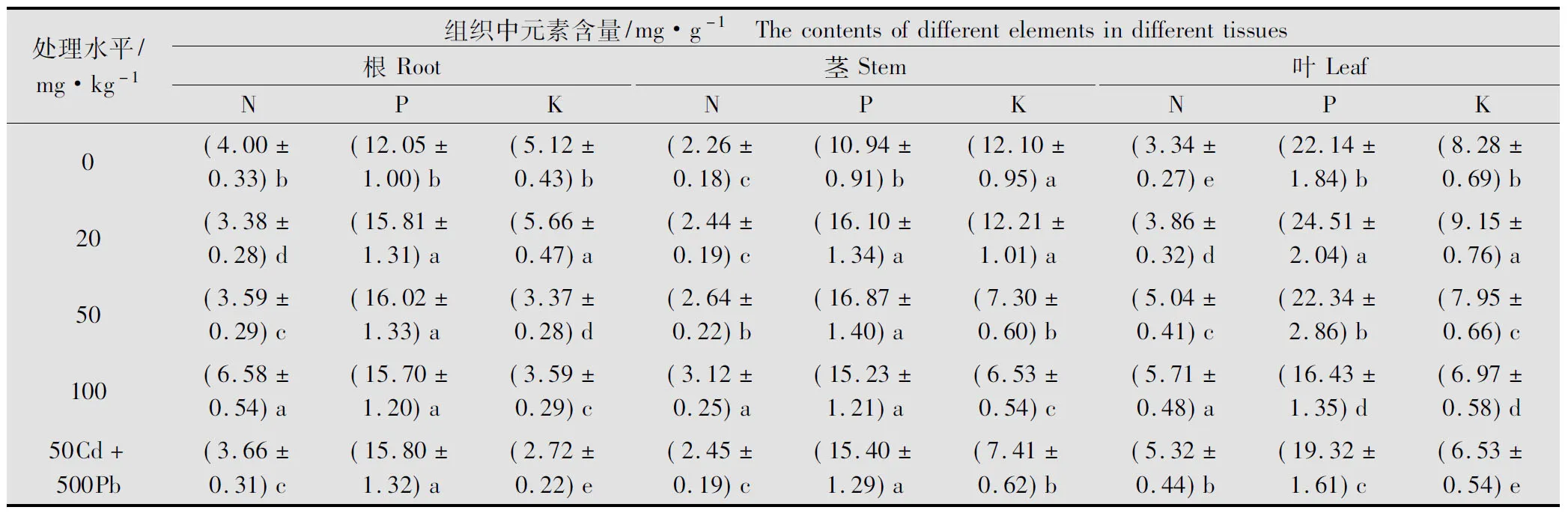
表2 Cd胁迫对紫茎泽兰N、P、K积累的影响
经Cd胁迫后,紫茎泽兰茎中N的累积量随Pb浓度的升高缓慢增加,50、100 mg/kg处理组均显著高于对照(P<0.05),低浓度20 mg/kg处理时与对照差异不显著;P的累积量则随Pb浓度的增加出现先升后降的趋势,但各处理均显著高于对照(P<0.05),50 mg/kg处理组为对照的1.54倍;茎中K的吸收则随Pb浓度的增加出现先缓慢增加后快速下降的趋势,100 mg/kg处理组仅为对照的53.96%;说明Cd胁迫下对N、K的吸收十分有利,低浓度时有助于K的吸收,高浓度时K的累积受到了明显的抑制。
紫茎泽兰叶中N的累积量随Cd胁迫浓度的增加持续上升,各处理与对照差异显著, 100 mg/kg处理时比对照增幅超过70%;P的吸收则随Cd胁迫浓度的增加先上升后缓慢下降,在20 mg/kg处理时达到最大,100 mg/kg处理时为最小,分别为对照的110.70%和74.21%;K的吸收情况类似于P,随Cd浓度的增加呈先升后降的变化趋势。Cd胁迫下紫茎泽兰叶表现出对N的强烈吸收,低浓度的Pb能促进叶对P、K的吸收,高浓度对P、K均有一定程度的抑制作用。
2.3 Pb-Cd复合污染胁迫对紫茎泽兰N、P、K吸收的影响
由表1和表2可得,Pb-Cd复合污染胁迫下紫茎泽兰根N累积量和单一Pb、Cd处理时相当,也明显低于对照;P的累积量明显低于单一Pb处理,与单一Cd处理无明显差异(P<0.05),且高于对照;K的累积量则显著低于对照和单一Pb、Cd处理,说明Pb-Cd复合污染胁迫紫茎泽兰根对K的吸收明显受到抑制,对P的吸收反而有一定的促进效应。紫茎泽兰茎N的累积量明显低于单一Cd处理,高于对照和单一Pb处理;对P的吸收则比单一Pb、Cd处理都有所降低,但却高于对照;K的累积量则Pb-Cd复合处理组高于单一Cd处理,显著低于单一Pb处理和对照。Pb-Cd复合污染胁迫下紫茎泽兰叶对N的吸收明显高于对照和单一Cd处理,与单一Pb相比有所下降;对P的吸收量则是显著低于对照和单一Cd处理,但却高于单一Pb处理;K的吸收则是大大下降,均低于对照和单一Pb、Cd处理。说明Pb-Cd复合污染胁迫下根茎叶对N的吸收有一定的促进作用,对K的吸收有较强的抑制效应。
2.4 紫茎泽兰各组织中Pb、Cd含量与营养元素的相关性分析
由表3可知,在Pb的胁迫下,紫茎泽兰根中Pb的含量与N和P的吸收呈正相关,与K的吸收呈显著性负相关;茎中Pb的含量与N呈显著的负相关,与P的吸收呈显著性正相关,与K则呈显著的负相关;叶中Pb的含量与N、P呈显著的正相关,进一步说明在Pb胁迫下,紫茎泽兰对N、P的吸收具有一定的促进作用,对K的吸收则表现为抑制作用。
Cd胁迫下,紫茎泽兰根中Cd的含量与N和P的吸收呈正相关,与K的吸收呈显著负相关;茎中Cd的含量也与N、P呈显著的正相关,与K的吸收呈显著负相关;叶中Cd的含量与N呈显著的正相关,与P、K的吸收呈显著负相关,表明Cd胁迫能有效促进紫茎泽兰对N的吸收,抑制其对K的吸收,对P吸收的影响在不同组织中并不一致。
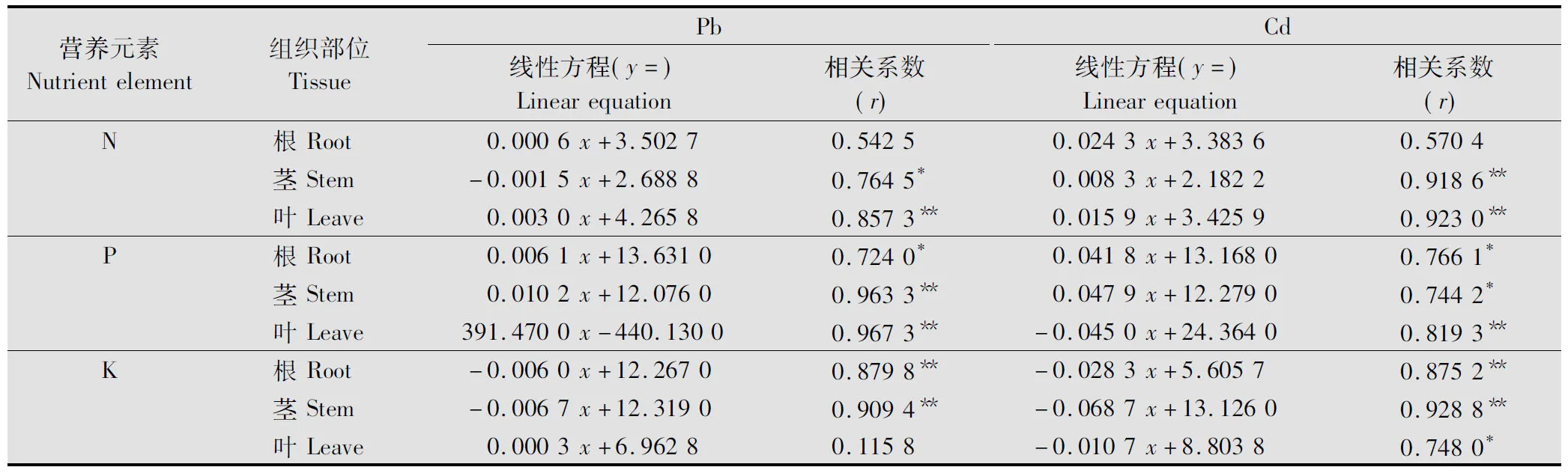
表3 紫茎泽兰各组织中Pb、Cd含量与营养元素的相关性分析1)
1)*相关显著性水平为0.05,**相关显著性水平为0.01。
*indicates significant correlation atP<0.05;**indicates significant correlation atP<0.01.
3 结论与讨论
作为我国危害最严重的外来入侵物种之一,紫茎泽兰能极快适应环境的变化,并具有异株克生现象和惊人的繁殖能力,可逐渐形成密集成片的单种优势群落,导致原有的植物群落衰退和消失[22-24]。目前,我国每年对紫茎泽兰防控投入了大量人力和物力[25],但对紫茎泽兰尤其是Pb、Cd污染严重地区紫茎泽兰的入侵并快速蔓延尚无根治措施,原因在于紫茎泽兰对Pb、Cd等环境因子的耐性机制及其胁迫下体内各营养元素的积累与分布特征尚不明确。
Pb和Cd是植物生长的非必需元素,前期研究表明,在Pb、Cd胁迫下时紫茎泽兰的长势更旺,生物量有所增加。Pb浓度达到1 000 mg/kg或者Cd浓度达到100 mg/kg,紫茎泽兰均能较好地生长,对Pb、Cd有较强的耐受能力,是一种耐Pb、Cd植物,在Pb、Cd胁迫下紫茎泽兰反而有竞争优势,依靠强大的生物量适应Pb、Cd污染环境。Fabian等[26]的研究表明,经Pb和Zn处理的冬小麦幼苗,叶和根的生长明显受到抑制,营养溶液中的其他重金属离子(铬、铜、镉、镍)对其也有不同程度的毒性抑制作用,而氮肥的施用减轻了以上重金属离子产生的毒性抑制作用,且随施氮水平的提高,毒性抑制作用降低;聂俊华[27]研究表明营养元素N和K会促进超富集植物对Pb的吸收;王学[28]的研究表明,外源多胺也可有效维持锌胁迫下植物体内各种营养元素的平衡。本研究发现N、P具有缓冲重金属引起植物毒害的能力,紫茎泽兰可通过自身的调节作用不断从土壤中吸收大量的N、P来消除Pb对其的毒害作用。
据报道外界较高的Cd浓度也引起玉米植株P浓度和吸收量的下降[11];随Cu、Cd浓度的增加,K的外流量增加[12];用Cu和Cd处理30 min的小麦切根在Na2EDTA存在的情况下,抑制了K+吸收,而Cr、Fe、Hg处理的小麦切根则对K+的吸收无抑制作用[29]; Wu等[30]的研究发现,添加外源Cd显著地降低了大麦根中Mn 和Zn 的含量;Zhang等[31]的研究表明,Cd胁迫显著地促进小麦根部P、K 和Mn 含量的增加;Liu等[32]的研究表明,Cd胁迫影响了水稻对Fe、Zn、Mn、Mg 等营养元素的吸收和积累,其影响效果因元素种类而异。本试验结果说明了紫茎泽兰各组织在Cd胁迫下对N、P有良好的吸收,低浓度Cd对K的吸收有一定促进作用,高浓度则表现出抑制作用。说明紫茎泽兰对Cd的耐受能力较强,其通过大量吸收土壤中的N、P来缓解重金属的毒害效应,从而保护其自身的正常生命活动。Pb-Cd复合污染胁迫下根茎叶对N的吸收有一定的促进作用,对K的吸收也有较强的抑制效应。
综上所述,紫茎泽兰能适应体内Pb、Cd的胁迫环境,避免其对生长发育产生不利影响。在Pb、Cd胁迫下,紫茎泽兰各组织通过大量吸收N、P等元素来有效缓解重金属的毒害作用,生命活动并没有受到影响,这可能也是紫茎泽兰能适应高Pb、Cd胁迫的一种耐性机制。因此,开展Pb、Cd胁迫下紫茎泽兰体内N、P、K等3种大量元素的积累与分布特征的研究,有助于寻找更为有效的紫茎泽兰防治手段,从而节约防治成本,减少对生态环境的破坏,对我国尤其是Pb、Cd污染严重地区紫茎泽兰的防治起到积极作用。
[1] 刘士奇. 综合治理外来入侵植物刻不容缓[J]. 福建农业,2004(3): 33-33.
[2] 刘小文,齐成媚,梁剑,等. 重金属铅、镉污染下紫茎泽兰对氨氯吡啶酸的耐性[J]. 植物保护,2013, 39(6): 32-36.
[3] 温三明. 植物“食人鱼”紫茎泽兰的好归宿[J]. 农村实用技术,2005(2): 11.
[4] 潘文龙. 国家环保总局公布的16种有害外来物种简介[J]. 生物学教学,2007, 32(11): 68-70.
[5] 邓雄,杨期和,叶万辉,等. 生物入侵的适应性进化及其影响[J]. 中山大学学报:自然科学版,2003, 42(A19): 204-210.
[6] Khaled H, Fawy H A. Effect of different levels of humic acids on the nutrient content, plant growth, and soil properties under conditions of salinity [J]. Soil and Water Research, 2011, 6(1): 21-29.
[7] Wang Chao, Zhang Songhe. Salicylic acid involved in the regulation of nutrient elements uptake and oxidative stress inVallisnerianatans(Lour.) Hara under Pb stress[J]. Chemosphere, 2011, 84(1): 136-142.
[8] Sawas Dimitrios, Colla Giuseppe, Rouphael Youssef, et al. Amelioration of heavy metal and nutrient stress in fruit vegetables by grafting[J]. Scientia Horticulturae, 2010, 127(2): 156-161.
[9] 潘瑞炽. 植物生理学[M]. 北京:高等教育出版社,2001.
[10]邓兰生,李楠,王海燕,等. 水肥一体化下番木瓜养分吸收累积规律[J]. 热带作物学报,2013,34(8): 1426-1429.
[11]Narwal R P, Singh M, Singh J P, et al. Cadmium-zinc interaction in maize grown on sewer water irrigated soil[J]. Arid Land Research and Management, 1993, 7(2): 125-131.
[12]Gussarsson M, Jensén P. Effects of copper and cadmium on uptake and leakage of K+in birch (Betulapendula) roots [J]. Tree Physiology, 1992, 11(3): 305-313.
[13]安志装,王校常,施卫明,等. 重金属与营养元素交互作用的植物生理效应[J]. 土壤与环境,2002, 11(4): 392-396.
[14]陈京都,何理,许轲,等.镉胁迫对不同基因型水稻生长及矿质营养元素吸收的影响[J].生态学杂志,2013,32(12):3219-3225.
[15]张自坤,刘作新,张颖,等.铜胁迫对嫁接和自根黄瓜幼苗光合作用及营养元素吸收的影响[J].中国生态农业学报,2009,17(1):135-139.
[16]程旺大,姚海根,张国平,等. 镉胁迫对水稻生长和营养代谢的影响[J]. 中国农业科学,2005, 38(3): 528-537.
[17]杨明杰,林咸永,杨肖娥. Cd对不同种类植物生长和养分积累的影响[J]. 应用生态学报,1998, 9(1): 89-94.
[18]Wu F, Zhang G, Dominy P. Four barley genotypes respond differently to cadmium: lipid peroxidation and activities of antioxidant capacity [J]. Environmental and Experimental Botany, 2003, 50(1): 67-78.
[19]汪文云,张朝晖. 贵州水银洞卡林型金矿苔藓植物研究[J]. 黄金,2008, 29(6): 8-12.
[20]李冰,张朝晖. 贵州烂泥沟金矿区苔藓植物及其生态修复潜力分析[J]. 热带亚热带植物学报,2008, 16(6): 511-515.
[21]隋方功. 土壤农化分析实验[M]. 北京:中国农业大学出版社,2004.
[22]胡楚娇,王崇云,和兆荣,等. 基于DPPH法对紫茎泽兰提取物抗氧化活性的研究[J]. 杂草科学,2013, 31(4): 9-12.
[24]黄梅芬,徐驰,曹后英,等.不同生境条件对紫茎泽兰营养生长的影响[J]. 热带作物学报,2009,30(10): 1429-1436.
[25]朱文达,曹坳程,颜冬冬,等.不同林木种群对紫茎泽兰营养生长和生殖生长的影响[J].生态环境学报,2013,22(11):1790-1794.
[26]Fabian G, Dezsi D M. Ecophysiological studies of the relationshipbetween heavy metal toxicity and nitrogen nutrition in the early development stage of winter wheat[J]. Acta Botanica Hungarica, 1987, 33(3/4): 219-234.
[27]聂俊华,刘秀梅,王庆仁. Pb超富集植物对营养元素N、P、K的响应[J]. 生态环境,2004, 13(3): 306-309.
[28]王学. 多胺对锌胁迫下水荷花叶片营养元素吸收的影响[J]. 安徽农业科学,2008, 36(30): 13027-13028.
[29]Bujtas C C E. Inhibition of K+absorption by heavy metal ions[J]. Acta Biochemica et Biophysica Academiae Scientiarum, 1981, 16(3/4): 253.
[30]Wu F B, Zhang G P, Yu J S. Interaction of cadmium and four microelements for uptake and translocation in different barley genotypes [J]. Communications in Soil Science and Plant Analysis, 2003, 34: 2003-2020.
[31]Zhang G P, Fukami M, Sekimoto H. Influence of cadmium on mineral concentrations and yield components in wheat genotypes differing in Cd tolerance at seedling stage [J]. Field Crops Research, 2002, 77: 93-98.
[32]Liu J G, Liang J S, Li K Q, et al. Correlations between cadmium and mineral nutrients in absorption and accumulation in various genotypes of rice under cadmium stress[J]. Chemosphere, 2000, 52: 1467-1473.
(责任编辑:田 喆)
Effects of lead, cadmium on N, P, K uptake ofAgeratinaadenophorum
Liu Xiaowen1,2,3, Qi Chengmei2, Ouyang Canbin3, Li Yuan3, Yan Dongdong3, Wang Qiuxia3, Guo Meixia3, Cao Aocheng3
(1. Key Laboratory of Comprehensive Utilization of Advantaged Plant Resources in Southern Hunan, Yongzhou 425199, China; 2. Department of Life Science & Chemical Engineering, Hunan University of Science and Engineering, Yongzhou 425199, China; 3.Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China)
The effects of Pb, Cd on nutrient elements (N, P, K) uptake in different parts ofAgeratinaadenophorumwere investigated in a soil pot experiment. The results showed that it had a positive effect on the absorption of N, P in different tissues under Pb stress, but negative effect on K uptake. Under the conditions of Cd stress, it was favorable for the absorption of N, P in different tissues, and the absorption of K was promoted at low Cd concentration, but inhibited at high concentration. It also had a positive effect on the absorption of N in different tissues under combined Pb-Cd pollution stress, but negatively affected K uptake. A large quantity of N and P was absorbed to alleviate the toxic effects onA.adenophorumby heavy metals. It is a possible tolerance mechanism ofE.adenophorumunder Pb and Cd stresses.
Ageratinaadenophorum; lead; cadmium; nutrient element
2014-05-28
2014-08-04
公益性行业(农业)科研专项(201103027); 湖南省自然科学基金(13JJ6080);湖南省重点学科建设项目(2011-76);湖南省高校科技创新团队支持计划(2012-318)
S 45
A
10.3969/j.issn.0529-1542.2015.04.015
* 通信作者 E-mail:caoac@vip.sina.com
