基于iTraq技术的加拿大一枝黄花提取物作用下铜绿微囊藻细胞差异表达蛋白
2015-11-19黄莹莹孔海南上海交通大学环境科学与工程学院上海200240上海交通大学公共卫生学院上海200025
黄莹莹,白 羽,王 艳,2*,孔海南(.上海交通大学环境科学与工程学院,上海 200240;2.上海交通大学公共卫生学院,上海 200025)
基于iTraq技术的加拿大一枝黄花提取物作用下铜绿微囊藻细胞差异表达蛋白
黄莹莹1,白 羽1,王 艳1,2*,孔海南1(1.上海交通大学环境科学与工程学院,上海 200240;2.上海交通大学公共卫生学院,上海 200025)
为研究加拿大一枝黄花提取物对藻类生长抑制作用的分子机理,应用iTraq技术结合LC-MS-MS,分析了铜绿微囊藻细胞蛋白质的差异表达.共鉴定出261种差异蛋白(变化倍数≥1.5),其中表达上调的220种,表达下调的41种.GO功能分类显示,加拿大一枝黄花提取物通过影响细胞代谢过程、蛋白质催化和结合功能等方式抑制藻细胞生长.KEGG通路分析表明,通路大类中富集程度位于前3位的均是代谢通路,包括能量代谢、碳水化合物代谢和氨基酸代谢,通路中富集程度最高的是糖酵解/糖异生通路.本研究发现了加拿大一枝黄花提取物对藻细胞生长抑制作用的分子靶点,为从分子层面阐明其抑藻作用提供参考,也为研究植物化感物质的抑藻机理提供了新方法.
iTraq标记;化感作用;差异表达蛋白组;抑藻机理
研究证实加拿大一枝黄花提取物能够有效抑制藻类的生长[1-2],但其作用机理尚不清楚.目前,对植物化感物质抑藻机理的研究多集中在细胞的生理生化过程,虽然分子水平的研究起步较晚,但研究者普遍认为植物化感物质能够影响藻细胞基因和蛋白的合成和表达[3-5].从现有报道看,与植物化感物质抑藻机理相关的分子水平研究多是从基因表达的角度来考虑[6-8],然而,蛋白质是生理功能的执行者,其特有的活动规律几乎无法从基因水平的研究来获知.
蛋白组学和基因组学技术已经在临床医学、分子生物学和免疫学等领域的研究中得到普遍应用,为从生物整体的蛋白和基因的表达及功能的角度阐明生命现象提供了依据.然而,该类技术尚未在植物化感抑藻研究领域广范展开.近年来,蛋白组学技术在原有基础上改进和衍生出许多新的技术,其中同位素标记相对和绝对定量(iTraq)技术[9]是2004年美国AB SCIEX公司研发的一种最新的蛋白质组学定量技术,具有较好的定量效果、较高的重复性和工作通量,还提高了蛋白质组的覆盖率和鉴定的可信度.鉴于以上优点,iTraq技术已经被越来越多的研究者采用,已成功应用于酵母菌株[10]、小鼠肝脏[11]、人体皮肤[12]、子宫内膜组织[13]、肺腺癌细胞膜蛋白[14]等多种样品的蛋白质组的鉴定和定量研究,但目前尚无将iTraq技术应用于植物化感物质的抑藻机理的研究报道.
本文采用iTraq标记结合LC-MS-MS技术,对加拿大一枝黄花提取物作用下的铜绿微囊藻细胞全蛋白质组进行鉴定,并对差异蛋白进行生物信息学分析,为从分子层面阐明加拿大一枝黄花提取物的抑藻机理提供参考.
1 材料与方法
1.1 藻种的培养
铜绿微囊藻(FACHB 942)购自中国科学院水生生物研究所淡水藻种库.采用BG-11培养基在人工气候室内培养,培养条件为:温度(25±1)℃,光照强度3000~4000lx,光暗周期14L:10D,静置培养.
1.2 加拿大一枝黄花提取物的制备
在上海交通大学闵行校区采集加拿大一枝黄花当年新出苗植株,植株生长土壤为潴育水稻土中的沟干泥、黄泥头,土壤有机质含量约为2%~3%,pH值呈中性或微碱性.加拿大一枝黄花提取物的制备采用以下方法[15]:植物叶片在80℃条件下烘干48h;取粉碎后的植物粗颗粒,用95%乙醇在室温下避光提取;乙醇提取液过滤后,50℃下采用旋转蒸发浓缩,得到乙醇浸膏;乙醇浸膏用蒸馏水温浸,搅拌使之悬浮,用石油醚反复萃取去除其中非极性组分,剩余溶液即为含有水溶性抑藻活性成分的提取物,4℃冷藏备用,使用前经0.45μm微孔滤膜过滤.提取物的浓度以水溶性成分的浓度计算(g/L).
1.3 试剂和仪器
细胞裂解液含40mmol/L Tris/HCl pH7.4、8mol/L Urea、0.2mmol/L Na3VO4、1mmol/L EDTA、1mmol/L NaF、0.5mmol/L EGTA、4%CHAPS、20mmol/L DTT,使用前加入1mmol/L PMSF.还原液含6mol/L盐酸胍和100mmol/L NH4HCO3.Solution A含98%超纯水,2%乙腈,0.1%甲酸.iTRAQ®试剂盒由美国AB Sciex公司提供.
液相色谱为1100LC(美国安捷伦科技公司).LC-MS-MS为Dionex ultimate3000Nano LC(美国ThermoFisher Dionex公司)和On-line Nano Electrospray,maxis impact Q-TOF(德国Bruker公司).
1.4 实验方法
1.4.1 实验设置 BG11培养基置于250mL锥形瓶中,接入对数生长期的铜绿微囊藻,添加0.2g/L的加拿大一枝黄花提取物,对照不添加提取物.
1.4.2 全蛋白的制备 通过离心分别收集加拿大一枝黄花提取物作用3d的藻细胞和对照细胞,加入5mL 4℃预冷细胞裂解液,在冰上进行超声破碎.将细胞破碎液在4℃下,15000r/min离心45min,取上清液,加入25mL的乙醇/丙酮/乙酸(50/50/0.1,V/V/V),-20℃沉淀过夜.沉淀后的细胞液在4℃下,15000r/min离心1h,沉淀用丙酮重悬,50℃下离心浓缩.用还原液重溶沉淀,-20℃保存备用.蛋白质浓度采用Bradford法测定.1.4.3 酶解和标记 200μg蛋白用还原液定容至100μL,采用iTRAQ®试剂盒进行酶解和标记.酶解步骤如下:加入4μL Reducing Reagent,60℃水浴1h;加入2μL Cysteine-Blocking Reagent,室温放置10min;将蛋白溶液加入10KD超滤管中,4℃下12000r/min离心20min,弃掉收集管底部溶液;加入100μL Dissolution Buffer,4℃下12000r/ min离心20min,弃掉收集管底部溶液;更换新的收集管,在超滤管中加入50μL(80μg/mL)胰蛋白酶液,37℃反应过夜.标记步骤如下:超滤管在4℃下12000r/min离心20min;在超滤管中加入50μL Dissolution Buffer,4℃下12000r/min离心20min,重复1次;合并收集管底部滤液,用Dissolution Buffer定容至100μL;iTraq试剂平衡至室温,每管中加入150μL乙醇;取100μg酶解产物,加入全部iTraq试剂,室温反应2~3h;加入100μL超纯水终止反应;分别取加拿大一枝黄花提取物作用的藻细胞和对照细胞的标记后的酶解产物,等体积混合,50℃离心浓缩,-20℃保存备用.
1.4.4 多肽混合物高pH反相分级 将标记样品用120μL流动相A复溶,12000r/min离心20min,通过液相色谱分离样品.色谱柱为Durashell-C18柱(4.6mm×250mm,5μm),柱温为20℃;进样量为100μL;流动相A为pH10的20mmol/L甲酸胺,流动相B为pH10的20mmol/L甲酸胺和80%的乙腈;流速为0.8mL/min;检测波长为214nm.采用梯度洗脱:0~5min,5%流动相B;5~30min,5%~15%流动相B;30~45min,15%~38%流动相B;45~46min,38%~90%流动相B;46~54.5min,90%流动相B;54.5~55min,90%~5%流动相B;55~65min,5%流动相B.从8min开始收集洗脱物,每个样品约1mL,60min结束收集,共收集42份洗脱物.洗脱物分别在50℃离心浓缩,得到样品粉末,-20℃保存备用.
1.4.5 蛋白质分析 分离所得的样品分别用30μL Solution A复溶.依据样品收集的时间,将样品合并为6份,通过LC-MS-MS进行蛋白质检测.色谱条件:捕集柱(trap column)为C18柱(0.1mm×2cm,5μm),流速为10μL/min;分析柱(Analytical Column)为C18柱(0.075mm×0.15cm,3µm),流速为400nL/min;柱温为50℃;流动相A为含0.1%甲酸的超纯水,流动相B为0.1%甲酸的乙腈.采用梯度洗脱:0~2min,2%~8%流动相B;2~75min,8%~20%流动相B;75~93min,20%~35%流动相B;93~98min,35%~80%流动相B;98~118min,80%流动相B.质谱检测条件:电离模式为正离子nanospray模式;质谱检测方式为MS全扫描及MS/MS扫描,扫描范围50~2500amu;母离子设为350~1500m/z.
蛋白鉴定采用Mascot 2.4软件鉴定多肽分子,肽段质量误差(Peptide mass tolerance)为20ppm,串联质谱误差(MS/MS tolerance)为0.05Da,允许最大的胰酶漏切位点数(Max missed cleavages)为2个,NCBInt数据库选用Microcystis aeruginosa.fasta.Mascot结果采用Scaffold 4软件过滤,蛋白阳性结果错误率(Protein FDR)≤2%,肽段阳性结果错误率(Peptide FDR)≤1%.得到表达差异量的结果,变化倍数(Fold Change Ratio)1.5倍以上,认为二者之间差异有统计学意义.
1.4.6 生物信息学分析 采用blast2go 2.7.0将变化倍数1.5倍以上的差异蛋白和参考菌株M.aeruginosa NIES-843的全蛋白质分别与NCBInt数据库比对,对应蛋白分别在基因本体(Gene Ontology,GO)3个分支——生物过程(Biological Process),分子功能(Molecular Function)和细胞组件(Cellular Component)——进行功能上的分类统计.
从KEGG网站下载了M.aeruginosa NIES-843的蛋白序列和其它相关信息作为KEGG分析的参照数据集,将差异蛋白序列和参照数据集进行序列相似性比对,采用相似度≥30%且evalue≤1×10-10进行双向最佳匹配(Bi-direction Best Hit,BBH).通过参照集的ID号映射到KEGG Pathway maps,以参照数据集的全部蛋白质为参照在通路水平和通路大类的水平进行计算P值,与此同时计算Z-score.Z-score>0且P<0.05,即显著富集.
通过BBH结果将匹配到的差异蛋白质定位到M.aeruginosa NIES-843基因组上,获得编码基因的相关信息.
2 结果与讨论
2.1 蛋白鉴定和差异蛋白筛选
以未经加拿大一枝黄花提取物作用的藻细胞为对照,共鉴定出576种差异蛋白,而差异倍数达到1.5倍的差异蛋白共261种,占到所鉴定的蛋白总数的45.3%,其中表达上调的差异蛋白220种,表达下调的蛋白41种,上调蛋白数是下调蛋白数的5.4倍.表1显示了上调和下调前10位的蛋白.
2.2 GO功能分类和显著性分析
基因本体(GO)是一个在生物信息学中广泛使用的本体,基因产物在数据库中被赋上GO的词条,进而可以到数据库中去查询这些生物学的相关信息.通过GO功能分类对蛋白功能进行聚类分析,第二层GO功能分类结果见表2.鉴定到的差异蛋白中193种能对应到生物过程,其第二层功能分类中180种蛋白参与代谢过程,占93%,第三层分支中,最多蛋白参与的是有机物代谢过程和初级代谢过程,均有109种蛋白,其次是生物合成过程(80种)和细胞代谢过程(78种).鉴定到的差异蛋白中191种能对应到分子功能,其第二层功能分类以催化和结合为主,分别有160和134种相关蛋白.催化功能的下层分支中,水解酶和转移酶最多,分别为45和38种蛋白;结合功能的下层分支中,具有与杂环化合物和有机环状化合物结合功能的蛋白最多,均有80种蛋白,其次是具有与小分子结合功能的蛋白(71种).鉴定到的差异蛋白中81种能对应到细胞组件.第二层GO分支中,这81种蛋白均属于细胞(cell),其中的39种蛋白还同时属于大分子复合物.第三层GO功能细胞分支中,有75种蛋白属于细胞部分(cell part);第三层GO功能大分子复合物分支中,有30种蛋白属于蛋白质复合物.这说明加拿大一枝黄花提取物作用下藻细胞的代谢过程显著变化,蛋白的催化功能和结合功能受到明显影响,而差异蛋白主要位于胞内.
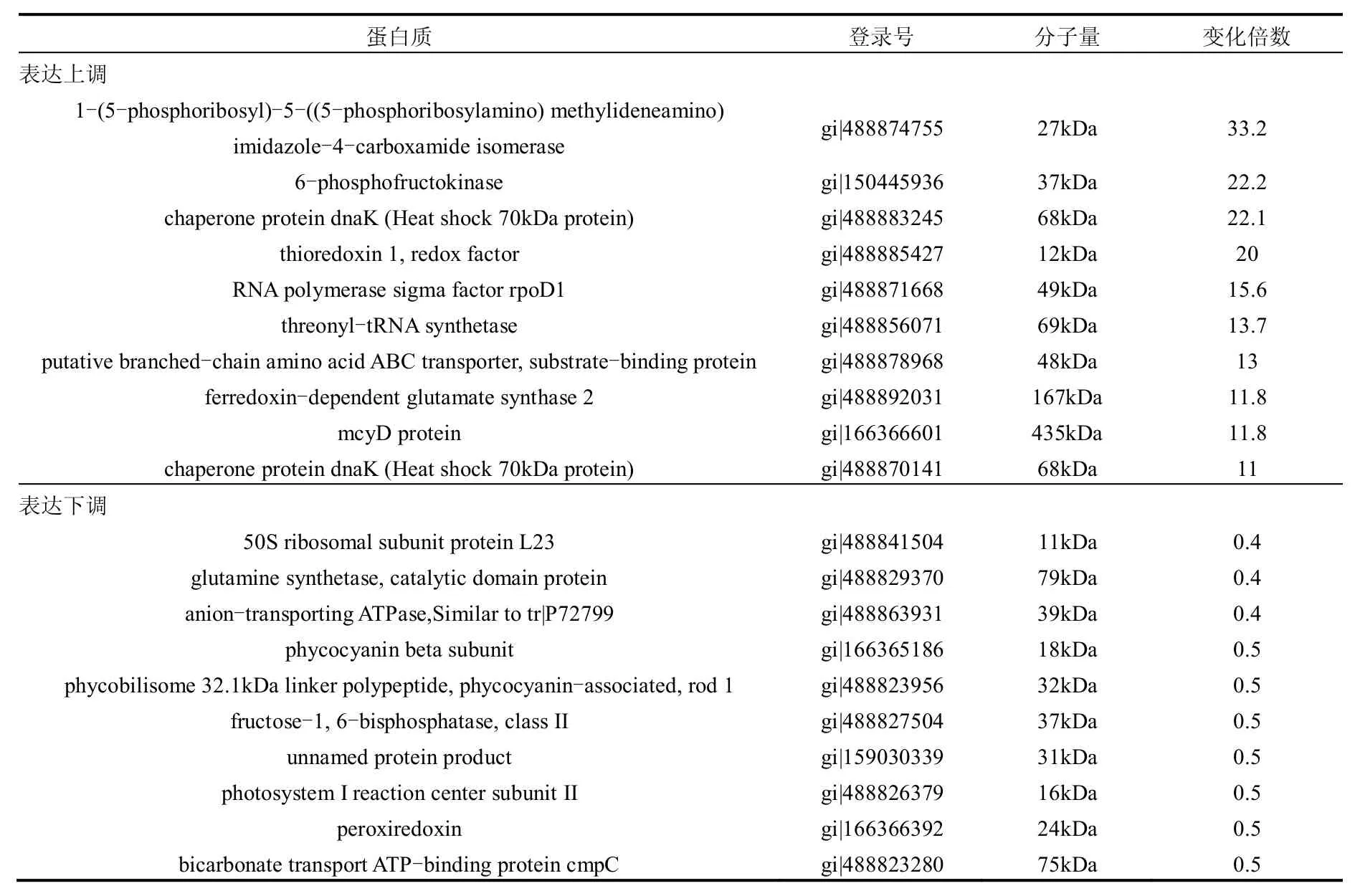
表1 铜绿微囊藻差异蛋白中差异倍数前10位的蛋白Table 1 Top 10 fold change ratios in differentially expressed proteins of M.aeruginosa
以M.aeruginosa NIES-843的全蛋白为参照,采用fisher’s exact test检验实验细胞在GO功能上的差异,得到的P值通过Bonferroni correction进行矫正.表3显示了富集度最高的5个功能分支.相比参考物种,代谢过程、小分子结合和细胞内组件在分别生物过程、分子功能和细胞组件功能分支中差异蛋白富集程度最高.在GO功能分类中,能对应到生物过程的差异蛋白中超过90%与代谢过程相关,下层分支中分解代谢过程、初级代谢过程、有机物代谢过程、生物合成过程和细胞代谢过程显著富集,与这些过程相关的蛋白质代谢、碳水化合物代谢、细胞生物合成、有机质生物合成和细胞大分子生物合成等过程同样显著富集.同样显著富集还有抗氧化活性,其对应的差异蛋白占2.30%(6种),远高于参照物种的0.19%,包括1种过氧化物酶、2种过氧化氢酶和3种谷胱甘肽还原酶,与前期研究中抗氧化系统对加拿大一枝黄花提取物的响应的结论一致[1].
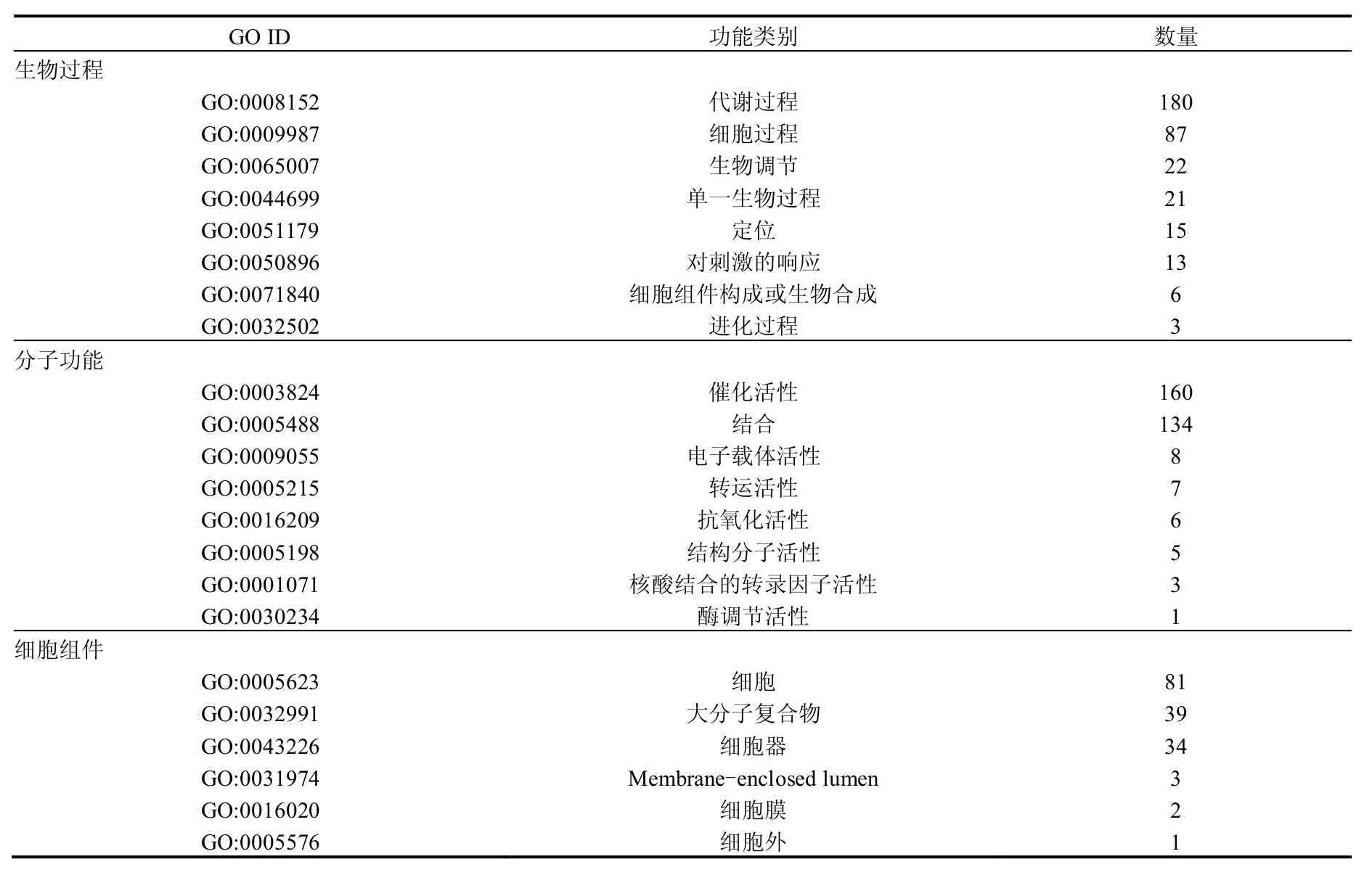
表2 铜绿微囊藻细胞差异蛋白的第二层GO功能分类Table 2 Level 2 of GO terms in differentially expressed proteins of M.aeruginosa
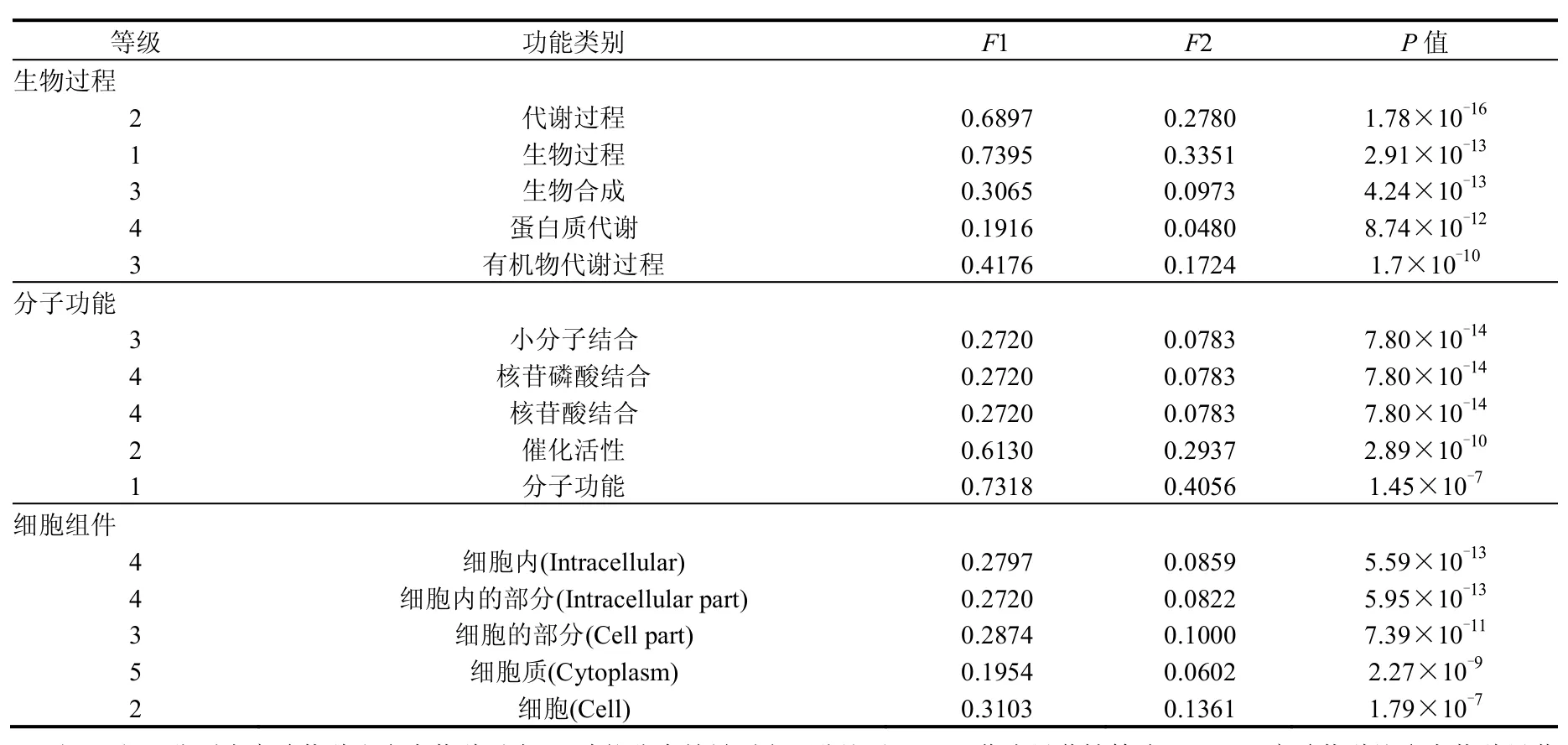
表3 GO功能分类中生物过程、分子功能和细胞组件前5位富集度的功能分类Table 3 Top 5 enrichment GO terms in biological process,molecular function and cellular component
2.3 KEGG通路分析
KEGG是由京都大学生物信息学中心于1995年建立的生物信息学数据库,其中的Pathway数据库可以计算或预测出蛋白质在对各种细胞活动的交互作用,包括图解的细胞生化过程和同系保守的子通路等信息.BBH结果显示有183种(70.8%)蛋白对应到参照数据集,共对应到60条KEGG通路,这些通路分别对应15个通路大类,表4显示了富集程度位于前5位的通路大类,其中位于前3位的均是执行代谢功能的通路大类,包括能量代谢、碳水化合物代谢和氨基酸代谢.60条通路中的22条显著富集,表5显示了富集程度位于前10位的通路,其中糖酵解/糖异生通路富集度最高,其次是氨酰-tRNA酶生物合成合成通路、氮代谢通路和缬氨酸、亮氨酸和异亮氨酸生物合成通路.结果进一步说明加拿大一枝黄花提取物对藻细胞代谢过程的影响最显著的是能量代谢.差异蛋白对应到的15各通路大类中有10个属于代谢过程(4个显著富集),这与GO功能的结果类似.研究证实光合系统是植物化感抑藻物质影响藻细胞生长的重要靶点[16-18],结果显示在对应KEGG通路中有3个与细胞光合生理相关,包括光合作用、光合作用-天线蛋白和碳固定,分别涉及6、5和5种蛋白.

表4 铜绿微囊藻细胞差异蛋白前5位显著富集的通路大类Table 4 Top 5 enrichment KEGG pathway classes in differentially expressed proteins of M.aeruginosa
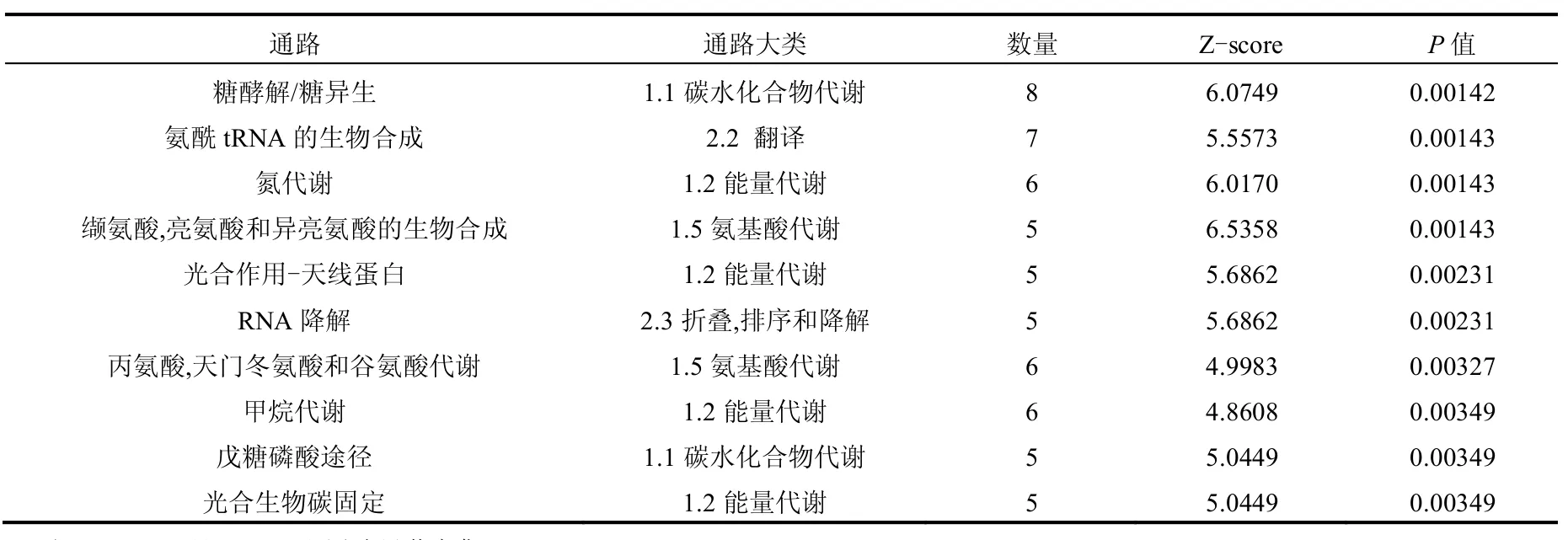
表5 铜绿微囊藻细胞差异蛋白前10位显著富集的通路Table 5 Top 10 enrichment KEGG pathways in differentially expressed proteins of M.aeruginosa
2.4 基因组定位
将BBH得到的183种差异蛋白质分别定位到M.aeruginosa NIES-843基因组上,得到各蛋白对应编码基因的相关信息.表6为上调和下调前5位的蛋白对应的编码基因.
2.5 差异蛋白的生物信息学分析
根据生物信息学分析结果,加拿大一枝黄花提取物对藻细胞的作用是多方面的,其能够同时影响多种物质代谢合成及干扰不同的生理生化反应的进行,通过这些信息还能够进一步解析差异蛋白的功能.
表达上调的蛋白中,1-(5-phosphoribosyl)-5-((5-phosphoribosylamino)methylideneamino)imidazole-4-carboxamide isomerase的差异最大,实验组为对照组的32.3倍.根据生物信息学分析,该酶由hisA基因编码,参与生物合成,具有催化功能,在KEGG通路中参与组氨酸代谢.上调差异倍数第二的是6-磷酸果糖激酶(6-phosphofructokinase),差异倍数为22.2.该酶是糖酵解途径的限速酶,负责催化ATP上的磷酰基转移给6-磷酸果糖,生成1,6-二磷酸果糖和ADP,但会受到高浓度ATP的抑制[19].根据生物信息学分析,该酶编码基因可定位于M.aeruginosa NIES-843基因组上WP_002770220.1位置,与细胞内多种物质(包括核酸碱基、杂环化合物、含氮化合物、芳香族化合物等)的代谢过程相关,具有催化和结合功能,在KEGG通路中参与糖酵解/糖异生、果糖和甘露糖代谢、半乳糖代谢、甲烷代谢.上调差异倍数第三的是热休克蛋白(Heat shock protein),差异倍数为22.1.热休克蛋白是生物体或细胞在不良环境因素作用下产生的具有高度保守性的应激蛋白,不仅能为热损伤所诱导,而且能为许多其它应激刺激所诱导,包括生理、病理、环境乃至机械刺激等方面,如缺氧、葡萄糖缺乏、重金属、代谢抑制因子、氧自由基、病毒感染、寄生虫感染[20].研究证实加拿大一枝黄花提取物作用下铜绿微囊藻会产生过量的活性氧,进而形成氧胁迫[1],这可能是导致热休克蛋白上调的原因.根据生物信息学分析,该蛋白由dnaK基因编码,与细胞内蛋白质和大分子的代谢过程相关,参与应激反应,具有与二磷酸核苷和核苷酸结合的功能,在KEGG通路中参与RNA降解.
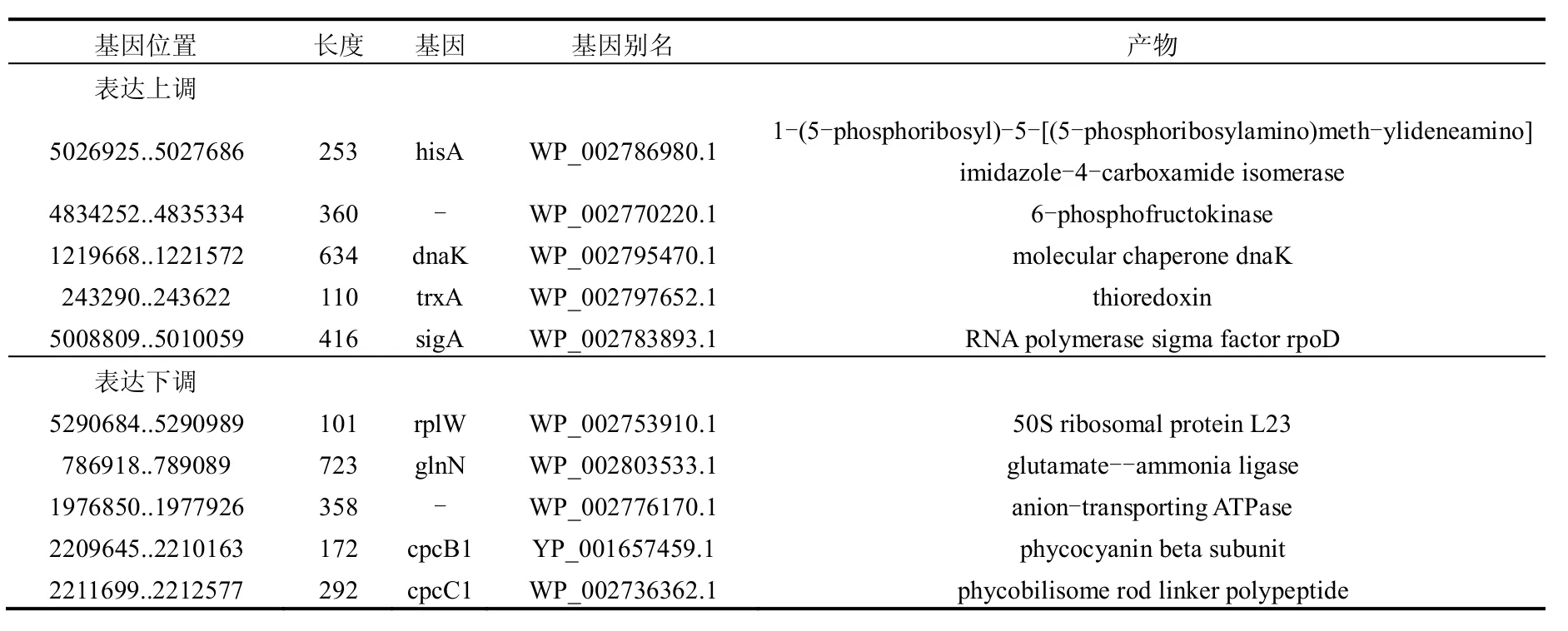
表6 铜绿微囊藻差异蛋白中差异倍数前5位的蛋白对应的编码基因Table 6 Gene of top 5 fold change ratios in differentially expressed proteins of M.aeruginosa
差异蛋白中差异倍数最小的为0.4,即表达下调最显著,包括3种蛋白:50S核糖体亚基蛋白L23(50S ribosomal subunit protein L23)、谷氨酰胺合成酶(glutamine synthetase)以及阴离子转运ATP酶(Anion-transporting ATPase,Similar to tr|P72799).50S核糖体亚基是原核细胞内70S核糖体中的较大亚基,由5S rRNA、23S rRNA和约35种核糖体蛋白质构成,在原核翻译中负责在tRNA转运来的氨基酸分子之间形成肽键,50S核糖体亚基还是某些抗生素的结合位点,这些抗生素可通过阻断肽链的延伸来抑制蛋白的质生物合成,最终杀灭细菌[21-22].根据生物信息学分析,50S核糖体亚基蛋白L23由rplW基因编码,与细胞内物质代谢、合成过程和基因表达翻译相关,具有与RNA结合的功能,在KEGG通路中参与核糖体翻译.谷氨酰胺合成酶是一种参与将无机氮同化为有机氮的酶,可以催化铵离子与谷氨酸反应合成谷氨酰胺,同时消耗ATP[23].该酶由glnN基因编码,与细胞内生物合成和碳水化合物代谢相关,具有催化功能,在KEGG通路中参与丙氨酸、天门冬氨酸和谷氨酸代谢、精氨酸和脯氨酸代谢、乙醛酸和二羧酸代谢、氮代谢和双组分系统.离子转运ATP酶是广泛存在于生物膜上的一种酶蛋白,参与离子的主动转运.阴离子转运ATP酶编码基因可定位于M.aeruginosa NIES-843基因组上WP_002776170.1位置,与细胞内多种物质(包括核酸碱基、杂环化合物、含氮化合物、芳香族化合物等)的代谢过程相关,具有与二磷酸核苷和核苷酸结合的功能以及转移含磷基团的活性,但在KEGG通路没有找到对应通路.
3 结论
3.1 通过对加拿大一枝黄花提取物作用下的铜绿微囊藻细胞蛋白质组进行分离和鉴定,比较对照细胞和实验细胞,共鉴定出576种差异蛋白,其中差异倍数达到1.5倍的差异蛋白共261种,包括220种表达上调的差异蛋白和41种表达下调的差异蛋白.
3.2 通过GO功能分类对蛋白功能分别进行3个分支的聚类分析.鉴定到的差异蛋白中193种能对应到生物过程,其中180种蛋白参与代谢过程,占93%;鉴定到的差异蛋白中191种能对应到分子功能,其中以催化和结合功能为主,分别有160和134种相关蛋白;鉴定到的差异蛋白中81种能对应到细胞组件,第二层分支中这81种均属于细胞,其中的39种蛋白还同时属于大分子复合物.
3.3 差异蛋白中有183种能对应到参照数据集,对应15个通路大类,能匹配到60条KEGG通路中,其中22条显著富集,位于前3位的均是执行代谢功能的通路.
3.4 加拿大一枝黄花提取物于铜绿微囊藻细胞时,能够同时影响多种物质代谢合成途径及干扰不同的生理生化反应,其中受到影响最大的是细胞的代谢功能,尤其是能量代谢和氨基酸代谢过程.
[1]Huang Y,Bai Y,Wang Y,et al.Allelopathic effects of the extracts from an invasive species Solidago canadensis L.on Microcystis aeruginosa[J].Letters in Applied Microbiology,2013,57(5):451-458.
[2]Huang Y,Bai Y,Wang Y,et al.Solidago canadensis L.extracts to control algal(Microcystis)blooms in ponds[J].Ecological Engineering,2014,70:263-267.
[3]张 彬,郭劲松,方 芳,等.植物化感抑藻作用机理[J].生态学杂志,2010,29(9):1846-1851.
[4]张庭廷,郑春燕,何 梅,等.亚油酸对铜绿微囊藻的抑制机理[J].中国环境科学,2009,29(4):419-424.
[5]Shao J,Li R,Lepo J E,et al.Potential for control of harmful cyanobacterial blooms using biologically derived substances:Problems and prospects[J].Journal of Environmental Management,2013,125:149-155.
[6]Zhang C,Yi Y L,Hao K,et al.Algicidal activity of salvia miltiorrhiza bung on microcystis aeruginosa-towards identification of algicidal substance and determination of inhibition mechanism[J].Chemosphere,2013,93(6):997-1004.
[7]Shao J,Wu Z,Yu G,et al.Allelopathic mechanism of pyrogallol to Microcystis aeruginosa PCC7806(Cyanobacteria): From views of gene expression and antioxidant system[J].Chemosphere,2009,75(7):924-928.
[8]Shao J,Yu G,Wang Z,et al.Towards clarification of the inhibitory mechanism of wheat bran leachate on microcystis aeruginosa NIES-843(cyanobacteria): Physiological responses[J].Ecotoxicology,2010,19(8):1634-1641.
[9]Elliott M H,Smith D S,Parker C E,et al.Current trends in quantitative proteomics[J].Journal of Mass Spectrometry,2009,44(12):1637-1660.
[10]Ross P L,Huang Y N,Marchese J N,et al.Multiplexed protein quantitation in Saccharomyces cerevisiae using amine-reactive isobaric tagging reagents[J].Molecular and Cellular Proteomics,2004,3(12):1154-1169.
[11]Walsh J,Jenkins R E,Wong M,et al.Identification and quantification of the basal and inducible Nrf2-dependent proteomes in mouse liver: Biochemical,pharmacological and toxicological implications[J].Journal of Proteomics,2014,108:171-187.
[12]Parkinson E,Skipp P,Aleksic M,et al.Proteomic analysis of the human skin proteome after In Vivo treatment with sodium dodecyl sulphate[J].PLoS ONE,2014,9(5):
[13]DeSouza L,Diehl G,Rodrigues M J,et al.Search for cancer markers from endometrial tissues using differentially labeled tags iTRAQ and cICAT with multidimensional liquid chromatography and tandem mass spectrometry[J].Journal of Proteome Research,2005,4(2):377-386.
[14]Lin H C,Zhang F L,Geng Q,et al.Screening for membrane proteins differentially expressed between the lung adenocarcinoma cell lines with high- and low-metastatic potential using iTRAQ technology[J].Tumor,2013,33(2):150-156.
[15]刘 湘,汪秋安.天然产物化学[M].北京:化学工业出版社,2010.
[16]陈卫民,张清敏,戴树桂.苦草与铜绿微囊藻的相互化感作用[J].中国环境科学,2009,29(2):147-151.
[17]Körner S,Nicklisch A.Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes[J].Journal of Phycology,2002,38(5):862-871.
[18]Zhu J,Liu B,Wang J,et al.Study on the mechanism of allelopathic influence on cyanobacteria and chlorophytes by submerged macrophyte(Myriophyllum spicatum)and its secretion[J].Aquatic Toxicology,2010,98(2):196-203.
[19]Rouwendal G J A,Zwiers L H,Wolbert E J H,et al.Cloning,sequence and expression in Escherichia coli of the gene encoding phosphofructokinase from Bacillus macquariensis[J].Gene,1996,171(1):59-63.
[20]De Maio A.Heat shock proteins: Facts,thoughts,and dreams[J].Shock,1999,11(1):1-12.
[21]Schlunzen F,Zarivach R,Harms J,et al.Structural basis for the interaction of antibiotics with the peptidyl transferase centre in eubacteria[J].Nature,2001,413(6858):814-821.
[22]刘望夷.细菌核糖体的结构和功能[J].生命科学,2009,21(6):771-780.
[23]陈胜勇,李观康,汪 云,等.谷氨酰胺合成酶的研究进展[J].中国农学通报,2010,26(22):45-49.
Differentially expressed proteins in Microcystic aeruginosa with Solidago canadensis L.extracts using iTraq labeling technique.
HUANG Ying-ying1,BAI Yu1,WANG Yan1,2*,KONG Hai-nan1(1.School of Environmental Science and Engineering,Shanghai Jiaotong University,Shanghai 200240,China;2.School of Public Health,Shanghai Jiaotong University,Shanghai 200025,China).China Environmental Science,2015,35(6):1822~1830
To investigate antialgal mechanism of Solidago canadensis L.extracts,iTraq labeling technique coupled with LC-MS-MS was used to analyze differentially expressed proteins in Microcystic aeruginosa.261 differentially expressed proteins(fold change ratio≥1.5)were identified,in which 220 proteins were upregulated and 41proteins were down-regulated.GO function analysis showed that the extracts of S.canadensis inhibited the algal growth by affecting metabolism process,protein catalytic activity and binding function.KEGG pathway analysis showed that top 3 enrichment pathway classes were all related to metabolism,including energy,carbohydrate and amino acid metabolism,and glycolysis/gluconeogenesis was the top enrichment pathway.This study elucidated the molecular targets of the extracts of S.canadensis,thereby giving insight into the antialgal effects at the molecular level and providing a new way to study the antialgal mechanism of plant allelopathic compounds.
iTraq labeling;allelopathy;differentially expressed protein;antialgal mechanism
X172
A
1000-6923(2015)06-1822-09
黄莹莹(1984-),女,广西柳州人,上海交通大学博士研究生,主要从事湖库生态及藻类控制技术研究.发表论文2篇.
2014-10-30
国家自然科学基金(21077074)
* 责任作者,研究员,wangyan66@sjtu.edu.cn
