湖光岩玛珥湖浮游植物、浮游细菌生物量与可培养细菌数量的周年变化
2015-11-01贺艳艳张瑜斌
贺艳艳,李 琴,张瑜斌
(广东海洋大学海洋资源与环境监测中心,广东 湛江 524088)
湖光岩玛珥湖浮游植物、浮游细菌生物量与可培养细菌数量的周年变化
贺艳艳,李琴,张瑜斌
(广东海洋大学海洋资源与环境监测中心,广东湛江524088)
采用丙酮萃取分光光度法、荧光显微计数法、平板计数法分别研究湖光岩玛珥湖5个站位表层水体浮游植物生物量、浮游细菌生物量、可培养细菌数量的周年变化。结果表明,湖光岩玛珥湖浮游植物生物量(以碳C计)和浮游细菌生物量(以C计)、可培养细菌数量周年波动范围分别为84.17~243.64 mg/m3、20.53~ 43.60 mg/m3、0.10×103~ 3.63×103mL-1。三者的周年变化趋势呈规律性同步增减,峰值均出现在3月份,且在5个采样站位间差异无统计学意义(P > 0.05)。与主要生态因子的相关分析显示,浮游植物与浮游细菌生物量均与总氮呈显著正相关(P < 0.05),而可培养细菌数量与氨氮呈显著正相关(P < 0.05),与溶解性氮和磷酸盐呈显著负相关(P < 0.05),但三者与多数调查的生态因子之间相关性无统计学意义(P > 0.05)。浮游植物的初级生产可能是调控细菌周年变化潜在的主导因素。
湖光岩玛珥湖; 浮游植物; 浮游细菌; 可培养细菌; 周年变化
微生物在生态系统中充当分解者[1]、生产者和能量储存库[2],浮游植物是水域生态系统浮游食物网的重要食物源[2],因此,研究微生物与浮游植物对于系统掌握水生生态系统动态有重要意义。玛珥湖是地质年代过程中火山喷发活动停止后形成的封闭锅底形湖泊,位于广东湛江市的湖光岩玛珥湖是世界上最大的深水玛珥湖,在形成和保存高分辨率气候环境记录方面具有独特的优势[3]。目前国内外对玛珥湖的研究主要集中在纹层年代学[4-5]、地球化学[6]、孢粉学[7]、磁性地层学[8-9]、沉积学[10]和藻类学[11]等方面,并取得一些重要成果。而有关玛珥湖的生态学研究虽已有细菌群落结构与功能[12-13]及其多样性[14],浮游植物群落结构及其周年变化的研究[15],但对浮游细菌、浮游植物生物量的周年变化尚未见报道。笔者研究湖光岩玛珥湖的浮游植物生物量、浮游细菌生物量和可培养细菌数量,并结合主要生态因子的周年调查,初步探究各指标的周年动态及其与主要生态因子之间的相互关系,以期为玛珥湖生态系统研究提供科学依据。
1 材料与方法
1.1研究湖泊概况
湖光岩玛珥湖位于中国广东省湛江市(21°8′52.8″ N,110°16′55.2″ E)(图1),是世界上保存完好的两大玛珥湖之一,为一座近似心形的封闭型火山湖,湖水主要来源于大气降水,既无流入也无流出。湖面积约2.1 km2,水面标高23 m,水深最大21 m,平均12 m[3]。
1.2样品的采集
根据湖光岩玛珥湖自身特征和周边环境情况,选取 5个位点作为研究采样站位,分别标记为S1、S2、S3、S4和S5(图1)。根据《水质分析实用手册》[16],使用HQM-1简易采水器采集表层0.5 m处水样,分别装于聚乙烯采样瓶(水样约1 L)以及无菌玻璃瓶(两瓶,水样约100 mL)中。立即在聚乙烯采样瓶中加入1 mL 10g•L-1的饱和碳酸镁悬浮液固定,用于测定叶绿素 a含量;一无菌玻璃瓶中加入经 0.22 μm 微孔滤膜过滤的无颗粒甲醛固定,保持其最终体积分数为 3% ~5%,用于细菌丰度测定,另一瓶直接用于可培养细菌数量测定。采样器具均作无菌处理,采集好的样品装入放有冰块的泡沫冷藏箱中,立即送回实验室分析。采样时间为2008年3月至2009年2月,每月定期采样。
1.3样品分析
浮游植物叶绿素 a含量可表征浮游植物生物量。叶绿素a含量测定采用丙酮萃取分光光度法[16]。浮游植物生物量与叶绿素 a的转换关系按照1个单位质量的叶绿素a相当于50个单位质量的碳含量来换算[17],单位为mg/m3。
浮游细菌丰度采用吖啶橙染色荧光显微计数法[18]测定,水样中加入适量染液后,将其滴入已放置聚碳酸酯微孔滤膜的玻璃滤器砂芯上,抽滤后滤膜转移至载玻片,在荧光显微镜(Olympus Bx51)下观察计数并计算细菌丰度[18]。浮游细菌生物量(以C计)以每个细胞20 fg C为转换系数,将细菌丰度转换为以碳质量浓度表示的细菌生物量[18],单位为mg/m3。
可培养细菌数量采用牛肉膏蛋白胨琼脂培养基的平板计数法[16],水样用无菌水稀释,取适当的3个浓度梯度涂布平板,每个梯度重复3个平板。接种后的平板于28℃下培养36 h后计数,计算每毫升水样菌落形成单位数,用mL-1表示。
1.4主要生态因子测定
为探讨浮游植物和细菌生物量、可培养细菌数量与主要生态因子的关系,在分析叶绿素 a含量和细菌丰度的同时,测定主要相关生态因子。根据《水质分析实用手册》[16],水温和 pH分别用温度计和pH计现场测定,其他指标按规范分析测定,重铬酸钾法测定化学需氧量(COD),磷钼蓝分光光度法测定磷酸盐(PO43--P),过硫酸钾氧化分光光度法测定总磷(TP)、溶解态磷(DP)、总氮(TN)、溶解性氮(DN),靛酚蓝分光光度法测定氨氮(NH4+-N),锌镉还原比色法测硝酸盐氮(NO3--N)。
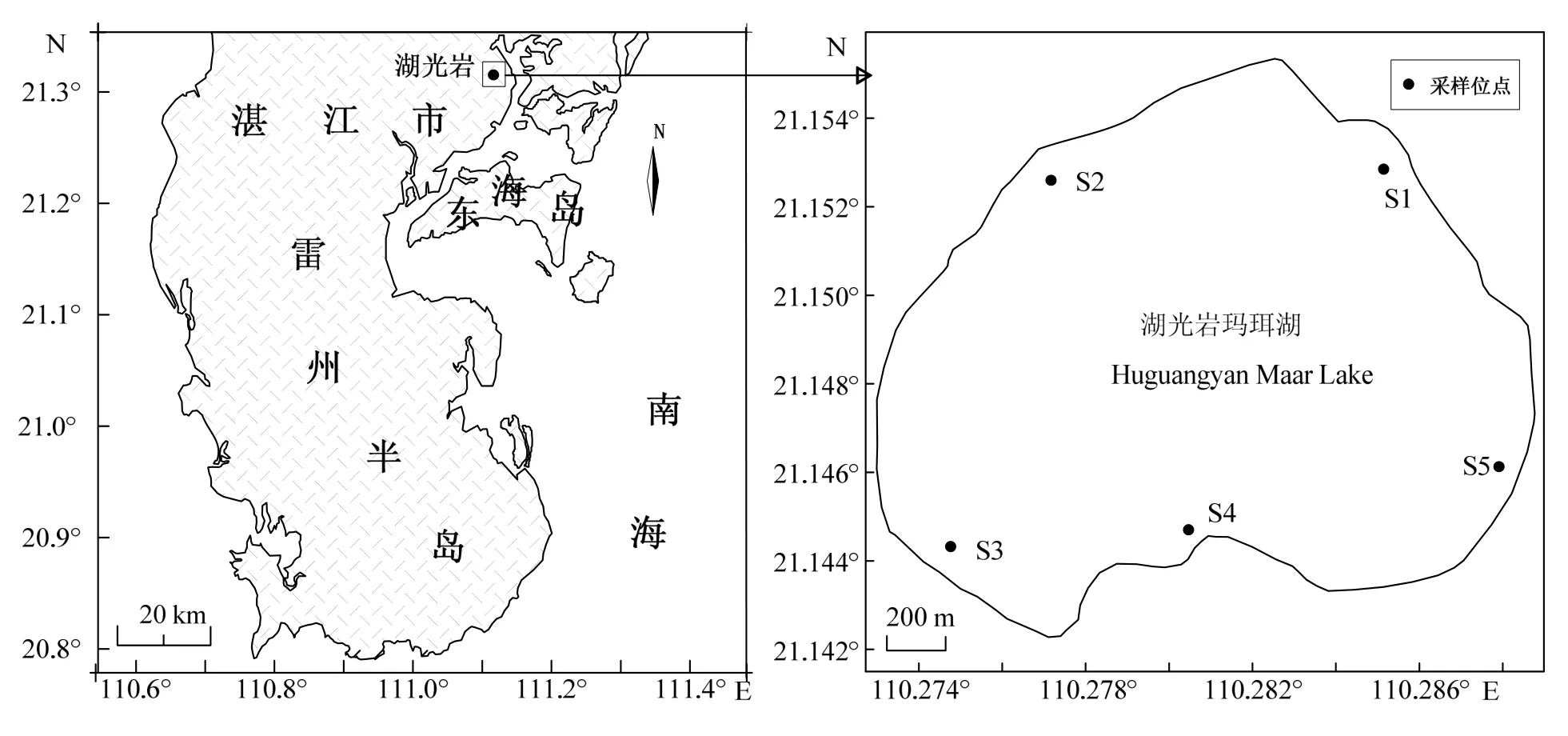
图1 湛江湖光岩玛珥湖采样站位Fig.1 Sampling stations in Huguangyan Maar Lake,Zhanjiang
1.5数据统计分析
进行每月 1次的周年调查,季节依照传统的气象划分法处理,即3 - 5月为春季,6 - 8月为夏季,9 - 11月为秋季,12 - 2月为冬季。浮游植物和浮游细菌生物量与可培养细菌数量时空变化差异性比较均采用单因素方差分析(ANOVA),三者之间的数量变化关系及其与主要生态因子之间关系采用Pearson相关分析,用SPSS16.0软件进行统计。
2 结果与分析
2.1浮游植物、浮游细菌生物量与可培养细菌数量的周年变化
周年调查期间,浮游植物生物量波动范围84.17~243.64 mg/m3(图2),年平均值为130.52 mg/m3,其中,3月生物量最多,为243.62 mg/m3,随后急剧减少,4月出现最低值,为84.17 mg/m3,经小幅波动后,12月出现一个小峰值,为189.01 mg/m3,季节变化模式为春季 > 冬季 > 秋季 >夏季,且春季浮游植物生物量高于夏季,差异有统计学意义(ANOVA,P < 0.05)。整体来说,浮游植物生物量的周年变化也呈双峰型,3月与 12月分别出现一个峰值(图2)。
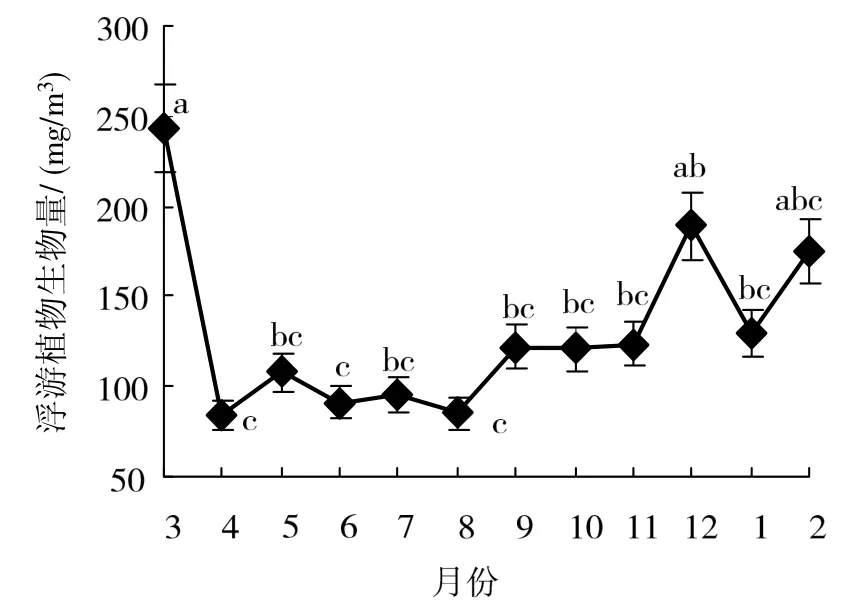
图2 湖光岩玛珥湖浮游植物生物量周年变化Fig.2 Annual dynamics of phytoplankton biomass in Huguanyan Maar Lake
调查期间,浮游细菌生物量的变化范围为20.53~43.60 mg/m3(图 3),年平均值为 30.32 mg/m3,其中,月平均值3月最高,达43.60 mg/m3,8月最低,为20.53 mg/m3。浮游细菌生物量多寡按季节排序依次是春季、秋季、冬季、夏季,且春季生物量高于夏季,差异有统计学意义(ANOVA,P < 0.05),其他季节间差异无统计学意义(ANOVA,P > 0.05)。整体来说,浮游细菌生物量周年变化呈现双峰型,峰值分别出现在 3月和10月。
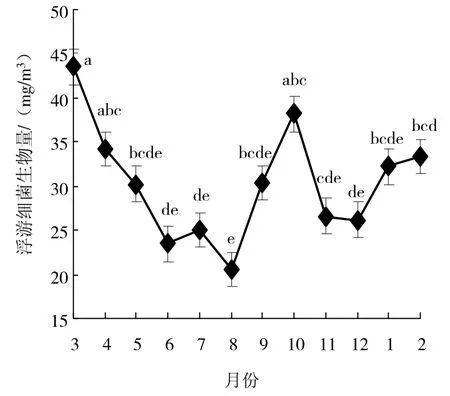
图3 湖光岩玛珥湖浮游细菌生物量周年变化Fig.3 Annual dynamics of bacterioplankton biomass in Huguanyan Maar Lake
不同采样时期可培养细菌数量变化范围为0.10×103~ 3.63×103mL-1(图4),年平均值为0.73×103mL-1,月平均值3月最高,为3.63×103mL-1,1月最低,仅0.10×103mL-1,季节变化模式表现为春季 > 秋季 > 夏季 > 冬季,但不同季节间的细菌数量差异无统计学意义(ANOVA,P >0.05)。从周年数量变化来看,可培养细菌数量呈现3月和9月的双峰型(图4)。
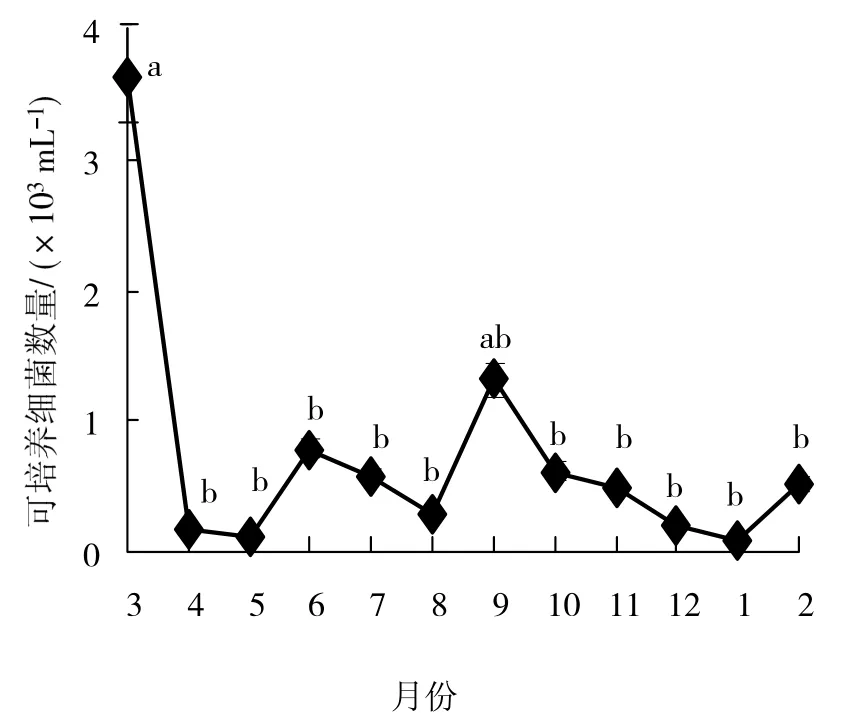
图4 湖光岩玛珥湖可培养细菌数量周年变化Fig.4 Annual dynamics of culturable bacteria densities in Huguanyan Maar Lake
调查结果显示,浮游细菌和浮游植物生物量、可培养细菌数量三者周年变化呈规律性的同步增减趋势。Pearson相关分析表明,三者之间的正相关均有统计学意义(P < 0.05)。
2.2浮游植物、浮游细菌生物量与可培养细菌数量的水平分布
浮游植物生物量年均值大小依次为 S3(154.21 mg/m3)> S5(130.50 mg/m3)> S1(127.22 mg/m3)> S2(124.02 mg/m3)> S4(116.64 mg/m3)(图5),浮游细菌生物量各站位的年均值大小依次为S2(30.99 mg/m3)> S1(30.88 mg/m3)> S3(30.32 mg/m3)> S5(30.24 mg/m3)> S4(29.18 mg/m3)(图6),可培养细菌数在各站位的水平分布却表现为S5(1.67×103mL-1)> S2(0.65×103mL-1)> S4(0.52×103mL-1)> S1(0.44×103mL-1)> S3(0.40×103mL-1)的格局(图7)。浮游植物生物量、浮游细菌生物量和可培养细菌数均存在各自不同的水平分布格局。单因子方差分析(ANOVA)表明,浮游细菌和浮游植物生物量以及可培养细菌数量在各采样站位间的差异性均无统计学意义(ANOVA,P > 0.05)。

图5 湖光岩玛珥湖浮游植物生物量水平分布/(mg/m3)Fig.5 Horizontal distribution of phytoplankton biomass in Huguangyan Maar Lake/(mg/m3)
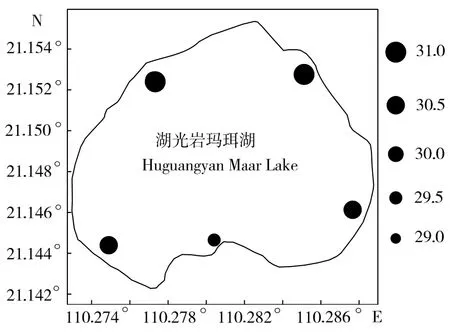
图6 湖光岩玛珥湖浮游细菌生物量水平分布(mg/m3)Fig.6 Horizontal distribution of bacterioplankton biomass in Huguangyan Maar Lake/(mg/m3)
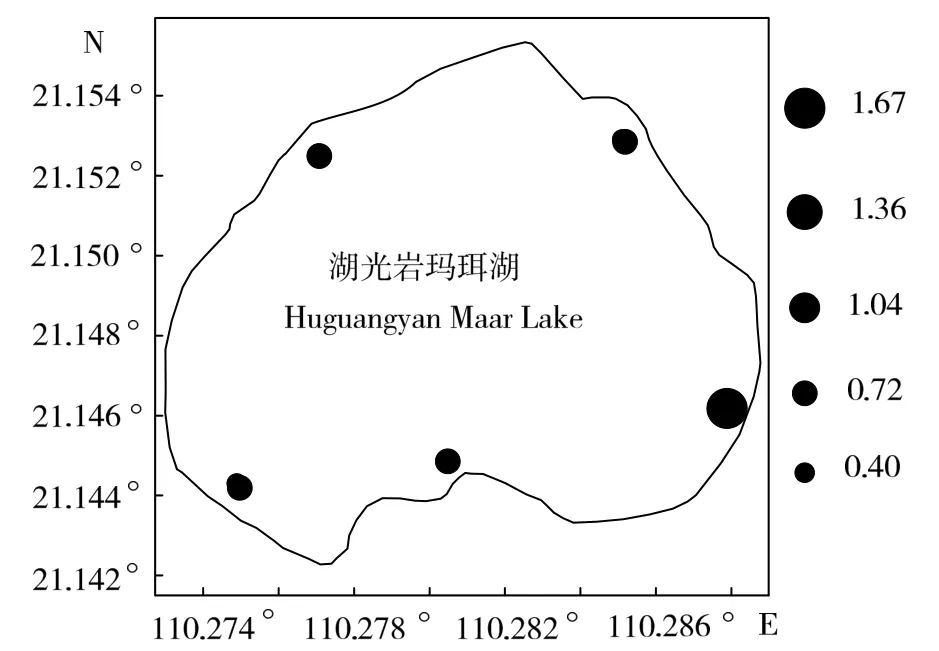
图7 湖光岩玛珥湖可培养细菌数量水平分布(×103 mL-1)Fig.7 Horizontal distribution of culturable bacteria densities in Huguangyan Maar Lake(×103mL-1)
2.3浮游植物生物量、浮游细菌生物量、可培养细菌数量与主要生态因子的关系
浮游植物、浮游细菌生物量,可培养细菌数量与主要生态因子相关分析结果(表1)显示,浮游植物与浮游细菌生物量均与总氮呈显著正相关(P <0.05),而与其他生态因子相关关系无统计学意义(P> 0.05),说明总氮与浮游细菌和浮游植物生物量变化的关系密切;可培养细菌数量与氨氮呈现极显著正相关(P < 0.01),与溶解性氮和磷酸盐呈显著性负相关(P < 0.05)。
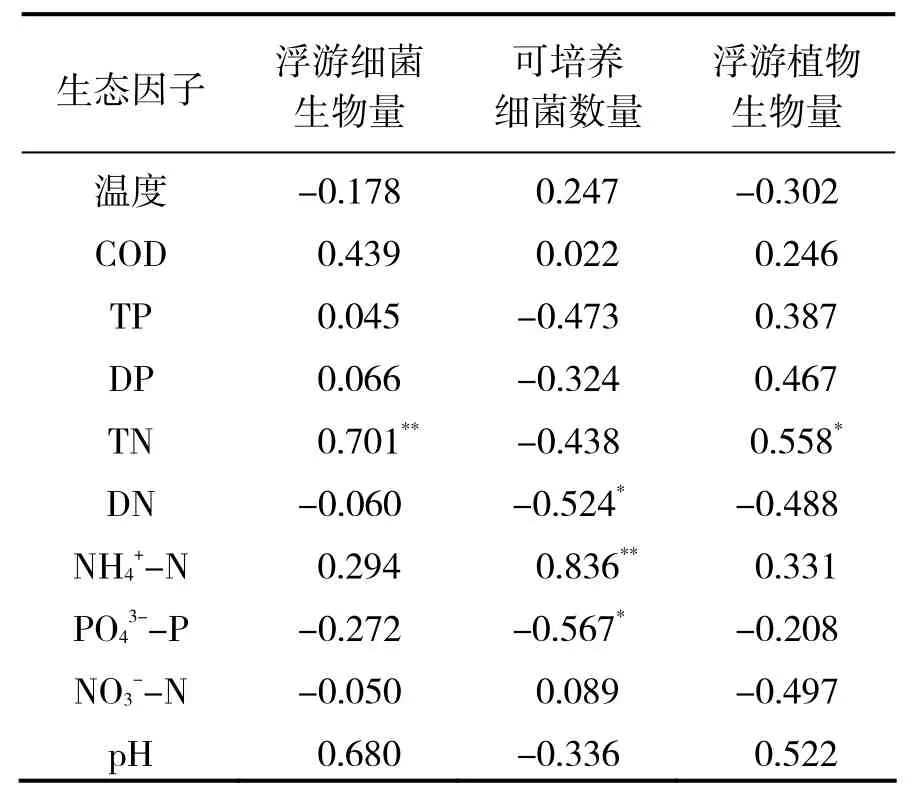
表1 浮游植物生物量、浮游细菌生物量、可培养细菌数量与主要生态因子的相关系数Table 1 Correlation coefficients between phytoplankton biomass, bacterioplankton biomass, culturable bacteria densities and selected ecological factors
3 讨 论
湖光岩玛珥湖浮游细菌与可培养细菌数量变化同步,最高峰值均出现在3月,且9、10月也出现较高的峰值。湖光岩玛珥湖浮游植物生物量在春季达到峰值,可能是由于浮游植物光合作用可产生大量的可溶性有机物,为细菌生长繁殖提供了丰富的营养物质[18],也可能与此时空气相对湿度较大有关;因此,细菌数量3月出现最高值。另外,宋玉芝等[19]在太湖的研究认为,水温对细菌丰度影响较大,且呈显著正相关关系,湖光岩玛珥湖实测水温在9月达到最高值,较高的水温能促进细菌自身的新陈代谢,加速细菌的生长繁殖速率,使得细菌数量在秋季也出现较高峰值。本研究中,浮游细菌最高峰值出现在3月,所以,湖光岩玛珥湖中浮游植物对浮游细菌数量变化的贡献率高于细菌自身繁殖,初级生产可能是调控细菌数量变化重要因子。
浮游植物数量在4个季节中仅出现一个明显的密度高峰,显示其从冬季开始增长,春季最高,而其他季节波动幅度小,这与一般浅水湖泊中的浮游植物生物量呈多峰变化不一致[20],可能与湖泊深度[21]和水温季节性分层[20]有关。玛珥湖是一个封闭式的深水湖泊,由于风场和湖流对其中的有机颗粒物沉积和再循环过程影响作用弱,从而阻碍浮游植物对无机营养盐的吸收[21]。此外,在夏季湖泊易出现水温垂直分层现象,表层水温较高,水体的稳定度较大,阻碍了表层水与较冷的深层水及底层沉积物之间的垂直混合交换,直接影响营养盐在系统内循环的速度和效率[20],导致夏季玛珥湖中营养物质浓度较低,但多雨的夏季又可输入大量外源营养物质[15],保证夏季湖水中具有一定量的浮游植物而不致出现峰谷。秋末和冬初,表层水温开始下降且雨水少,水体出现混合层,使后续的冬、春季表层营养盐含量增加,导致了浮游植物的增殖,其生物量于春季达到最大值。因此,浮游植物生物量周年变化呈相对稳定的单峰型。
浮游植物和细菌之间存在复杂且密切的关系。一方面,细菌能够利用浮游植物光合作用产生的溶解性有机营养物质促进自身繁殖,同时为浮游植物生长提供必要的营养和生长因子,并有效调节浮游植物的微生长环境[2];另一方面,细菌可以通过参与生物竞争、分泌特殊物质等途径抑制浮游植物的细胞生长,甚至裂解其细胞[22]。本研究表明,浮游植物生物量、浮游细菌生物量与可培养细菌数量三者间均呈显著正相关(P < 0.05),这说明在未受到污染的贫营养湖光岩玛珥湖水体中,浮游植物分泌的有机物对细菌的生长刺激起着潜在的主导作用[15];可培养细菌与浮游细菌生物量之间呈显著性正相关(P < 0.05),缘于可培养细菌为浮游细菌的极小部分,相同生境下对外界变化存在着相似的适应性,以至于可培养细菌与浮游细菌数量变化态势一致而呈现显著的正相关。
湖泊中浮游细菌和浮游植物生物量、可培养细菌数量水平分布受湖泊面积及深度[21]、纬度[23]、风浪[24]等外界环境因素影响,但归根结底都是通过影响有机营养物质水平分布从而间接影响生物量的分布[21,23-24]。本研究显示,湖光岩玛珥湖中浮游细菌与浮游植物生物量、可培养细菌数量在 5 个采样站位差异无统计学意义(P > 0.05),这可能与玛珥湖自身特征有关[3],湖光岩玛珥湖较一般湖泊水深深度大但面积较小,易形成相对稳定的水体,底层沉积的有机营养物质很难通过水体垂直交换至表层[21];另外玛珥湖为封闭的环形湖泊,无外部水系流入及湖水自身的流出,从而一定程度减少了与外界系统之间的交换,各生态因子在湖泊中分布较为均匀,导致浮游细菌和浮游植物生物量、可培养细菌数量的水平分布差异不显著。
Pearson 相关分析显示,浮游细菌与浮游植物生物量均与总氮呈显著正相关(P < 0.05),而与其他生态因子相关关系较弱(P > 0.05),这说明湖光岩玛珥湖水体中的大部分氮素被细菌充分利用,合成了细胞蛋白质[25],测定的水体总氮可能大部分来自于浮游藻类和浮游细菌细胞中的氮素。一般来说,水温是影响浮游植物与细菌生长繁殖重要的生态因子[19,26]。但本研究显示,浮游植物和细菌数量变化与水温无显著相关性,这可能与湖光岩玛珥湖所处地理位置有关,该处常年年均气温较高且波动幅度小[15],很难成为生物量周年或季节变化的主导因素。可培养细菌数量与氨氮呈现极显著正相关(P< 0.05),可能缘于其是水体中的分解者,可培养细菌不仅可分解动植物残体,也可降解藻类分泌的溶解性有机物和释放的颗粒性有机物,而氨氮是矿化主要产物之一。此外,细菌需要大量的氨氮来合成自身蛋白[27]。可培养细菌与溶解性氮和磷酸盐呈显著性负相关(P < 0.05),可培养细菌能正常利用营养物质进行生物合成过程是以细菌菌体生理状态达到平衡为前提条件,即细菌自身的 C、N、P的比值[28]。先前的研究认为,全年中湖光岩玛珥湖氮和磷均是营养限制因子[15],不能满足可培养细菌维持此平衡的需要,可能进而影响细菌对其他元素的利用和矿化,导致细菌与营养盐的相关性发生改变[29],两者间的关系有待进一步深入研究。
4 结 论
分析湛江湖光岩玛珥湖中细菌和浮游植物周年动态及其与主要生态因子的关系,结果表明:1)由于湖泊深度和水温季节性分层等原因,浮游植物生物量周年变化呈现单峰模式,最大值出现在春季;2)浮游细菌和可培养细菌数量周年变化同步,均在春、秋季出现峰值,这可能与温度及浮游植物数量有关,其中,初级生产可能是调控湖光岩玛珥湖细菌数量变化的潜在主导因子;3)浮游植物生物量、浮游细菌生物量和可培养细菌数量三者间均呈显著正相关关系(P < 0.05);4)由于湖光岩玛珥湖自身特征,其中的浮游植物和浮游细菌生物量、可培养细菌数量在5个不同采样站位间无显著性差异(P > 0.05);5)浮游细菌与浮游植物生物量均与总氮呈显著正相关(P < 0.05),而与其他生态因子相关性较差(P > 0.05),可培养细菌数量与氨氮呈现极显著正相关(P < 0.05),与溶解性氮和磷酸盐呈显著性负相关(P < 0.05)。
影响湖光岩玛珥湖浮游植物与细菌周年变化的主导因子有待进一步观测研究。
[1]Shchur L,Aponasenko A,Lopatin V,et al.Functional characteristics of bacterioplankton with reference to its aggregation in water bodies of different types [J].Biology Bulletin of the Russian Academy of Sciences,2002,29(5):431-436.
[2]Niu Y,Shen H,Chen J,et al.Phytoplankton community succession shaping bacterioplankton community composition in Lake Taihu,China [J].Water Research,2011,45(14):4169-4182.
[3]刘嘉麒,Negendank J,王文远,等.中国玛珥湖的时空分布与地质特征[J].第四纪研究,2000,20(1):78-86.
[4]Zolitschka B,Brauer A,Negendank J F,et al.Annually dated late Weichselian continental paleoclimate record from the Eifel,Germany [J].Geology,2000,28(9):783-786.
[5]Stebich M,Mingram J,Han J,et al.Late Pleistocene spread of(cool-)temperate forests in Northeast China and climate changes synchronous with the North Atlanticregion [J].Global and Planetary Change,2009,65(1):56-70.
[6]Liu J,Chu G,Han J,et al.Volcanic eruptions in the Longgang volcanic field,northeastern China,during the past 15,000 years [J].Journal of Asian Earth Sciences,2009,34(5):645-654.
[7]吕厚远,刘嘉麒,储国强,等.末次冰期以来湛江湖光岩玛珥湖孢粉记录及古环境变化[J].古生物学报,2003,42(2):284-291.
[8]Nilsson A,Muscheler R,Snowball I,et al.Multi-proxy identification of the Laschamp geomagnetic field excursion in Lake Pupuke,New Zealand [J].Earth and Planetary Science Letters,2011,311(1):155-164.
[9]Mingram J,Schettler G,Nowaczyk N,et al.The Huguang maar lake—a high-resolution record of palaeoenvironmental and palaeoclimatic changes over the last 78,000 years from South China [J].Quaternary International,2004,122(1):85-107.
[10]Kliem P,Enters D,Hahn A,et al.Lithology,radiocarbon chronology and sedimentological interpretation of the lacustrine record from Laguna Potrok Aike,southern Patagonia [J].Quaternary Science Reviews,2013,71(7):54-69.
[11]Wang L,Lu H,Liu J,et al.Diatom-based inference of variations in the strength of Asian winter monsoon winds between 17,500 and 6000 calendar years BP [J].Journal of Geophysical Research:Atmospheres(1984-2012),2008,113(21):1-9.
[12]秦青英,曾永辉,郭倩茹,等.环境16S rDNA和16S rcDNA序列分析湖光岩玛珥湖的浮游细菌组成[J].广东海洋大学学报,2013,33(3):1-9.
[13]郭潇.湖光岩水域细菌的群落结构与功能的研究[D].广东海洋大学,2010.
[14]纪建达,吴灶和,曾永辉,等.湖光岩玛珥湖可培养浮游细菌的 BOX-PCR 图谱及生物多样性分析[J].微生物学通报,2011,38(7):988-999.
[15]张才学,孙省利,谢少英,等.湖光岩玛珥湖的浮游植物[J].水生生物学报,2008,32(5):620-630.
[16]哈希公司.水质分析实用手册[M].北京:化学工业出版社,2009:34-329.
[17]Harris G P.Phytoplankton Ecology:Structure,Function and Fluctuations [M].London:Chapman and Hall Press,1986:65-70.
[18]Josette Garnie,Danielle Benest.Seasonal coupling between phyto- and bacterioplankton in a sand pit lake(Créteil lake,France)[J].Hydrobiologia,1990,207(1):71-77.
[19]宋玉芝,赵淑颖,黄瑾,等.太湖水体附着细菌和浮游细菌的丰度与分布特征[J].环境工程学报,2013,7(8):2825-2831.
[20]Scheffer M.Ecology of shallow lakes [M].Turkey:Kluwer Academic Pub,2004.
[21]Faithfull C,Bergström A-K,Vrede T.Effects of nutrients and physical lake characteristics on bacterial and phytoplankton production:A meta-analysis [J].Limnology and Oceanography,2011,56(5):1703-1713.
[22]周玉航,潘建明.细菌,病毒与浮游植物相互关系及其对海洋地球化学循环的作用[J].台湾海峡,2001,20(3):340-345.
[23]曹金玲,许其功,席北斗,等.我国湖泊富营养化相关指标的空间分布特征[J].环境工程技术学报,2012,2(4):284-289.
[24]Ostrovsky I,Rimmer A,Yacobi Y Z,et al.Long-term changes in the Lake Kinneret ecosystem:the effects of climate change and anthropogenic factors [M]//Climatic change and global warming of inland waters:impacts and mitigation for ecosystems and societies New York:John Wiley & Sons,2013,271-293.
[25]Karafstan A,Ark-Colakoglu F.Physical,chemical and microbiological water quality of the Manyas Lake,Turkey [J].Mitigation and Adaptation Strategies for Global Change,2005,10(1):127-143.
[26]Tian C,Lu X,Pei H,et al.Seasonal dynamics of phytoplankton and its relationship with the environmental factors in Dongping Lake,China [J].Environmental Monitoring and Assessment,2013,185(3):2627-2645.
[27]Ferber L,Levine S,Lini A,et al.Do cyanobacteria dominate in eutrophic lakes because they fix atmospheric nitrogen? [J].Freshwater Biology,2004,49(6):690-708.
[28]李云,李道季.长江口邻近海域浮游细菌分布与环境因子的关系[J].海洋通报,2008,26(6):9-18.
[29]郭凯旋,张瑜斌,章洁香,等.雷州半岛近海夏季浮游植物和浮游细菌生物量的分布及其影响因素 [J].生态学杂志,2012,31(1):8-15.
(责任编辑:刘庆颖)
Annual Dynamics of Phytoplankon Biomass, Bacterioplankton Biomass and Densities of Culturable Bacteria in Huguangyan Maar Lake
HE Yan-yan,LI Qin,ZHANG Yu-bin
(Monitoring Center for Marine Resources and Environments,Guangdong Ocean University,Zhanjiang 524088,China)
The fluorescence microscopic enumeration,spectrophotometry method based on extraction with acetone and plate count method were respectively employed to survey the annual dynamics in phytoplankon biomass,bacterioplankton biomass and densities of culturable bacteria inhabiting in the surface water at five sampling stations in Huguangyan Maar Lake,Zhangjiang.The biomass of phytoplankton and bacterioplankton ranged from 84.17 mg/m3to 243.64 mg/m3,and from 20.53 mg/m3to 43.60 mg/m3,respectively.Nevertheless,the densities of culturable bacteria varied from 0.10×103to 3.63×103cfu/mL.The bacterioplankton biomass,phytoplankton biomass and densities of culturable bacteria revealed the synchronous increase or decline with the peak in March within one year.No significant differences in three investigated objectives were found among 5 sampling stations(P > 0.05).Pearson correlation coefficients showed the significantly positive correlations between the biomass of bacterioplankton and phytoplankton and total nitrogen(TN)(P < 0.05),while the densities of culturable bacteria was significantly positivecorrelative to NH3--N(P < 0.05),and significantly negative correlative to dissoluble nitrogen(DN)and PO43--P(P < 0.05).No significant correlations were discovered between the bacteria,phytoplankton and other selected ecological factors(P>0.05).The primary production from phytoplankton was perhaps a potential key factor regulating the annual variation of bacteria.
Huguangyan Maar Lake; phytoplankton; bacterioplankton; culturable bacteria;annual dynamics
Q938.1; Q948.8
A
1673-9159(2015)01-0062-07
2014-11-18
广东省科技计划项目(2013B020309011)、.广东省教育厅科技创新项目(2013KJCX0095)
贺艳艳(1987-),女,硕士研究生,主要方向为水生微生物生态学。275944326@qq.com
张瑜斌(1970-),男,教授,主要从事水生微生物学与海洋生态学研究。microyubin@163.com
