温度对全缘马尾藻幼孢子体生长和生理组分的影响
2015-11-01孙宗红麦惠欣刘志刚谢恩义
孙宗红,麦惠欣,刘志刚,谢恩义
(广东海洋大学水产学院,广东 湛江,524025)
温度对全缘马尾藻幼孢子体生长和生理组分的影响
孙宗红,麦惠欣,刘志刚,谢恩义
(广东海洋大学水产学院,广东 湛江,524025)
为探究全缘马尾藻(Sargassum intrgerrimum)幼孢子体对温度的适应性,在照度为10 000 lx,光照周期为12L:12D,温度分别为 11.0、16.0、21.0、26.0、31.0、36.0℃的条件下,研究不同温度对全缘马尾藻幼孢子体生长、光合色素含量、SOD活力和CAT活力的影响。结果表明,幼孢子体在16.0~26.0℃的温度范围内生长明显,差异有统计学意义(P<0.05),温度为21.0℃时生长最快;温度为11.0℃时生长缓慢,温度为31.0和36.0℃时幼孢子体的生长因温度过高有溃烂现象。在本实验条件下,全缘马尾藻幼孢子体的适宜温度范围为16.0~26.0℃,最适温度为21.0℃。
温度;全缘马尾藻;幼孢子体;生长;光合色素;酶活力
全缘马尾藻(Sargassum intrgerrimum)隶属于褐藻门(Phaeophyta)圆子纲(Cyclospreae)墨角藻目(Fucales)马尾藻科(Sargassaceae)马尾藻属(Sargassum),主要分布于广东省湛江市硇洲岛的低潮带石沼中,是中国特有的藻类种类,经济价值较大。目前关于马尾藻的研究主要集中在营养成分[1-5]、分布特征[6]以及生态价值[7]等方面。卢虹玉等[1-2]开展了全缘马尾藻的主要营养成分分析与评价,以及其褐藻多酚的抗氧化和抗肿瘤细胞增殖作用研究;黄冰心等[6]报道了我国沿海马尾藻属的物种多样性及其区系分布特征;谢恩义等[7]研究了全缘马尾藻及其他 6种海藻对栽培海区水质的影响;刘加飞等[8]研究了湛江近岸马尾藻中重金属元素含量及富集分析。然而对于与全缘马尾藻的人工育苗密切相关的全缘马尾藻幼孢子体生态学研究较少,开展有关环境因子对全缘马尾藻幼孢子体生长影响的研究十分必要。温度通过直接影响光合作用、呼吸作用等机制来影响幼孢子体的生长、存活率和产量等,是影响全缘马尾藻幼孢子体生长的重要生态因子之一。笔者研究温度对全缘马尾藻幼孢子体生长和叶绿素a(Chl a)、类胡萝卜素(Car)、超氧化物歧化酶(SOD)及过氧化氢酶(CAT)等生理组分的影响,以探讨全缘马尾藻幼孢子体对温度的适应性,为养殖生产提供参考。
1 材料与方法
1.1材料
全缘马尾藻幼孢子体取自2013年8月湛江银浪海洋生物有限公司人工培育的苗帘,生长良好,单株藻体质量0.008~0.011g,长度1~2 cm。
1.2方法
1.2.1实验方案将幼孢子体放于20℃的室内环境中用海水暂养一周,取叶长约1 cm的幼孢子体为实验样本。
预实验表明,10.0℃藻苗存活率低于50%,可视为生存临界温度。因此,实验设置11.0、16.0、21.0、26.0、31.0、36.0℃等6个温度梯度,每个温度梯度组设置3个平行组。在装有500 mL海水的锥形瓶中放入全缘马尾藻幼孢子体30株,放入气石,使锥形瓶中形成水流,以模拟海洋环境中的海水动力条件,瓶口用保鲜膜包裹,以避免因海水蒸发而影响盐度。实验在光照培养箱中进行,各组照度10 000 lx,光照周期12L:12D。实验用水为过孔径75 μm筛网并煮沸的消毒海水,盐度26.0~28.0,pH = 8.1,营养盐为0.12 mmol/L NaNO3和0.0074 mmol/L KH2PO3,每3 d换水1次。
1.2.2测量指标及方法包括全缘马尾藻幼孢子体质量(g)、光合色素含量(Chl a、Car)、酶(包括超氧化物歧化酶SOD、过氧化氢酶CAT)含量和蛋白质浓度的测定。每5 d测1次藻体质量,每7 d测1次光合色素含量,每10 d测1次酶活力和蛋白质浓度。各组每次测定均取幼孢子体2株。
藻体质量:温和除掉杂藻,用纱布吸干幼孢子体表面的水分,称量,计算每个平行组幼孢子体的平均质量。
光合色素含量的测定:采用体积分数95%乙醇提取的分光光度计法。温和除去杂藻后,将全缘马尾藻幼孢子体表面的水分吸干,再进行光合色素含量测定。根据 Porra[9]的公式计算Chl a 含量。根据Parsons & Strickland[10]的公式计算Car含量。

式中,ρChla、ρChlb、ρT、ρCar分别为叶绿素a、b,总叶绿素及类胡萝卜素的质量浓度(mg·L-1),D646nm、D663nm、D470nm分别为各波长下的光密度值。色素含量 = 色素质量浓度(mg · L-1)×提取液体积(L)×稀释倍数 /干样品质量(g)。
超氧化物歧化酶(SOD)活力、过氧化氢酶(CAT)活力、蛋白质浓度的测定均使用南京建成公司的测定试剂盒。总超氧化物歧化酶T-SOD活力单位定义:每毫克组织在 1 mL反应体液中SOD 抑制率达 50% 时所对应的 SOD量为一个SOD 活力单位(U)。过氧化氢酶活力的定义:每毫克组织蛋白每秒钟分解1 μmol的H2O2的量为一个活力单位(U)。本文用比活力(U/mg)形式表达。
1.2.3数据处理运用Excel 2007统计软件进行数据处理和统计分析。用One Way ANOVA检验差异的显著水平(α = 0.05),Duncan法多重比较各组间差异性。
2 结果与分析
2.1温度对全缘马尾藻幼孢子体质量的影响
在不同温度中培养的全缘马尾藻幼孢子体质量的变化见图1。从图1可见,当温度为16.0、21.0和26.0℃时,幼孢子体质量总体上呈增长趋势,25、30、35 d时,各温度组间差异有统计学意义(P<0.05),且当温度为21.0℃时,生长速度最快,其次是16℃和26℃,说明这些温度较适于藻体的生长。31.0、36.0℃温度组的幼孢子体分别在实验30、10 d时因温度过高而全部死亡,不利于藻体生长。11.0℃温度组的藻体质量增长幅度较小,且实验20 d时出现叶子严重脱落,藻体质量急剧下降的现象,说明该温度不利于藻体生长。16.0、21.0、26.0和31.0℃等4个温度组的幼孢子体在实验 15 d时均出现轻微的叶子脱落、藻体质量曲线短暂下降现象,但对生长的影响不大。
2.2温度对全缘马尾藻幼孢子体叶绿素 a含量的影响
在不同温度条件下幼孢子体叶绿素a含量的变化见图2。从图2可见,随温度上升,幼孢子体的叶绿素a含量总体均呈现“高-低-高”变化趋势。随实验时间的变化,11.0、16.0、21.0、26.0℃等温度组幼孢子体的叶绿素a含量总体呈现上升趋势;21℃温度组的幼孢子体叶绿素a含量最高,且同一时间内与其他各温度组差异有统计学意义(P<0.05);16、26℃温度组叶绿素 a含量次之,且同一时间两组间差异均无统计学意义(P > 0.05)。11.0℃温度组的叶绿素a含量增长缓慢,说明低温不利于叶绿素的积累;31.0℃温度组的叶绿素a含量偏低,且随实验时间的变化,呈现先缓慢增长后下降的趋势,且35 d时,该组的幼孢子体因温度过高而均已全部死亡,说明该温度不利于藻体叶绿素的积累。

图1 温度对全缘马尾藻幼孢子体体质量的影响Fig.1 Effects of different temperatures on juvenile sporophyte mass of Sargassum integerrimum
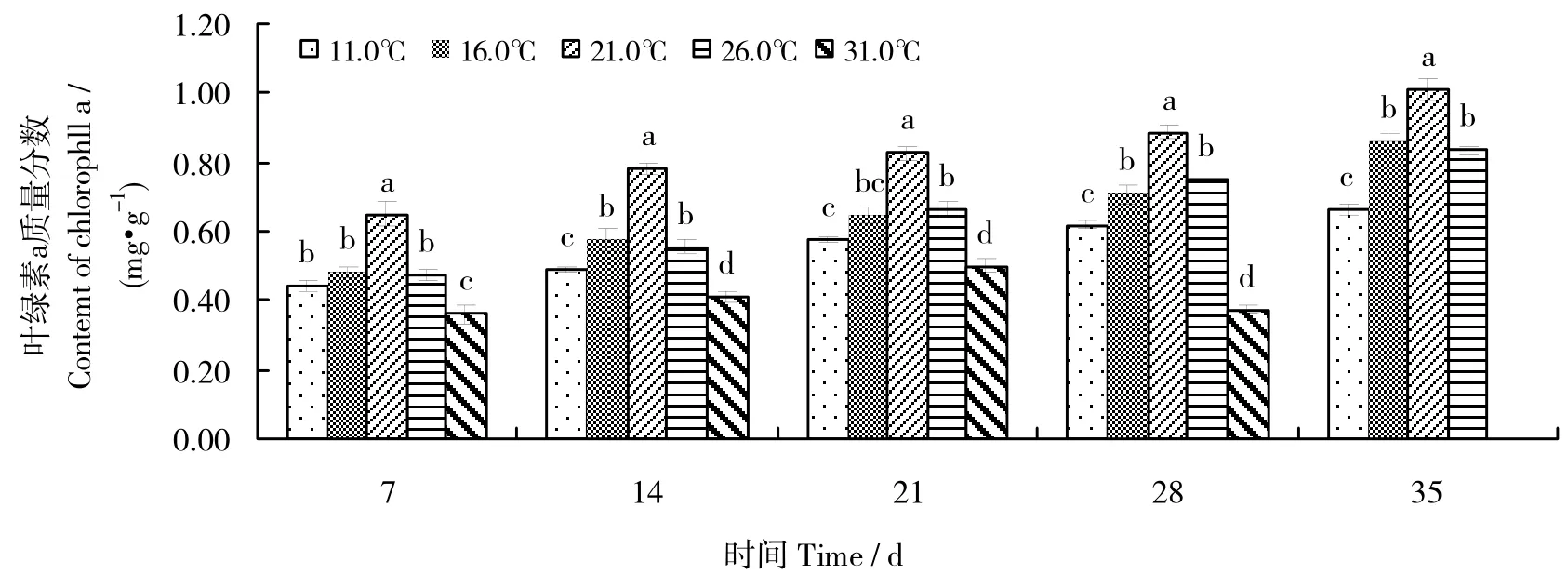
图2 温度对全缘马尾藻幼孢子体叶绿素a含量的影响Fig.2 Effects of different temperatures on the content of chlorophyll a of juvenile sporophyte of Sargassum integerrimum
2.3温度对全缘马尾藻幼孢子体类胡萝卜素含量的影响
不同温度条件下幼孢子体类胡萝卜素的含量变化见图 3。由图 3可知,同一测量时间内,随温度上升,幼孢子体类胡萝卜素含量均呈“低-高-低”的变化趋势。随实验时间的变化,各组的类胡萝卜素含量总体上呈现上升趋势,16、21和26℃等3个温度组上升幅度较大,说明这3个温度均较利于藻体类胡萝卜素的积累,21℃组幅度最高,其次是16℃,再次是26℃组,且同一时间内,16℃和 21℃两组间类胡萝卜素含量差异无统计学意义(P > 0.05)(35 d除外),16和26℃两组间类胡萝卜素含量差异无统计学意义(P > 0.05)(14、21 d除外)。11.0和31℃两组的类胡萝卜素含量较低且上升幅度较小,而31℃温度组21 d时类胡萝卜素含量下降,至35 d时藻体已全部死亡,说明这两个温度不利于藻体类胡萝卜素的积累。
2.4温度对全缘马尾藻幼孢子体SOD活力的影响
不同温度对幼孢子体SOD活力影响见图4。由图4可见,同一时间内,幼孢子体的SOD活力随温度变化,总体上呈现“高-低-高”的变化趋势,而随实验时间的变化,同一温度组的 SOD活力总体上均呈现“先升后降”的变化趋势。当温度为11.0℃时,SOD活力最高,且在同一测量时间内与其他温度组差异有统计学意义(P < 0.05)(20 d的31℃组及40 d的16、26℃两组除外),说明该温度对藻体生长具有胁迫作用,不利于藻体的正常生长。21.0℃条件下,在同一时间内,幼孢子体 SOD活力均低于其他温度组(40 d的31℃组因温度过高藻体均已死亡,SOD活力为0),且在40 d时与其他温度组差异有统计学意义(P < 0.05),说明21.0℃有利于藻体的正常生长。

图3 温度对全缘马尾藻幼孢子体类胡萝卜素含量的影响Fig.3 Effects of different temperatures on the content of carotenoid of juvenile sporophyte of Sargassum integerrimum

图4 温度对全缘马尾藻幼孢子体SOD活力的影响Fig.4 Effects of different temperatures on activity of super oxide dismutase(SOD)of juvenile sporophyte of Sargassum integerrimum
2.5温度对全缘马尾藻幼孢子体CAT活力的影响
不同温度对幼孢子体的 CAT活力影响见图5。由图 5可见,同一时间内,幼孢子体的 CAT活力随温度上升,总体上呈“高-低-高”的变化趋势。而随实验时间的变化,同一温度组的CAT活力总体上呈“先升后降”的变化趋势。同一时间内,21.0℃温度组的幼孢子体CAT活力低于其他温度组,差异有统计学意义(P < 0.05)(30、40 d的31℃温度组,因温度过高藻体部分或全部死亡导致CAT活力较低或为0),说明21℃有利于藻体正常生长;而 11.0℃温度组的幼孢子体CAT活力均高于其他温度组,差异有统计学意义(P < 0.05),说明11℃不利于藻体的正常生长。
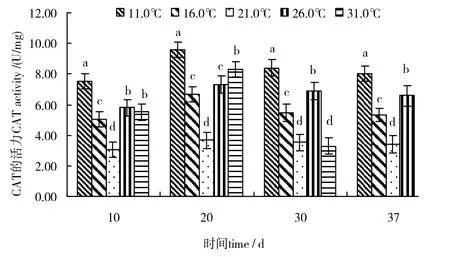
图5 温度对全缘马尾藻幼孢子体CAT活力的影响Fig.5 Effects of different temperatures on activity of catalase(CAT)of juvenile sporophyte of Sargassum integerrimum
3 讨 论
3.1温度对全缘马尾藻幼孢子体生长的影响
温度可影响藻类的光合作用和呼吸作用,是影响全缘马尾藻生长的重要生态因子之一。本研究表明,在11~31℃温度范围内,随着温度的升高,全缘马尾藻幼孢子体质量增长幅度呈现“低-高-低”的变化趋势。温度为11.0℃时,幼孢子体的生长极为缓慢,且实验20 d时出现严重脱叶现象,推断是由于温度过低,光合作用及各种酶活性受抑制所致。在16.0~26.0℃的温度范围内,幼孢子体的生长较快,其中 21.0℃时藻体质量最大,16.0和26.0℃次之。由此推断,本实验条件下,幼孢子体生长的最适宜温度为21.0℃。此结果与Hata M和Yokohama Y[11]的在冬夏两季,大部分海藻光合作用最适温度约为20.0℃的结果一致。当温度为31.0~36.0℃时,全缘马尾藻幼孢子体生长缓慢,且仅可存活一段时间,此结果与贾柽[12]的水温超过27.0℃时,硇洲马尾藻(Sargassum naozhouense Tseng et Lu)幼孢子体苗生长较缓慢的结果一致。这可能与实验温度超过藻体的高温阈值时,影响藻体细胞中酶的活性、叶绿素的合成,并加速了叶绿素的分解有关。
在15 d时,16.0、21.0、26.0和31.0℃的幼孢子体出现少量的落叶现象,导致质量曲线短暂下降。推测落叶现象有两个原因,一是全缘马尾藻幼孢子体的新陈代谢所致;二是旧的藻叶比较细软,且处于老化状态,不能耐受 24 h不间断的水流冲击。
本研究表明,温度影响全缘马尾藻幼孢子体的生长,其幼孢子体生长的适宜温度范围为 16.0~26.0℃,最适温度为21.0℃。姜宏波等[13]研究表明,鼠尾藻(Sargassum thubergii)在15.0和20.0℃条件下生长最快,这与本研究结果相似,进一步证实温度对藻体生长的影响。
3.2温度对全缘马尾藻幼孢子体光合色素含量的影响
藻类的生长是在光合作用合成物质的基础上进行的。叶绿素a是全缘马尾藻最重要的光合色素,其含量大小直接影响光合作用,进而影响藻体的营养及生长。类胡萝卜素是辅助色素,不仅可吸收光能,并传递给叶绿素,还可保护叶绿素。
本研究表明,温度为21.0℃时,全缘马尾藻幼孢子体的叶绿素a和类胡萝卜素的含量均达最大,光合作用最强。Hata M和Yokohama Y[11]研究表明,在冬夏两季,大部分海藻的光合作用最适温度在20.0℃左右。杨彬等[14]研究表明,马尾藻(Sargassum)质量与叶绿素a含量成正相关,王永川等[15]研究发现,瓦氏马尾藻(Sargassum vachellianum)在 20.0℃以下,以及半叶马尾藻[ Sargassum hemiphyllum(Turn.)Ag ]在23.0℃以下的光合作用随温度的上升而增大,超过该温度阀值后光合作用随温度上升而下降。这些结果与本研究结果一致,表明在最适生长温度条件下,藻体光合作用最强,光合色素含量最高。
本研究表明,全缘马尾藻幼孢子体处于温度为11.0℃的低温环境时,其光合色素含量较低,这是低温抑制了藻体细胞中酶的活性和光合色素的合成所致。随着温度逐渐升高,光合色素的合成能力逐渐增强,光合色素含量也相应升高。在21.0℃条件下,光合色素含量达到最大值,之后开始明显下降。当温度超过了藻体的阈值时,不仅藻体细胞内的酶活性和叶绿素的合成受到影响,也加速了叶绿素的分解。这与在实验中观察到 36.0℃和 31.0℃锥形瓶中的海水会不同程度变黄及幼孢子体叶子变白的现象相符合,且36.0℃锥形瓶中的海水颜色比31.0℃锥形瓶中的黄。
本研究表明,温度对全缘马尾藻幼孢子体的光合色素含量的影响显著。全缘马尾藻幼孢子体光合作用的适宜温度范围为16.0~26.0℃,最适温度为21.0℃。这与朱仲嘉等[16]的羊栖菜(Sargassum fusiforme)光合作用的适宜水温约为15.0~25.0℃,以20.0℃为最适水温的结果相近。
3.3温度对全缘马尾藻幼孢子体酶活力的影响
当植物处于逆环境时,植物细胞内的活性氧含量增加,进而损伤植物细胞。此时,植物的抗氧化酶系统会加强植物体内活性氧的代谢。因此,当植物所处逆环境越恶劣,SOD和CAT的活力就越高。朱政等[17]研究表明,一定的低温胁迫条件下,茶树叶片内的SOD活性会有所上升,茶树抗逆性提高。宋新华等[18]亦指出,CAT活性增加可增加植物体对温度胁迫的抗性。本研究表明,11.0和31.0℃组的全缘马尾藻幼孢子体SOD和CAT活力均高于其他温度组(P < 0.05),表明11.0和31.0℃组的全缘马尾藻幼孢子体均处于逆环境中,11.0℃为低温胁迫,31.0℃为高温胁迫。而随着温度升高,幼孢子体中SOD和CAT活力均呈现“高-低-高”的变化趋势,21.0℃时最低。这亦从生长机制方面验证了幼孢子体在21.0℃条件下,生长速度快、光合色素含量高,而在11.0、31.0℃条件下,生长缓慢、光合色素含量低等结果及40 d时31.0℃温度组的幼孢子体均已死亡的结果。由此可见,全缘马尾藻幼孢子体的适宜温度范围为16.0~26.0℃,最适温度为21.0℃。
4 结 论
本研究表明,温度对全缘马尾藻幼孢子体质量,光合色素含量,SOD、CAT活力具有显著的影响;全缘马尾藻幼孢子体的适宜温度范围为16.0~26.0℃,最适温度为21.0℃。
[1]卢虹玉,杨小青,谢恩义,等.全缘马尾藻的主要营养成分分析与评价[J].食品研究与开发,2013,34(7):120-122.
[2]卢虹玉,刘义,吉宏武,等.全缘马尾藻褐藻多酚的抗氧化和抗肿瘤细胞增殖作用研究[J].现代食品科技,2013,29(4):702-705.
[3]肖颖宾,孟庆勇.半叶马尾藻多糖的一般生殖毒性[J].广东海洋大学学报,2012,32(3):19-24
[4]李来好,李刘冬,杨贤庆,等.马尾藻的营养组成及其袋泡茶饮料的试制[J].湛江海洋大学学报,2001,21(1):39-42.
[5]黄俊辉,曾庆孝,佘纲哲.南海海域蜀中海藻总脂质及脂肪酸含量的比较.湛江海洋大学学报,2001,21(2):23-29.
[6]黄冰心,丁兰平,谭华强,等.我国沿海马尾藻属的物种多样性及其区系分布特征[J].海洋与湖沼,2013,44(1):69-76.
[7]谢恩义,申玉春,叶宁,等.流沙湾的底栖大型海藻调查[J].热带海洋学报,2011,30(3):69-75.
[8]刘加飞,谢恩义,孙省利,等.湛江近岸马尾藻中重金属元素含量及富集分析[J].海洋开发与管理,2012(11):71-75.
[9]Porra R J.The chequered history of the development and use of simultaneous equations for the determination of chlorophylls a and b[J].Photosynthesis Res,2002,73(1):149-156.
[10]Parsons T R,Strickland J D H.Discussion of spectrophotometric determination of marine plant pigments,with revised equation for ascertaining chlorophylls and carotenoids[J].J Mar Res,1963,21(3):155-163.
[11]Hata M,Yokohama Y.Photosynthesis-temperature relationships in seaweeds and their seasonal changes in the colder region of Japan [J].Bull Jpn Soc Phycol,1976,24(1):1-7.
[12]贾柽.基于人工育苗的硇洲马尾藻增殖与移植[D].广东海洋大学,2012.
[13]姜宏波,田相利,董双林,等.温度和光照强度对鼠尾藻生长和生化组分的影响[J].应用生态学报,2009,20(1):185-189.
[14]杨彬,谢恩义,曲元凯.不同环境因子对莫氏马尾藻幼苗生长和光合色素的影响[J].南方水产科学,2013,9(4):39-44.
[15]王永川,黄良民,李少芬.温度对几种海藻的光合作用及其分布的影响[J].热带海洋,1983,2(1):55-60.
[16]朱仲嘉,陈培明.羊栖菜马尾藻光合作用与水温、光强的关系[J].水产学报,1997,21(2):165-170.
[17]朱政,蒋家月,江昌俊,等.低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响[J].安徽农业大学学报,2011,38(1):24-26.
[18]宋新华,赵凤云.植物体内过氧化氢酶的研究进展[J].安徽农业科学,2007,35(31):9824-9827.
(责任编辑:刘庆颖)
Effects of Different Temperature on the Growth and Physiological Components of Juvenile Sporophyte of Sargassum integerrimum
SUN Zong-hong,MAI Hui-xin,LIU Zhi-gang,XIE En-yi
(Fisheries College,Guangdong Ocean University,Zhanjiang 524025,China)
Content of photosynthetic pigment and activity of SOD and CAT of juvenile sporophyte of Sargassum intrgerrimum were determined under the conditions of illuminance 10 000 lx,photoperiod 12L:12D,and temperature 11.0,16.0,21.0,26.0,31.0,36.0℃,and the effects of different temperatures on the growth of the juvenile sporophyte were studied to make sure its adaptability to temperature.Results showed that the juvenile sporophyte had a statistical growth(P<0.05)under the temperatures from 16.0 to 26.0℃,and the fastest and the slowest growth were gained at 21.0 and 11.0℃,respectively.The juvenile sporophyte festered due to the high temperature at 31.0 and 36.0℃.Generally,the suitable temperature of the juvenile sporophyte ranged from 16 to 26℃,and the optimal temperature was 21℃.
temperature; Sargassum intrgerrimum; juvenile sporophyte; growth;photosynthetic pigment; enzyme activity
Q949.280.8
A
1673-9159(2015)01-0051-06
2014-05-30
广东省海洋经济创新发展区域示范专项(GD2012-A03-015)
孙宗红(1988-),女,硕士,研究方向为贝类遗传育种及养殖研究。E-mail:zonghongsun@163.com
刘志刚(1963-),男,教授,从事贝类遗传育种及养殖技术研究。E-mail:liuzg919@126.com
