巴什拜羊BPI基因的克隆与真核表达
2015-10-20陈冬梅等
陈冬梅等
摘要:从巴什拜羊外周血多形核粒细胞(PMN)中提取总RNA,经RT-PCR扩增出巴什拜羊BPI基因片段,将其克隆至pPIC9K载体中,构建真核表达质粒pPIC9K-BPI,经PCR、酶切及测序鉴定正确后,电转化至毕赤酵母GS115中并用甲醇诱导表达,SDS-PAGE鉴定重组BPI蛋白的表达,通过微量肉汤稀释法检测重组BPI蛋白的抑菌活性。结果表明:RT-PCR扩增获得597 bp巴什拜羊BPI基因;经PCR、酶切及测序鉴定证实成功构建BPI基因的真核表达载体,SDS-PAGE检测结果表明,重组BPI蛋白成功表达,通过抑菌试验表明表达的重组BPI蛋白具有明显抑菌活性。
关键词:巴什拜羊;杀菌性;通透性增加蛋白;毕赤酵母;基因克隆;真核表达
中图分类号: Q785文献标志码: A文章编号:1002-1302(2015)09-0039-04
收稿日期:2014-09-16
我国是养羊大国,新疆维吾尔自治区是我国主要的绵羊生产基地,巴什拜羊是新疆乃至国内外不可多见的绵羊地方优良品种,具有特殊的生物学特性[1],原产自新疆维吾尔自治区裕民县。该羊具有生长发育速度快、生产性能高、抗病力强等特征,其羔羊成活率达95%以上,是发展我国重要的羊品种资源[2]。杀菌性/通透性增加蛋白(BPI)主要存在于人、哺乳动物的多形核粒细胞(PMN)嗜天青颗粒中,是PMN的主要抗菌成分[3-5],具有杀菌、中和内毒素以及促进补体活化增强调理功能的作用。Vander等发现,BPI蛋白可以诱导血管内皮细胞的凋亡,抑制血管生成,在体外血管生成试验中,在1.8 μmol/L浓度下作用24 h后BPI蛋白能够抑制81%的血管生成,这一发现使BPI蛋白有望用于治疗癌症或与血管形成有关的疾病[6]。BPI蛋白在抗真菌感染[7]、抗弓形虫感染[8]等领域也有非常重要的意义。2005年,美国食品药品管理局批准rBPI21/rBPI23进行Ⅱ~Ⅲ期临床试验,该临床试验主要研究儿童急性脑膜炎菌血症、出血、腹腔感染、肝切除以及胆囊纤维化导致的严重感染,还有其他革兰氏阴性菌、内毒素进入血液循环而造成的致病、致死等症状 [9]。Levy证实了rBPI21不仅可以降低病死率,显著降低血液IL-6的水平,还可以减少感染性并发症和成人呼吸窘迫综合征的发生率[10]。周红等从猪PMN 中提取了猪源BPI 蛋白,发现猪源性BPI 蛋白具有中和内毒素、杀灭革兰氏阴性菌的作用[11] 。朱璟研究BPI基因对猪抗F18大肠杆菌感染的功能及机理[12]。2004年,Alexander等用腺病毒介导的BPI转基因小鼠模型进行体内外试验,均显示腺病毒为载体的重组BPI介导的基因转化可以进行细胞外分泌,且分泌的BPI蛋白可以中和脂多糖LPS,并且减少致炎细胞因子TNF-α和巨噬细胞炎症蛋白-2(MIP-2)的产生[13]。但有关羊BPI基因的功能还未见报道。本试验从巴什拜羊外周血中提取多形核粒细胞PMN总RNA,经RT-PCR获得BPI目的基因片段,构建巴什拜羊BPI真核表达载体,并对其表达产物进行生物活性检测,旨在为进一步深入研究巴什拜羊重组BPI蛋白的生物学功能奠定基础。
1材料与方法
1.1试验动物、菌株与载体
本试验所用的5只巴什拜羊由石河子大学动物科技学院试验站提供。巴斯德毕赤酵母GS115菌株及表达载体pPIC9K为新疆农垦科学院兽医研究所薄新文研究员馈赠,DH5ɑ菌株由石河子大学动物科技学院生化实验室保存。抑菌试验所用的大肠杆菌(Escherichia coli)ATCC25922、沙门氏菌(Salmonella spp.)ATCC14028、金黄色葡萄球菌(Staphylococcus aureu)ATCC25923均为石河子大学动物科技学院中兽医实验室馈赠。
1.2主要试剂
反转录试剂盒,限制性内切酶SnaBⅠ、NotⅠ、SacⅠ和Taq聚合酶均购自TaKaRa公司;T4 DNA连接酶购自天根生化科技有限公司;蛋白marker购自Fermentas公司;酵母基因组DNA快速提取试剂盒、质粒小量提取制备试剂盒、琼脂糖凝胶DNA回收试剂盒、DNA marker DL 2501、DNA Marker DL 2503购自上海捷瑞生物工程有限公司;D-山梨醇、酵母氮碱(YNB)、G418、生物素均购自北京索莱宝科技有限公司;普通肉汤培养基中的蛋白胨、酵母提取物购自上海坤肯生物化工有限公司;中性粒细胞分离液购自天津市灏洋生物制品科技有限责任公司。
1.3目的片段的扩增及重组表达质粒pPIC9K-BPI的构建
巴什拜羊BPI基因cDNA全长序列由笔者所在实验室通过cDNA末端快速扩增技术(RACE)获得,登录号为KF523344。根据该基因的ORF设计活性段表达引物,上游引物为5′-TACGTA ATGACCAACCCAGGCATCGTG-3′,下游引物为5′-GCGGCCGCTCATAGTTTGGTTGTCACTGGCAG-3′。通过颈静脉采集巴什拜羊血液,用肝素抗凝,按照中性粒细胞分离液说明书方法分离外周血中性粒细胞。用Trizol法抽提巴什拜羊中性粒细胞总RNA。提取的总RNA用无菌DEPC水溶解,用琼脂糖凝胶电泳检测RNA的完整性。以提取的PMN总RNA为模板,反转录合成cDNA,再以cDNA模板进行PCR扩增。反应条件:94 ℃ 5 min;94 ℃ 40 s,54 ℃ 30 s,72 ℃ 60 s,32个循环;72 ℃延伸10 min,4 ℃ 保存。用 NotⅠ、SnaBⅠ分别对巴什拜羊BPI基因及pPIC9K表达载体进行双酶切,用T4 DNA连接酶在16 ℃连接过夜,将10 μL连接产物转化于100 μL感受态细胞DH5α,涂布于含有 100 mg/L 氨苄青霉素的LB琼脂板上,于37 ℃恒温箱中静止过夜培养。挑取单个菌落,接种于5 mL含有 100 mg/L氨苄青霉素的LB液体培养基中,37 ℃ 220 r/min振荡过夜培养。取2~3 mL菌液,提取质粒,进行双酶切、PCR鉴定,同时将 1 mL 菌液和5 μL质粒送往北京六合华大基因科技股份有限公司测序,将测序结果正确的重组质粒命名为pPIC9K-BPI。
1.4重组表达质粒pPIC9K-BPI的转化与鉴定
按Riffault等的方法[14]制备感受态酵母菌GS115,取80 μL 感受态GS115与20 μL线性化的重组表达质粒pPIC9K-BPI混合,电击转化后涂布于MD平板上,28 ℃孵育3 d,每天观察菌落的生长情况。挑取单菌落,用酵母基因组DNA快速提取试剂盒提取酵母基因组DNA,以其为模板,分别用表达引物和用酵母载体上的通用引物(α-Factor:5′-TACTATTGCCAGCATTGCTGC-3′、3′AOX1:5′-GCAAATGGCATTCTGACATCC-3′)进行基因型、表型鉴定。PCR产物通过琼脂糖凝胶电泳分析,根据扩增片段的大小判断酵母染色体中是否有目的基因片段插入。基因型鉴定中PCR的反应条件:94 ℃ 5 min;94 ℃ 40 s,63 ℃ 30 s,72 ℃ 40 s,32个循环;72 ℃延伸10 min,4 ℃保存。若能够扩增出1条目的条带,再进行表型鉴定,表型鉴定的反应条件:94 ℃ 5 min;94 ℃ 40 s,54 ℃ 30 s,72 ℃ 60 s,32个循环;72 ℃延伸 10 min,4 ℃ 保存。在表型鉴定中,若能扩增出2条带,一条是含有目的基因片段的扩增条带,另一条是野生型AOX1基因的扩增带,约为 2.2 kb,则为阳性转化子即Mut+(甲醇利用野生型)表型特征。
1.5重组菌株GS115/pPIC9K-BPI的表达与SDS-PAGE分析
将PCR鉴定正确的重组酵母菌GS115/pPIC9K-BPI接种于50 mL YPD培养基中,于28 ℃培养至D值为4时,6 000 r/min离心3 min,收集菌体。将富集的菌体转接于250 mL BMMY培养基中继续培养。每隔24 h加入终浓度为0.5%的甲醇诱导表达。同时按0、24、48、72、96、108 h分别取1 mL菌液样品,离心收集上清、菌体,并将上清、菌体分别进行SDS-PAGE鉴定。
1.6抑菌试验检测表达产物的活性
1.6.1不同浓度的重组BPI蛋白对大肠杆菌、沙门氏菌、金黄色葡萄球菌的抑制作用在配好的普通肉汤培养基中加入重组BPI蛋白,使其终浓度分别为5、10、20、40、80 μg/mL,将大肠杆菌、沙门氏菌、金黄色葡萄球菌稀释至浓度为0.01亿CFU/mL,取30 μL接种于含有各种浓度重组蛋白的肉汤培养基中。另外在不含重组BPI蛋白的肉汤培养基中分别加入30 μL大肠杆菌、沙门氏菌、金黄色葡萄球菌作为阳性对照,不含细菌也不含有重组蛋白的普通肉汤培养基设为阴性对照。在96孔板中以微量稀释法测定重组BPI蛋白的抑菌活性,37 ℃培养12 h后使用酶标仪在600 nm波长下进行观察并记录其D值[15]。重复3次,取平均值(下同)。
1.6.23个不同浓度重组BPI蛋白对大肠杆菌的抑制作用在普通肉汤培养基中加入重组BPI蛋白,使其终浓度为20、40、80 μg/mL,将30 μL大肠杆菌加入其中。阴性对照与阳性对照设置同上。37 ℃下分别培养1、2、3、4、5 h,设置酶标仪在600 nm波长下,对96孔板进行扫描,测定D值,保存数据。
1.7数据处理
数据以“平均值±标准差”表示,采用SPSS 13.0软件对所测数据进行Two-way ANOVA方差分析,比较重组BPI蛋白的抑菌效果。
2结果与分析
2.1RNA的完整性与纯度
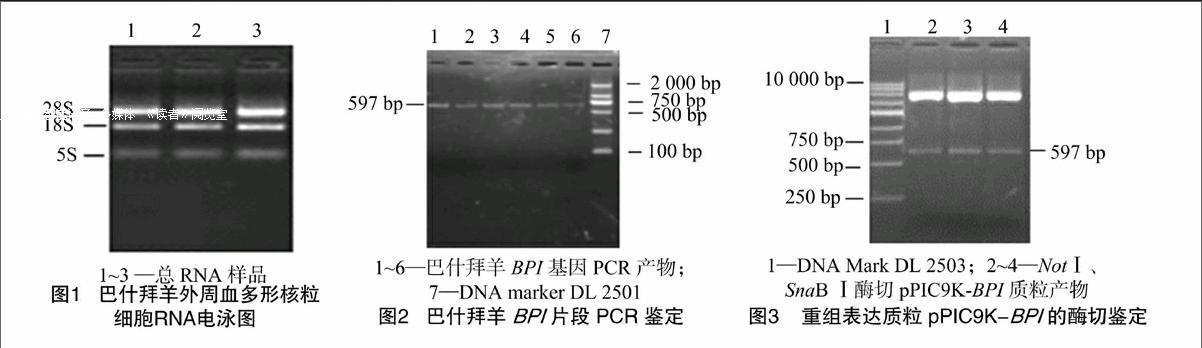
从巴什拜羊PMN中提取的总RNA进行琼脂糖凝胶电泳检测,可见明显的28S、18S、5S条带(图1),说明提取总RNA基本没有降解,具有较好的完整性。对其进行紫外分光光度分析得出D260 nm/D280 nm的值>1.8,表明RNA具有较高的纯度,符合反转录要求。
2.2重组表达质粒pPIC9K-BPI的PCR与双酶切鉴定
重组表达型质粒pPIC9K-BPI经Not、SnaBⅠ双酶切和PCR鉴定,均得到了大小为597 bp的条带,与预期片段大小相符合(图2、图3)。测序结果正确,表明目的片段已正确插入到真核表达载体中。
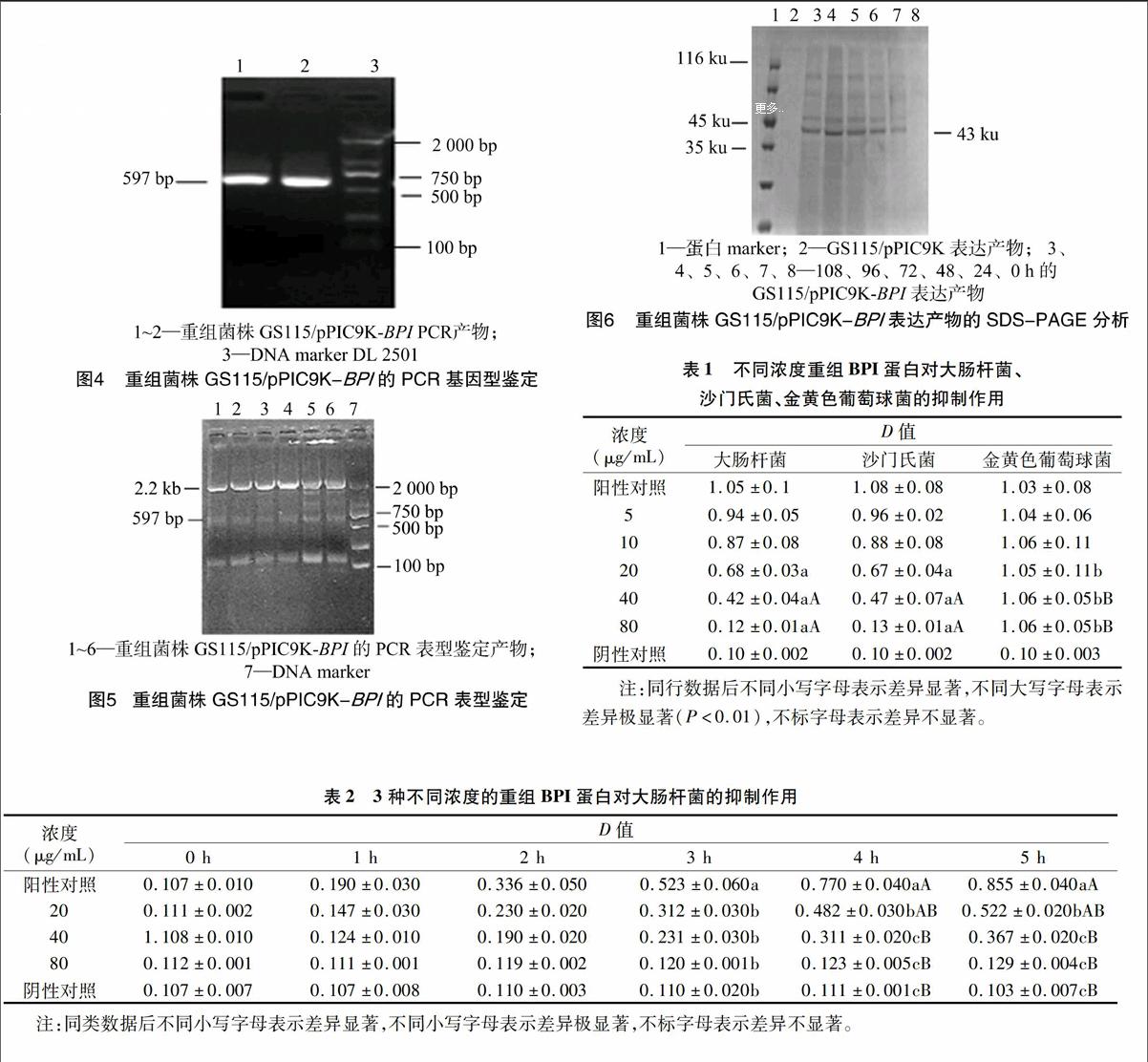
2.3重组菌株GS115/pPIC9K-BPI的PCR鉴定
挑取YPD-G418平板上的单个菌落,以重组菌株的
DNA为模板,分别进行基因型、表型鉴定。在基因型鉴定中可见阳性转化子条带大小为597 bp,与预期条带大小相符合(图4)。在表型鉴定中,用酵母通用引物进行PCR 扩增鉴定,可见阳性转化子能扩增出2条带,一条是含有目的基因片段的扩增条带,约597 bp,另一条是野生型AOX1基因的扩增带,约2.2 kb,为Mut+(甲醇利用野生型)表型PCR鉴定特征(图5)。
2.4重组菌株GS115/pPIC9K-BPI表达产物的SDS-PAGE分析
将重组菌株GS115/pPIC9K-BPI经甲醇诱导表达后,分别收集上清、菌体,上清经过SDS-PAGE分析,在约43 ku处出现条带,与预期目的蛋白分子量相符,在空载质粒对照组和0 h表达载体未见有条带,表明BPI蛋白成功表达(图6),收集的菌体没有表达出目的蛋白。
2.5抑菌试验检测重组BPI的蛋白活性
表1表明,各试验组细菌的阳性对照组(肉汤培养基中
分别加入大肠杆菌、沙门氏菌、金黄色葡萄球菌)D值基本一致,阴性对照组(肉汤培养基)的D值也基本一致。当蛋白浓度为5、10 μg/mL时, 大肠杆菌组、沙门氏菌组与金黄色葡萄球菌组的D值差异不显著。当蛋白浓度为20 μg/mL时,大肠杆菌组、沙门氏菌组的D值显著低于金黄色葡萄球菌组。当蛋白浓度为40、80 μg/mL时,大肠杆菌、沙门氏菌组的D值极显著低于金黄色葡萄球菌组。由此可见,重组BPI蛋白的浓度在20 μg/mL以上可以抑制大肠杆菌、沙门氏菌的生长,对金黄色葡萄球菌没有抑制作用。
3结论与讨论
本试验选用pPIC9K表达载体,并在巴斯德毕赤酵母表达系统进行表达。pPIC9K载体可以快速筛选高拷贝整合转化子,并且具有来自转座子Tn903的卡那霉素耐药基因,能够根据基因剂量效应,对G418的抗性水平快速筛选出高拷贝的转化子,高拷贝整合的转化子能够提高外源基因的表达量[16]。本试验中,笔者充分利用该载体的特性,先进行高拷贝整合的转化子筛选,然后再进行表达,结果发现重组蛋白表达量高达28 mg/mL。巴斯德毕赤酵母作为真核表达系统,具有严格调控外源蛋白的表达、加工修饰表达产物、表达量高、营养要求低等优点。此外,巴斯德毕赤酵母中表达产物可分泌至胞外,有利于表达产物的分离纯化[17]。本试验选用pPIC9K表达载体,构建pPIC9K-BPI表达质粒,并在巴斯德毕赤酵母表达系统进行表达,结果表明,用此技术表达重组蛋白高效、稳定、表达产物易于纯化。天然BPI蛋白由456个氨基酸残基组成的碱性蛋白,含N端、C端、中间连接单位3部分组成,经蛋白酶水解后可裂解为N端片段、C端片段,N端功能片段具有天然BPI的完整杀菌、中和细菌脂多糖(LPS)等活性[18]。目前有2种BPI蛋白获取方法,但使用柱层析法获得的BPI蛋白工作量大,获取量低,成本高。而Gazzno-Santoro等用逆转录PCR的方法获得分子量为23 ku的重组蛋白rBPI23,由于具有较高的活性,且容易获取,广泛应用于临床试验,将人的BPI N-末端的 cDNA 扩增转入细胞载体后通过田仓鼠卵细胞中表达重组蛋白[19]。由于目前国内外采用基因工程方法在猪、鼠、兔和牛上都获得了重组BPI蛋白。在此之前,笔者所在实验室通过RACE技术获得了巴什拜羊BPI基因全长cDNA序列(GenBank登录号为KF523344),本试验根据此序列设计特异性引物进行PCR扩增,克隆出巴什拜羊BPI N-端序列,并在巴斯德毕赤酵母中成功表达出43 ku的含有199个氨基酸的重组BPI蛋白,经过抑菌试验检测该重组蛋白具有明显的抑菌活性,为研究绵羊BPI蛋白的其他生物学功能奠定基础。PMN细胞内诸多抗菌成分中,BPI是唯一能够直接对革兰氏阴性菌发挥杀菌作用和中和内毒素作用的抗菌物质,被誉为“超级抗生素”[20]。本试验中巴什拜羊重组BPI蛋白对G-菌均具有杀菌活性,但对G+无明显效果。研究发现,BPI的N端、C端均含有1个非极性的脂质结合口袋,该非极性口袋为BPI与LPS相互作用的位点。BPI分子N端的碱性氨基酸残基比酸性氨基酸残基多16个,而C端的碱性氨基酸残基比酸性氨基酸残基少2个。BPI蛋白分子N端和C端电荷的不对称分布促使的带正电荷的N端更加能够促进BPI分子与G-菌细胞壁上带负电荷的LPS之间的结合而起杀菌作用[21]。
参考文献:
[1]决肯·阿尼瓦什. 巴什拜羊生物学特性及其遗传多样性研究[D]. 南京:南京农业大学,2010.
[2]决肯·阿尼瓦尔,韩业东,李齐发,等. 巴什拜羊微卫星标记多态性及其与生长指标关联性分析[J]. 中国农业科学,2010,43(16):3425-3432.
[3]Domingues M M,Castanho M A,Santos N C. rBPI(21) promotes lipopolysaccharide aggregation and exerts its antimicrobial effects by (Hemi)fusion of PG-containing membranes[J]. PLoS One,2009,4(12):e8385.
[4]Elsbach P,Weiss J. Role of the bactericidal/permeability-increasing protein in host defence[J]. Current Opinion in Immunology,1998,10(1):45-49.
[5]Marra M N,Wilde C G,Collins M S,et al. The role of bactericidal/permeability-increasing protein as a natural inhibitor of bacterial endotoxin[J]. Journal of Immunology ,1992,148(2):532-537.
[6]Vander S D,Toebes E A,Haseman J R,et al. Bactericidal/permeability increasing protein inhibit sangiogenesis via induction of apopotosis invascular endothelial cells[J]. Blood,2000,96(1):176-181.
[7]de Lucca A J,Walsh T J. Antifungal peptides:novel therapeutic compounds against emerging pathogens[J]. Antimicrobial Agents and Chemotherapy,1999,43(1):1-11.
[8]Arnold H,Horwitz,Paul M,et al. Fungicidal peptides from bactericidal/permeability-increasing protein(BPI) act synergystically with fluconazole on a variety of candida strains[M]. Toronto:ICAAC,1997:928-1001.
[9]程玉磊,祁克宗,潘玲,等. 杀菌性/通透性增加蛋白的生物学功能及其应用[J]. 动物医学进展,2005,26(3):16-18.
[10]Levy O. A neutrophil-derived anti-infective molecule:bactericidal/permeability-increasing protein[J]. Antimicrobial Agents and Chemotherapy,2000,44(11):2925-2931.
[11]周红,郑江,肖光夏. 猪源杀菌性/通透性增加蛋白对革兰阴性菌的作用[J]. 中华传染病杂志,1999,17(1):47-48.
[12]朱璟.BPI基因对断奶仔猪F18大肠杆菌抗性的功能分析[D]. 扬州:扬州大学,2012.
[13]Alexander S,Bramson J,Foley R,et al. Protection from endotoxemia by adenoviral-mediated gene transfer of human bactericidal/permeability-increasing protein[J]. Blood,2004,103(1):93-99.
[14]Riffault S,Carrat C,van Reeth K,et al. Interferon-α-producing cells are localized in gut-associated lymphoid tissues in transmissible gastroenteritis virus(TGEV) infected piglets[J]. Veterinary Research,2001,32(1):71-79.
[15]徐红,陈裕充,温海,等. Neo-Sensitab纸片法与微量稀释法检测常见酵母菌对酮康唑敏感性的比较[J]. 检验医学,2004,19(5):385-386.
[16]Scorer C A,Clare J J,Mccombie W R,et al. Rapid selection using G418 of high copy number transformants of Pichia pastoris for high-level foreign gene expression[J]. Biotechnology,1994,12(2):181.
[17]Yamada M,Azuma T,Matsuba T,et al. Secretion of human intracellular aspartic proteinase cathepsin E expressed in the methylotrophic yeast,Pichia pastoris and characterization of produced recombinant cathepsin E[J]. Biochimica et Biophysica acta,1994,1206(2):279-285.
[18]周宇麒. 杀菌性/通透性增加蛋白(BPI)研究和进展[J]. 医学综述,2001,7(10):588-590.
[19]Gazzano-Santoro H,Parent J B,Grinna L,et al. High-affinity binging of the bactericidal/permeability-increasing protein and a recombinant amino-terminal fragment to the lipid A region of lipopolysaccharide[J]. Infection and Immunity,1992,60(11):4754-4761.
[20]Gray P W,Flaggs G,Leong S R,et al. Cloning of the cDNA of a human neutrophil bactericidal protein,structural and functional correlations[J]. Journal of Biological Chemistry,1989,264(16):9505-9509.
[21]Bülow E,Gullberg U,Olsson I. Structural requirements for intracellular processing and sorting of bactericidal/permeability-increasing protein(BPI):comparison with lipopolysaccharide-binding protein[J]. Journal of Leukocyte Biology,2000,68(5):669-678.
