马铃薯晚疫病菌生理小种研究进展
2015-10-20杨胜先等
杨胜先等

摘要:马铃薯晚疫病是威胁马铃薯生产发展的首要病害,选育和推广马铃薯抗晚疫病新品种是防治马铃薯晚疫病的有效措施。自20世纪80年代以来,由于各地方品种的频繁调运和晚疫病菌A2交配型的迅速蔓延,马铃薯晚疫病在全国范围内有加重趋势。对马铃薯晚疫病菌生理小种研究概况进行综述,为马铃薯抗晚疫病新品种选育及生产品种的布局提供科学依据,并对今后马铃薯晚疫病菌生理小种研究提出工作思路。
关键词:马铃薯;晚疫病菌;生理小种;研究进展;展望
中图分类号: S435.32文献标志码: A文章编号:1002-1302(2015)09-0009-05
收稿日期:2014-12-23
基金项目:国家星火计划(编号:2013GA820003);贵州省科学技术基金(编号:黔科合J字[2013]2004号);贵州省科技成果重点推广计划(编号:黔科合成字[2013]5077);贵州省农业动植物育种项目(编号:黔农育专字[2013]007号)。
作者简介:杨胜先(1976—),男,贵州台江人,硕士,助理研究员,从事作物抗病育种和栽培技术研究。E-mail:ysxnj@163.com。马铃薯晚疫病是由致病疫霉[Phytophthora infestans (Mont.) de Bary]引起的一种真菌性病害,是引起粮食作物产量损失最严重的一种病害,被称为马铃薯生产的头号敌人。1845年,马铃薯晚疫病首次在比利时被报道,随后在荷兰、丹麦、芬兰、法国、意大利、英国等欧洲国家迅速传播;1847年,晚疫病在爱尔兰大面积暴发流行,使当地马铃薯几乎绝收,引起著名的“爱尔兰饥荒”;近年来,马铃薯生产发展不断加快,各地方品种日益频繁的调运使晚疫病致病疫霉的变异进一步加快,生理小种越来越复杂,对抗晚疫病品种的致病性逐年增强,甚至使携带垂直抗病基因的品种(系)丧失抗病能力,给病害的有效防治带来很大困难。马铃薯晚疫病菌生理小种的组成与变异直接关系到病害的发生与流行,对其开展相关研究历年来受到世界各国植物病理学家的重视。笔者对当前马铃薯晚疫病菌的生理小种研究进行综述,为抗晚疫病品种选育及生产布局提供依据,以有效预防马铃薯晚疫病的发生。
1马铃薯晚疫病菌生理小种的鉴定与命名
传统的植物病害研究中,致病型的鉴别常常只对表型进行鉴定分析,这就无法准确预测复杂多变的马铃薯晚疫病菌致病型变化。20世纪40年代,“基因对基因”学说(gene-for-gene hypothesis)问世,该学说认为:植物与病原物实质上是一个相互作用的生物系统,植物对某种病原物的抗性取决于该植物是否具有相应的抗性基因,而病原物对该植物的专一致病性则取决于无毒基因在病原物中的有无,当携带抗病基因R的植物与携带相应无毒基因Avr的病原物相遇时,就会激发植物对病原物的抗病反应,而其他情况下二者则表现亲和,即植物感病[1]。目前,这一学说已在大量的植物-病原物相互作用系统中得到证实[2]。
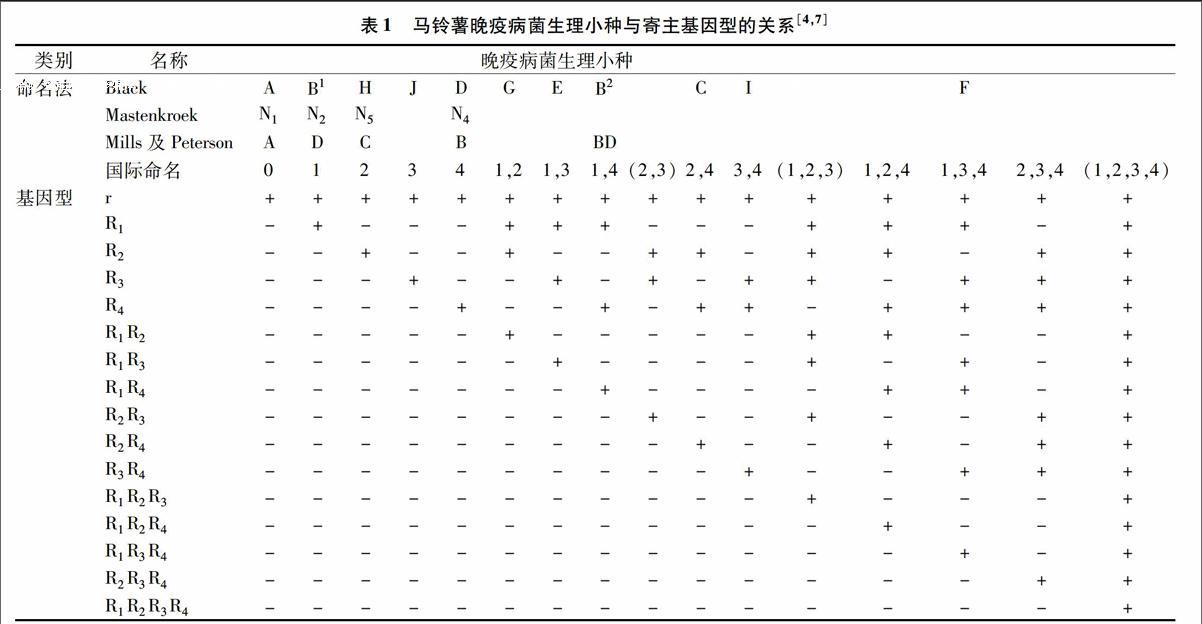
寄主马铃薯单一抗性基因与马铃薯晚疫病菌相应的无毒基因之间的相互作用系统符合“基因对基因”学说,因此,马铃薯晚疫病菌生理小种的鉴定和分类是以基因对基因学说理论为基础的,通常采用携带R0、R1、R2、R3、R4、R5、R6、R7、R8、R9、R10、R11的1个无毒基因和11个抗病基因的1套鉴别寄主,对活体分别进行鉴定,并根据晚疫病菌在这12个鉴别寄主上的反应来确定生理小种的类型[3]。20世纪中期,英国Black等、荷兰Mastenbroek、美国Mills等分别较为系统地研究了寄主抗性的遗传规律及寄主与晚疫病菌生理小种的关系[4-6],并在Black的命名系统基础之上,共同提出了马铃薯晚疫病菌生理小种鉴定及其命名方案(表1)。自从该国际命名方案发表以后,各国关于马铃薯晚疫病菌生理小种的鉴定及命名研究都引用了这一命名系统,该系统对各地的研究结果起到了较好的统一作用。自开展马铃薯晚疫病菌生理小种研究至今,该命名系统在广泛的应用中也得到了不断发展。
2马铃薯晚疫病菌生理小种研究进展
2.1国外马铃薯晚疫病生理小种研究概况
马铃薯晚疫病菌生理小种毒力基因的组成经历了一个从简单到复杂的发展变化过程。1984年,Gurtler报道丹麦马铃薯晚疫病菌生理小种的组成情况,在16个地区发现17个马铃薯晚疫病菌生理小种,毒力基因有1、3、4、5、7、10、11[8];1989年,Tooley等通过分析秘鲁的生理小种组成情况,鉴定得到0、1、1.5共3个优势小种,发生频率分别是33%、39%、15%[9];1994年,Andrivon报道1991—1993年法国马铃薯晚疫病菌的生理小种组成,在116个菌株中共鉴定出19个生理小种,小种组成较为单一,其中小种1.3.4.7.11、1.3.4.7.8.11 和1.3.4.7.10.11在1991年发生最普遍,1.3.4.7.11 、1.3.4.7.10.11分别为1992、1993年的优势小种[10];1998年,Peters等研究加拿大的马铃薯晚疫病菌小种变化情况发现,1996年致病疫霉群体的致病力比1994年明显增强,许多携带垂直抗性基因的抗晚疫病品种丧失了抗病能力[11];2002年,Knapova等鉴定法国和瑞典的生理小种组成,发现91%的菌株都属于1.3.4.7.10.11、1.3.4.6.7.10.11、1.3.4.7.8.10.11、1.3、4.6.7.8.10.11等4个小种[12]; Yigal表1马铃薯晚疫病菌生理小种与寄主基因型的关系[4,7]
注:“+”表示感病,“-”表示抗病;“()”中表示为虚设的生理小种。
通过鉴定1983—2000年以色列的马铃薯晚疫病菌生理小种情况发现,1983—1991年共有5个生理小种,优势小种为13.4.7.8.10, 1993—1998年有19个生理小种,优势小种为1.3.4.7.8.10.11,1999—2000年有42个生理小种,其中新出现的小种有34个[13];Corbiere等报道法国2008年的马铃薯晚疫病菌生理小种组成,发现了3株“超级生理小种”可克服R1~R11这11个抗病基因,而且小种的组成结构趋于复杂[14]。
2.2国内马铃薯晚疫病生理小种研究概况
我国马铃薯晚疫病生理小种的研究工作起始于20世纪60年代,张明厚等开展了马铃薯生育期与马铃薯晚疫病菌生理小种抗性关系的研究[15]。此后,河北、黑龙江、甘肃、山西、内蒙古、青海、北京等地的科研人员陆续报道了当地的马铃薯晚疫病菌生理小种鉴定结果[16-17]。中断一段时间后,20世纪90年代,晚疫病菌生理小种的研究工作又重新开始,刘晓鹏等相继报道了湖北、云南、河北、内蒙古、甘肃等地的马铃薯晚疫病菌生理小种发生情况[18-22]。截至目前,我国黑龙江、内蒙古、湖北、云南、河北、福建、四川、青海、甘肃、山西、吉林、北京等地均报道了马铃薯晚疫病菌生理小种的鉴定结果。由表2可见,50 年来,马铃薯晚疫病菌生理小种在我国各省(市、自治区)发生了激烈的变化,全国各地马铃薯生理小种的组成结构日趋复杂,生理小种的类型不断增多,来自 S. demissum的11个抗性基因已经相继全部被克服,寻找并筛选新的抗病基因是目前马铃薯抗晚疫病育种亟待解决的首要问题。因此,应密切监测各地马铃薯晚疫病菌生理小种的组成及动态变化情况,及时预测现有品种的使用寿命并调整种植品种结构,最大限度地防止马铃薯晚疫病因品种丧失抗性而造成严重损失。
3晚疫病菌A2交配型研究
目前为止,已经发现马铃薯晚疫病菌有A1、A2、自育3种不同的交配型。A1、A2这2种不同的菌体交配型若在晚疫病群体中同时存在,或者在晚疫病群体中有自育型存在时,就可能发生有性生殖并产生有性卵孢子,而卵孢子的特点是壁厚、抗逆性强,可在马铃薯产区顺利越冬,并萌发作为新的侵染源[23-24]。A1交配型存在历史较久,1956年在墨西哥发现了A2交配型[25]。过去一直认为,A2交配型只存在于墨西哥中部,在世界其他地方只有A1交配型,无性繁殖是其他国家马铃薯晚疫病菌的主要繁殖方式。但是,自20世纪80年代初A2交配型在瑞士被报道后[26],A2交配型在瑞典、英国、爱尔兰、埃及、德国、法国、苏格兰、荷兰、波兰、以色列、美国、加拿大、秘鲁、哥伦比亚、玻利维亚、厄瓜多尔、印度、俄罗斯、日本、韩国等世界各国被陆续发现[27-28](表3)。此后,各国科学家们开始针对病菌群体的遗传结构及生物学特性开展大量研究。Goodwin等对采自美国、荷兰等国的A2交配型菌系进行研究发现,这些国家的A2交配型同工酶位点、DNA指纹及线粒体DNA位点与原墨西哥是一致的,说明欧洲国家的A2交配型是由墨西哥传入的[29-30],对其进一步研究发现,美国、波兰、加拿大等国马铃薯晚疫病菌A2交配型的遗传背景与原墨西哥相同[31-32],这进一步说明马铃薯晚疫病菌群体结构变化的主要动力来源于病菌的不断迁移。Masayasu等研究A1、A2菌系在不同培养基上的菌落形态发现,在V8蔬菜汁培养基和土豆片上,A1菌落紧贴培养基,菌丝生长不茂盛,产孢量大,A2菌落棉絮状,菌丝生长茂盛,产孢量小;在燕麦培养基上A1长势较差,A2长势较好,菌落紧贴培养基生长,产孢量较大[33]。1990年Tun-Tschu等发现,在瑞毒霉长期存在的情况下,A1交配型可以转变为A2交配型[34]。我国最早报道发现A2交配型是在内蒙古和山西[35],此后,云南、河北、四川、重庆、黑龙江、福建、甘肃也陆续报道发现有A2交配型[36-40]。目前,除吉林、青海尚未报道发现A2交配型外,其他省份均有相关报道。近年来,马铃薯晚疫病菌研究不断深表2中国各省、市、自治区马铃薯晚疫病生理小种的分化情况
国别报道时间
(年)国别报道时间
(年)墨西哥1956爱尔兰1991东德1993加拿大199l瑞士1984厄瓜多尔1993英国1986哥伦比亚1993荷兰1987玻利维亚1992以色列1989韩国1992埃及1985印度1994瑞典1987中国1996原苏联1991匈牙利1997日本1991尼泊尔1998西德 1986摩洛哥2000巴西1988法国2001波兰1991爱沙尼亚2009美国1991
入,研究手段也日益先进。Goodwin等采用DNA探针技术,通过DNA指纹分析墨西哥中北部马铃薯晚疫病菌的群体结构,发现中部和北部马铃薯的遗传结构明显不同,西北部整个群体的基因型基本一致,而在墨西哥中部遗传多样性非常明显[41]。王鹤通过随机扩增多态DNA(RAPD)引物筛选法,对200个RAPD引物与交配型分子探针连锁的引物进行筛选,获得2个鉴定准确率较高、可用于致病疫霉交配型检测的RAPD引物标记[42]。
4马铃薯晚疫病抗病育种
马铃薯抗晚疫病育种可以上溯至19世纪40年代欧洲马铃薯晚疫病大流行之时,大致历经以晚疫病田间抗性(field resistance)筛选、晚疫病垂直抗性选育和利用、未知R基因存在条件下的泛义水平抗性(broader horizontal resistance)鉴定和选育3个发展阶段[43]。20世纪80年代以来,由于A2交配型的大流行对世界各地马铃薯生产频繁造成严重影响,以化学防治为主要措施的晚疫病综合防控技术及其理论基础已经不能满足生产上对马铃薯晚疫病的防治需求。自1994年起,为建立新的马铃薯晚疫病综合防控技术体系,如全球晚期疫病防治倡议(GILB)、马铃薯晚疫病国际合作项目(PICTIPAPA)、康奈尔-东欧-墨西哥马铃薯晚疫病国际合作项目(CEEM)等多个以晚疫病综合防控为主要目标的合作研究计划先后被发起和实施,并投入大量资金,旨在培育和合理利用持久稳定的高抗马铃薯晚疫病新品种以及开展相关的防治技术和理论基础研究。
在实施马铃薯晚疫病综合防治合作研究计划工作中存在2种不同的研究策略,第1种是以PICTIPAPA计划中的“回归自然”(RETONA)策略为代表,该策略着重强调开发利用已知和未知的R基因抗性,以持久稳定的高抗晚疫病马铃薯新品种、新品系或群体的创制和培育为主要目标,其主要方法是通过分子标记辅助选择或转基因等手段,将已知的R基因或紧密连锁的不同R基因组合,分别导入某一商品性状优良的现有品种不同株系中,分别构建携带有某个R基因或不同R基因组合的“多系群体”(multilineal population),继而有针对性地利用该群体中R基因抗性及其遗传多样性来开展马铃薯晚疫病的防治工作[43];第2种是以CIP及其倡导的GILB计划为代表,其思路是通过构建“无R基因(R-genes free)的马铃薯晚疫病水平抗性育种群体”(population B),并以无R基因马铃薯晚疫病水平抗性新品种的选育和利用为核心,开展晚疫病综合防治等研究工作,该策略现已建立了3个无R基因的晚疫病水平抗性育种亚群,并从这些无R基因的亚群中成功选育得到一系列高抗马铃薯晚疫病的新品种或新品系[44-45]。
我国的马铃薯抗晚疫病育种起始于20 世纪50 年代中期至60 年代中期,通过引进和杂交育种[46],选育出米拉、虎头、跃进、晋薯2 号、克新2号等一系列马铃薯抗晚疫病品种在各地广泛应用,对防治马铃薯晚疫病曾起到一定的积极作用。随着A2交配型在全国各地的陆续出现,生产推广的抗病品种其抗性也逐渐丧失。我国开展马铃薯抗晚疫病育种工作以来,所利用的抗病种质资源多为国外材料,其抗性主要是垂直抗性[47]。20 世纪70 年代,我国开始与国际马铃薯中心合作,引进了大批的抗晚疫病种质材料。近年,又先后引入没有R 基因却具有水平抗性的一系列无性系,在云南、湖北、四川等省进行鉴定研究,并育成川芋4号、川芋5号、华恩1号等[48-49]一批具有水平抗性、综合性状较优良的品种或无性系。
5马铃薯晚疫病菌生理小种研究展望
马铃薯晚疫病病原菌生理小种组成趋于复杂化,类型不断增多,再加上A2交配型的大流行,促使马铃薯晚疫病频发和品种抗病性丧失。综合前人研究结果及马铃薯晚疫病防治实践,笔者认为,今后应从以下3个方面加强对马铃薯晚疫病菌生理小种的研究。
第一,加强马铃薯种质资源的研究力度,挖掘抗性优异种质和抗病基因,培育抗病新品种。生产实践证明,选育马铃薯抗晚疫病品种是防治马铃薯晚疫病最经济、最有效的方法。马铃薯育种工作者可利用RAPD、扩增片段长度多态性(AFLP)、特定序列扩增(SCAR)及简单重复序列(SSR)等分子标记技术发掘抗晚疫病基因,建立完善的抗病基因辅助选择体系,为今后培育多重抗病基因品种提供可能。
第二,统一协调全国各地马铃薯晚疫病研究工作,启动联合攻关专项。选育马铃薯抗晚疫病品种(系),其关键是筛选抗源,查明生理小种的类型和分布情况。A2交配型的广泛流行,不仅加速了生理小种的变异,而且极易产生致病力更强、适应性更广的种群[50]。因此,应启动面向全国甚至全球马铃薯产区范围、以马铃薯晚疫病菌生理小种遗传结构研究及其病害防治为主题的科研专项,并建立切实可行的研究成果共享办法和交流平台,不断加强与国际间的合作攻关力度,为各地区选育、引种马铃薯品种,合理搭配马铃薯生产品种及马铃薯晚疫病综合防治提供科学信息。
第三,进一步拓展和加大马铃薯晚疫病的研究范围和研究深度,开展马铃薯晚疫病菌交配型的发育生物学和流行学机理研究。为从源头上阻止马铃薯晚疫病菌生理小种的生理分化,应大力实施马铃薯晚疫病菌发育生物学、病理学、病害流行学及群体遗传学等研究,积极探索马铃薯晚疫病菌交配型的生长发育、繁殖和流行等生活习性与环境条件的互作机理。综合各学科研究成果,提出能有效控制晚疫病菌交配型正常生长繁殖、变异和流行的方法,以实现从根本上防治马铃薯晚疫病的目的。
参考文献:
[1]de Wit P J G M. Molecular characterization of gene-for-gene systems in plant-fungus interaction and the application of avirulence genes in control of plant pathogens[J]. Annual Review of Phytopathology,1992,30:39l-418.
[2]王庆华,尹小燕,张举仁. 植物的基因对基因抗病性学说[J]. 生命的化学,2003,23(1):23-26.
[3]朱杰华. 中国马铃薯晚疫病菌群体遗传结构研究[D]. 保定:河北农业大学,2004.
[4]Black W,Mastenbroek C,Mills W R,et al. A proposal for an international nomenclature of race of Phytophthora infestans and of genes controlling immunity in Solanum demissum derivative[J]. Euphytica,1953,2(3):173-179.
[5]Mastenbroek C. Investigations into the inheritance of the immunity from Phytophthora infestans de Bary of Solaum demissum Lindl[J]. Euphytica,1952,1(3):187-198.
[6]Mills W R,Peterson L C. The development of races of Phytophthora infestans (Mont.) de Bary on potato hybrids[J]. Phytopathology, 1952,42:26.
[7]张明厚. 马铃薯晚疫病菌[Phytophthora infestans(Mont.)de Bary]生理分化现象的研究概况[J]. 东北农学院学报,1964(1):33-46.
[8]Gurtler H. Physiological races of Phytophthora infestans in Denmark and low temperature storage of isolates[J]. Potato Research,1984,27(1):25-31.
[9]Tooley P W,Therrien C D,Ritch D L. Mating type,race composition,nuclear DNA content and isozyme analysis of Peruvian isolates of Phytophthora infestans[J]. Phytopathology,1989,79(4):478-481.
[10]Andrivon D. Races of Phytophthora infestans in France,1991—1993[J]. Potato Research,1994,37(3):279-286.
[11]Peters R D,Platt H W,Hall R. Changes in race structure of Canadian populations of Phytophthora infestans based on specific virulence to selected potato clones[J]. Potato Research,1998,41(4):355-370.
[12]Knapova G,Gisi U. Phenotypic and genotypic structure of Phytophthora infestans populations on potato and tomato in France and Switzerland[J]. Plant Pathology,2002,51(5):641-653.
[13]Yigal C.Populations of Phytophthora infestans in Israel underwent three major genetic changes during 1983 to 2000[J]. Phytopathology,2002,92(3):300-307.
[14]Corbiere R,Rekad F Z,Galfout A,et al. Phenotypic and genotypic characteristics of algerian isolates of Phytophthora infestans[C]. Researchgate,2010.
[15]张明厚,刘淑静,吕文清. 马铃薯品种各生育期对于晚疫病菌小种抵抗性的比较[J]. 植物病理学报,1965,8(1):17-22.
[16]黄河,程汉清,徐天宇,等. 我国北部马铃薯晚疫病菌生理小种的发生和变化[J]. 植物病理学报,1981,11(1):45-49.
[17]李克来. 马铃薯抗晚疫病和研究方法的探讨[J]. 青海师范大学学报:自然科学版,1988(1):38-45.
[18]刘晓鹏,谢从华,宋伯符. 湖北恩施地区马铃薯晚疫病病菌生理小种的组成及分布[J]. 马铃薯杂志,1995,9(2):81-83.
[19]杨艳丽,胡先奇,鲁绍凤,等. 云南省马铃薯晚疫病菌生理小种的组成与分布[J]. 华中农业大学学报,2007,36(3): 297-301.
[20]张国宝,朱杰华,彭学文. 河北省部分地区马铃薯晚疫病菌生理小种鉴定[J]. 河北农业大学学报,2004,27(1):77-79.
[21]郭军,屈冬玉,巩秀峰,等. 内蒙古马铃薯晚疫病菌交配型和生理小种研究[J]. 西北农林科技大学学报:自然科学版, 2007,35(11):120-124.
[22]成兰芳,张文解,李惠霞,等. 甘肃省马铃薯晚疫病菌生理小种鉴定[J]. 甘肃农业大学学报,2011,46(3):65-69.
[23]Gallegly M E,Galindo J. Mating types and oospores of Phytophyhora infestans in nature in Mexico[J]. Phytopathology,1958,48:274-277.
[24]Galindo A J,Gallegly M E. The nature of sexuality in Phytophyhora infestans[J]. Phytopathology,1960,50(2):123-128.
[25]Niederhauser J S. Division of mycology:the blight,the blighter,and the blighted[J]. Transactions of the New York Academy of Science,1956,19(1):55-63.
[26]Hohl H R,Iselin K. Strains of Phytophyhora infestans from Switzerland with A2 mating type behaviour[J]. Transactions of the British Mycological Society,1984,83(3):529-530.
[27]Sujkowski L S,Goodwin S B,Dyer A T,et al. Increased genotypic diversity via migration and possible occurrence of sexual reproduction of Phytophyhora infestans in Poland[J]. Phytopathology,1994,84(2):201-207.
[28]宋伯符,王军,张志铭,等. 我国马铃薯晚疫病研究的进展和建议[J]. 马铃薯杂志,1996,10(3):138-142.
[29]Goodwin S B,Sujkowski L S,Dyer A T,et al. Direct detection of gene flow and probable sexual reproduction of Phytophthora infestans in northern North America[J]. Phytopathology,1995,85(4):473-479.
[30]Mosa A,Kobayashi K,Ogoshi A,et al. Isoenzyme polymorphism and segregation in isolates of Phytophthora infestans from Japan[J]. Plant Pathology,1993,42(1):26-34.
[31]Goodwin S B,Cohen B A,Deahl K L,et al. Migration from northern Mexico as the probable cause of recent genetic changes in populations of Phytophthora infestans in the United States and Canada[J]. Phytopathology,1994,84(6):553-558.
[32]Koh Y J. Migrations and displacements of Phytophthora infestans population in east Asian countries[J]. Phytopathology,1994,84(9):922-927.
[33]Kato M,Sato N,Mosa A A,et al. Cultural features associated with mating types of Phytophthora infestans isolates from potato crops in Japan[J]. Annals of Phytopathological Society of Japan,1992,58:267-275.
[34]Tun-Tschu C,Wen-Hsiung K. Effect of metalaxyl on mating type of Phytophthora infestans and P. parasitica[J]. Annals of Phytopathological Society of Japan,1990,56(2):194-198.
[35]张志铭,李玉琴,田世民,等. 中国发生马铃薯晚疫病菌(Phytophthora infestans)A2 交配型[J]. 河北农业大学学报, 1996,19(4):62-65.
[36]赵志坚,何云昆,李成云,等. 云南省发现马铃薯晚疫病菌(Phytophthora infestans)A2 交配型[J]. 西南农业学报,1999, 12(3):1-3.
[37]朱杰华,张志铭,李玉琴. 马铃薯晚疫病菌(Phytophthora infestans)A2 交配型的分布[J]. 植物病理学报,2000,23(3):73-75.
[38]张志铭,朱杰华,宋伯符,等. 中国马铃薯晚疫病菌 A2 交配型的进一步研究[J]. 河北农业大学学报,2001,24(2):32-37.
[39]陈庆河,翁启勇,谢世勇,等. 福建省致病疫霉交配型分布及对甲霜灵的抗药性[J]. 植物保护学报,2004,31(2):151-156.
[40]吕伟. 甘肃马铃薯晚疫病菌生物学特性和交配型初步研究[D]. 兰州:甘肃农业大学,2010.
[41]Goodwin S B,Drenth A,Fry W E. Cloning and genetic analyses of two highly polymorphic,moderately repetitive nuclear DNAs from Phytophthora infestans[J]. Current Genetics,1992,22(2):107-115.
[42]王鹤. 2009年中国4省马铃薯晚疫病菌特性分析及其交配型分子标记的开发[D]. 保定:河北农业大学,2011.
[43]李灿辉,隋启君,龙维彪,等. 马铃薯抗晚疫病育种新策略及其研究进展[C]//中国作物学会马铃薯专业委员会2004年学术年会. 中国作物学会马铃薯专业委员会,2004.
[44]Landeo J A,Castelo M,Forbes G,et al. Developing horizotal resistance to late blight in potato[R]. CIP Program Report 1995—1996.
[45]Landeo J A,Castelo M,Beltran G,et al. Quantifying genetic variance for horlzotal resistance to late blight in potatobreeding populationB3C1[R]. CIP Program Report 1999—2000.
[46]黑龙江省农业科学院马铃薯研究所. 中国马铃薯栽培学[M]. 北京:中国农业出版社,1994.
[47]金光辉. 中国马铃薯主要育成品种的种质资源分析[J]. 作物品种资源,1999(4):12-13.
[48]梁远发. 四川省马铃薯种质资源材料引进、评价及利用[J]. 中国农学通报,2005,21(10):348-352.
[49]吴承金,谢从华,宋波涛,等. 马铃薯晚疫病水平抗性新品种——华恩1号[J]. 中国马铃薯,2010,24(4):255-256.
[50]CIP. Late blight:a global initiatire[R]. Convenor:International Potato Center,1996.
