抗枇杷根腐病病菌的6株枇杷主干内生真菌生物学特性研究
2015-10-20陶宏征等
陶宏征等


摘要:为筛选适合工业发酵的抗枇杷根腐病病菌中的枇杷主干内生真菌菌株,对抗枇杷根腐病病菌的6株枇杷主干内生真菌进行生物学特性研究。结果表明:在参试菌株中,DPZG15菌株生长适应性最强,对pH值、光无特殊要求,在马铃薯蔗糖琼脂培养基、麦芽糖或蔗糖为碳源、牛肉膏或酵母膏为氮源、25~28 ℃条件下生长达到最佳水平。
关键词:枇杷;内生真菌;枇杷根腐病;生物学特性
中图分类号: S436.67+9文献标志码: A文章编号:1002-1302(2015)09-0166-03
枇杷(Eriobotrya japonica)为蔷薇科常绿乔木果树,是我国亚热带地区的特色水果[1],含川贝碱、西贝素等多种生物碱[2]。植物内生真菌普遍存在于各种植物健康组织内或细胞间隙,不仅能促进寄主植物生长,还可以产生与寄主相同或相似的次级代谢产物,具有抗菌、杀虫等多种生物活性[3-6]。目前,尚未见枇杷内生真菌的研究报道。枇杷根腐病是枇杷生产上发生严重而普遍的土传病害之一,其田间症状初期表现出树势衰退,叶片起初变黄,之后变成褐色;叶子在脱落之前缓慢下垂;偶尔下大雨时,植株发病较快,只留下褐色干枯的叶片挂在死树枝上;茎干颜色呈暗褐色,基部腐烂,韧皮部呈鱼鳞状;地上部维管组织呈红褐色;后期植株萎蔫、树皮脱落,最后整株枯死。台湾[7]、福建[8]、云南[9]等省份陆续报道有枇杷根腐病发生,损失率高达40%以上,且有逐渐加重之势,制约着枇杷产业发展。化学防治对于土传病害收效甚微,加上化学农药不符合绿色农业生产要求,生物防治日趋受到人们的重视。鉴于此,本研究从枇杷主干内生真菌中挑选对枇杷根腐病病菌抑制率高于50%的菌株[10],对其进行生物学特性研究,为其工业发酵提供理论依据。
1材料与方法
1.1材料
1.1.1枇杷主干内生真菌从蒙自枇杷园区采集枇杷主干韧皮部,常规组织分离[11]获得6株参试菌株,其中菌株DPZG11、DPZG12、DPZG13、DPZG14、DPZG15、DPZG16 均为枝顶孢属(Acremonium)。
1.1.2 供试培养基马铃薯葡萄糖琼脂培养基(PDA):200 g 马铃薯、16 g葡萄糖、20 g琼脂粉、1 000 mL蒸馏水;马铃薯蔗糖琼脂培养基(PSA):200 g马铃薯、16 g蔗糖、20 g琼脂粉、1 000 mL蒸馏水;胡萝卜琼脂培养基(CA):200 g胡萝卜、20 g琼脂粉、1 000 mL蒸馏水;玉米琼脂培养基(MA):30 g 玉米、17 g琼脂粉、1 000 mL蒸馏水;小麦琼脂培养基(WA):30 g小麦、20 g琼脂粉、1 000 mL蒸馏水;枇杷琼脂培养基(LA):200 g枇杷叶、20 g琼脂粉、1 000 mL蒸馏水;察氏 培养基:2.00 g硝酸钠、1.00 g磷酸二氢钾、0.50 g氯化钾、0.50 g七水硫酸镁、0.01 g硫酸铁、30.00 g蔗糖、20.00 g琼脂粉、1 000 mL蒸馏水。所用试剂均为分析纯。
1.1.3试剂碳源为可溶性淀粉、麦芽糖、蔗糖、α-乳糖、葡萄糖、甘露醇;氮源为酵母膏、牛肉膏、蛋白胨、尿素、硫酸铵、硝酸铵、磷酸二氢铵;均为分析纯。
1.2试验方法
1.2.1测定不同培养基对参试菌株菌丝生长的影响将枇杷主干内生真菌在PDA平板培养基中,28 ℃恒温扩大培养 7 d[11],在培养基同一半径周围用打孔器取直径为5 mm的菌饼,同时接种于PDA、PSA、CA、MA、WA、LA 6种培养基平板中央,设3次重复,在28 ℃下恒温培养,7 d后用十字交叉法[11]测定各菌株菌落直径。
1.2.2测定碳、氮源对参试菌株菌丝生长的影响以查氏培养基为基础培养基,分别用相等质量分数的碳源(可溶性淀粉、麦芽糖、α-乳糖、葡萄糖、甘露醇)和氮源(酵母膏、牛肉膏、蛋白胨、尿素、硫酸铵、硝酸铵、磷酸二氢铵)替换蔗糖和硝酸钠,接种及测量方法同“1.2.1”节相关步骤[11]。
1.2.3测定不同pH值对参试菌株菌丝生长的影响以PDA为供试培养基,用0.1%盐酸、0.1%氢氧化钠将pH值调至4、5、6、7、8、9、10,接种及测量方法同“1.2.1”节相关步骤[11]。
1.2.4测定不同温度对参试菌株菌丝生长的影响以PDA为供试培养基,接种后分别在5、10、15、20、25、30、35、40 ℃下恒温培养,接种及测量方法同“1.2.1”节相关步骤[11]。
1.2.5测定不同光处理对参试菌株菌丝生长的影响以PDA为供试培养基,接种后分别在光暗交替(12 h光照、12 h黑暗)、全黑暗、全光照3种光处理下培养,接种及测量方法同“1.2.1”节相关步骤[11]。
以上所有培养基用高压蒸汽灭菌锅121 ℃灭菌25 min。
1.2.6数据统计所有试验数据均采用SPSS 19.0软件的Duncans多重比较法进行统计分析,计算处理间的差异显著性。
2结果与分析
2.1不同培养基对参试菌株菌丝生长的影响
表1显示,参试菌株DPZG11在PDA、PSA 2种培养基间菌落直径差异不显著,在WA、LA 2种培养基间差异显著,在其余2种供试培养基差异极显著;DPZG12、DPZG13在CA、MA 2种培养基间菌落直径差异不显著,但与其余4种供试培养基差异极显著;DPZG14在PSA、MA 2种培养基间菌落直径差异不显著;DPZG15在供试的6种培养基间菌落直径差异极显著;DPZG16在PSA、CA 2种培养基间菌落直径差异不显著,在WA、LA 2种培养基间差异也不显著,但与其余2种供试培养基差异极显著。其中,菌株DPZG14、DPZG16 菌丝生长最适合的培养基为PDA,菌株DPZG12、DPZG13菌丝生长最适合的培养基为PSA,DPZG15菌丝生长最适合的培养基为CA,DPZG11菌丝生长最适合的培养基为PDA或PSA。结果说明,参试菌株营养适应性存在多样性。
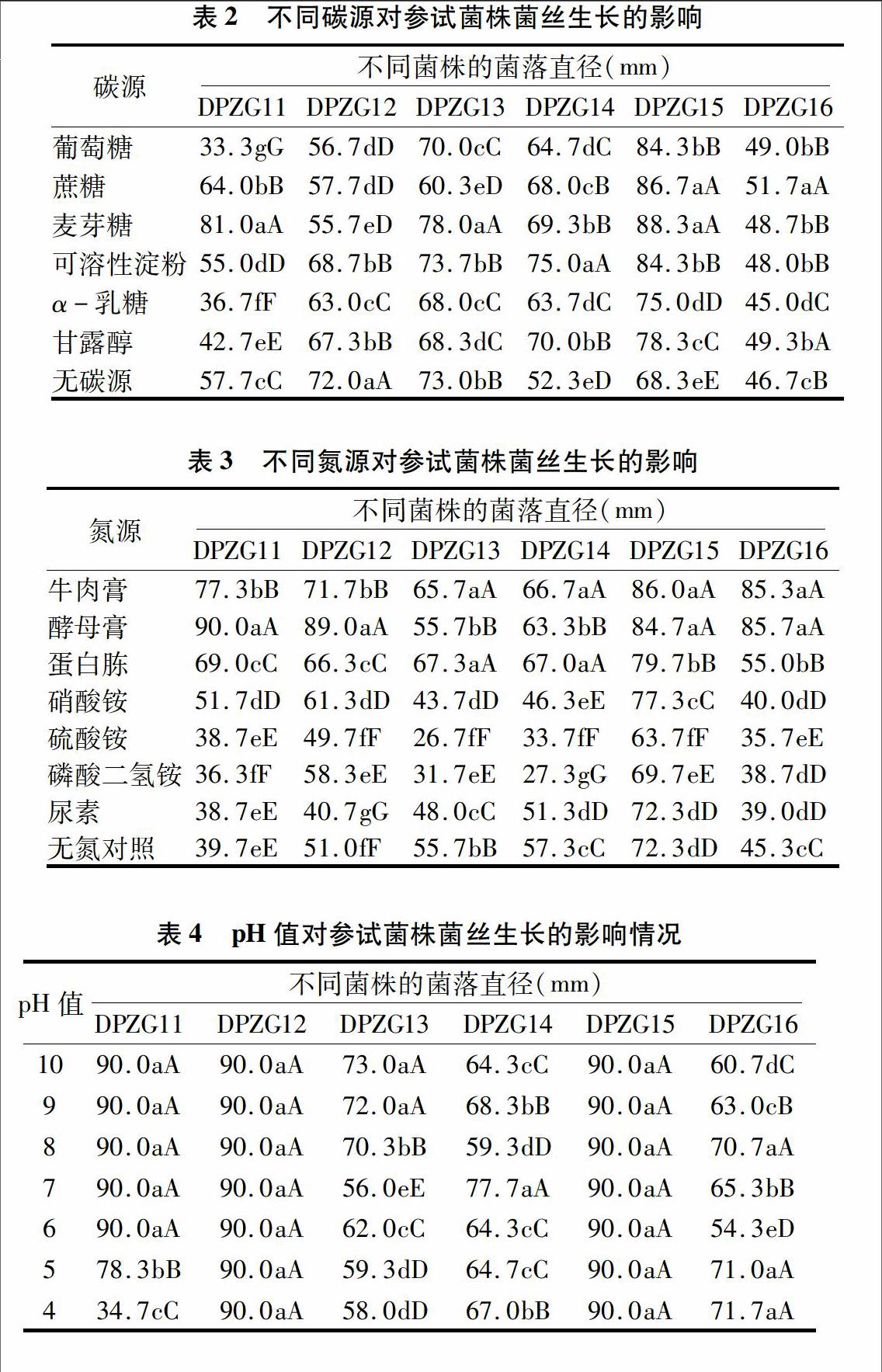
2.2不同碳源对参试菌株菌丝生长的影响
由表2显示,参试菌株DPZG11在供试的6种碳源间菌落直径差异极显著;DPZG12在葡萄糖、蔗糖2种碳源间菌落直径差异不显著,在可溶性淀粉、甘露醇2种碳源间菌落直径差异也不显著,但与α-乳糖差异极显著;DPZG13在葡萄糖、α-乳糖2种碳源间菌落直径差异不显著,但与其余3种供试碳源差异极显著;DPZG14在麦芽糖、甘露醇2种碳源中菌落直径差异不显著,在葡萄糖、α-乳糖 2种碳源间菌落直径差异也不显著,但与可溶性淀粉差异极显著;DPZG15在葡萄糖、可溶性淀粉2种碳源间菌落直径差异不显著,在蔗糖、麦芽糖2种碳源间菌落直径差异也不显著,但与其余2种供试碳源差异极显著;DPZG16在葡萄糖、麦芽糖、可溶性淀粉3种碳源中菌落直径差异不显著,所有碳源均与α-乳糖差异极显著。其中,DPZG11、DPZG13菌株菌丝生长最适合的碳源为麦芽糖,DPZG14最适合的碳源为可溶性淀粉,DPZG15最适合的碳源为蔗糖或麦芽糖,DPZG16最适合的碳源为蔗糖或甘露醇,DPZG12在不添加碳源的培养基中菌落直径最大,对碳源无依赖性,结果表明,参试菌株碳源适应性同样存在多样性。
2.3不同氮源对参试菌株菌丝生长的影响
由表3显示,参试菌株DPZG11在硫酸铵、尿素2种氮源
2.5不同温度对参试菌株菌丝生长的影响
由表5显示,参试菌株在10~35 ℃范围内均能生长。DPZG11菌株菌丝生长最适合的温度为25~30 ℃;DPZG12最适合的温度为25 ℃;DPZG13、DPZG14最适合的温度为30 ℃;DPZG15最适合的温度为25~28 ℃;DPZG16最适合的温度为20 ℃。说明各参试菌株的温度适应性也存在差异。
2.6不同光处理对参试菌株菌丝生长的影响
由表6显示,在光暗交替(12 h光照、12 h黑暗)、全黑暗、全光照3种光处理下,DPZG11、DPZG12、DPZG15菌株菌落直径均达90.0 mm,生长不受光的影响;DPZG13、DPZG14在全光照时菌落直径最大;DPZG16菌株全黑暗最适合其菌丝生长。说明参试菌株对光的适应性也存在差异。
3结论与讨论
在6种供试培养基中,PDA、PSA、CA培养基最适合各参试菌株菌丝生长,此研究结果与笔者所在课题组报道的另外6株枇杷内生真菌的研究结果[12]一致。在供试碳源中,麦芽糖、可溶性淀粉、蔗糖、甘露醇最适合各参试菌株菌丝生长,此结果与笔者所在课题组报道的另外6株枇杷内生真菌的研究结果[12]不一致。在供试氮源中,牛肉膏、酵母膏、蛋白胨最适合各参试菌株菌丝生长,此结果与笔者所在课题组报道的其余6株枇杷内生真菌[12]、银杏内生真菌[13]的研究结果一致。20~30 ℃最适合各参试菌株菌丝生长,此结果与鹿蹄草属内生真菌的研究结果[14]不一致。DPZG12、DPZG15生长不受pH值及光照限制,其他4个菌株最适合的pH值及光照各不相同,此研究结果与笔者所在课题组报道的另外6株枇杷内生真菌的研究结果[12]不一致。试验结果表明,枇杷主干中的各内生真菌菌株生长喜好不尽相同,其中DPZG15菌株生长适应性最强,较其他菌株更具开发应用优势。
各参试菌株均为枝顶孢属真菌,叶利芹等曾研究赭红枝顶孢(Acremonium salmoneum)培养条件[15]及其竹叶锈病菌的重寄生作用[16]。枝顶孢属作为杀虫生防制剂的研究也有报道[17-18]。另外,黎起秦等用针刺接种法和浸根法将内生菌B47成功地定殖到番茄体内[19]。江木兰等通过浸种法把油菜内生生防菌BY-2定殖到油菜体内[20]。李红刚等又通过研磨组织涂抹法把内生放线菌定殖植株体内[21]。而定殖的生防菌株受到植物组织的良好保护[22],这为今后DPZG15定殖到植株体内提供了理论支持和经验借鉴。
参考文献:
[1]江国良. 枇杷在四川不同生态型区的生态适宜性及调控技术研究[D]. 雅安:四川农业大学,2011:1-152.
[2]何志刚,林晓姿,李维新,等. 枇杷的营养保健与川贝枇杷低糖果酱的研制[J]. 食品科学,2005,26(9):258-260.
[3]文才艺,吴元华,田秀玲. 植物内生菌研究进展及其存在的问题[J]. 生态学杂志,2004,23(2):86-91.
[4]黎万奎,胡之璧. 内生菌与天然药物[J]. 中国天然药物,2005,3(4):193-199.
[5]郭良栋. 内生真菌研究进展[J]. 菌物系统,2001,20(1):148-152.
[6]王维,马养民,张弘弛,等. 黑果枸杞内生真菌E21菌株次生代谢产物的研究[J]. 中国新药杂志,2013,22(4):460-464.
[7]Chern L L,Ann P J, Young H R. Root and foot rot of loquat in Taiwan caused by Phytophthora[J]. Plant Dis. 1998,82:651-656.
[8]庄文远,吴志珍,曾忠坚.枇杷根腐病的发生与防治技术[J]. 广西植保,2002,15(1):8-9.
[9]张莹,陈帆,冯光荣,等. 蒙自枇杷生长结果习性及栽培技术[J]. 云南农业科技,2009(6):25-28.
[10]鲁海菊,董梅,崔同敏,等. 从枇杷内生真菌中筛选抗枇杷根腐病菌的活性菌株[J]. 江苏农业科学,2014,42(1):95-97.
[11]方中达. 植病研究方法[M]. 3版.北京:中国农业出版社,1998:57-125.
[12]鲁海菊,郝小燕,崔同敏,等. 抗枇杷根腐病病菌的枇杷主干内生真菌生物学特性研究[J]. 北方园艺,2014(8):108-111.
[13]陈凤美,刘群,蒋继宏,等. 银杏内生链格孢菌GI009生物学特性研究[J]. 西北林学院学报,2005,20(4):112-114,121.
[14]孙梅青,陈忠,喻其林,等. 鹿蹄草属内生真菌生物学特性研究[J]. 中国农学通报,2011,27(4):125-128.
[15]叶利芹,吴小芹,王靓. 竹叶锈病重寄生菌赭红枝顶孢(Acremonium salmoneum)培养条件的研究[J]. 南京林业大学学报:自然科学版,2012,36(2):64-68.
[16]叶利芹,吴小芹,叶建仁. 赭红枝顶孢Acremonium salmoneum对竹叶锈病菌的重寄生作用研究[J]. 中国生物防治学报,2011,27(4):528-534.
[17]宋丽雯. 顶孢霉菌株(Acremonium hansfordii)的生物学特性和对蚜虫致病机理的研究[D]. 兰州:甘肃农业大学,2006:1-56.
[18]王琰. 10%顶孢霉(Acremonium hansfordii)可湿性粉剂加工工艺及药效评价[D]. 兰州:甘肃农业大学,2010:1-55.
[19]黎起秦,罗宽,林纬,等. 内生菌B47的定殖能力及其对番茄青枯病的防治作用[J]. 植物保护学报,2006,33(4):363-368.
[20]江木兰,赵瑞,胡小加,等. 油菜内生生防菌BY-2在油菜体内的定殖与对油菜菌核病的防治作用[J]. 植物病理学报,2007,37(2):192-196.
[21]李红刚,马林,刘慧平,等. 两株植物内生放线菌在植株体内定殖能力的测定[J]. 山西农业大学学报:自然科学版,2008,28(3):287-289.
[22]印敬明,刘晓光,万慧,等. 螺旋毛壳(Chaetomium spirale)ND35防病促生作用初探[J]. 莱阳农学院学报,2006,23(4):272-275,279.吴秀臣,芦建国. 南京城区绿地的外来入侵植物[J]. 江苏农业科学,2015,43(9):169-172.
