生物成因气生成演化模式探讨
2015-10-18林小云高甘霖
林小云,高甘霖,徐 莹,任 莹,薛 颖
(油气资源与勘探技术教育部重点实验室 长江大学,湖北 武汉 430100)
引 言
生物成因气分布广,埋藏浅,为天然气水合物的主要气源,将成为人类重要的接替能源[1-2],引起世界各国的重视。
研究生物成因气源对评价天然气水合物资源潜力具有重要的理论和现实意义。20世纪80年代以来,国内外对生物气的成因进行了研究[1-5],大多数学者基本认同生物成因气生成过程是一种包括各种辅酶在内的独特的生物化学反应,在微生物的作用下,通过CO2还原或者乙酸发酵形成甲烷。有机质在产甲烷菌的作用下最终生成生物气的过程是一样的,但由于在不同深度,有机质所处的环境和微生物种类的不同,有机质被降解的过程以及形成的产物都会有明显的区别,直接影响到生物气的生成量及特征[3-5]。
通过生烃模拟实验资料及生物成因气藏实例分析,系统总结生物气源岩有机质演化过程和生物成因甲烷气生成过程的阶段性及特征,建立生物气生成演化模式,为生物气源岩生气能力评价提供依据。
1 生物成因气类型及分布
1.1 生物成因气的分类
生物成因气是在微生物作用下生成的,以甲烷为主的天然气,根据生气时间和母质以及地质条件的不同,可分为原生型和次生型两大类。原生传统生物气是有机质在未成熟阶段(Ro≤0.5%)经厌氧细菌生物化学降解的气态产物[6-9];若由泥炭或低煤级煤的有机质在煤化作用早期经细菌分解等复杂过程形成则为煤层生物气[10-14];介于生物甲烷生成高峰之后和成熟石油生成高峰之前(Ro为0.3%~0.6%)有机质中的化学活性热不稳定组分经低温生物化学或(和)低温化学反应合成气态烃类,称为生物 -热催化过渡带气[12-14]。次生型生物气则是经过一定热演化的有机质及其产物,由于构造抬升等再次受到微生物作用而形成,根据有机质类型和微生物作用的不同,包括原油菌解气、晚期生物成因气、煤层菌解气、浅层次生蚀变改造型气[15-16]。前三者是原油、热演化有机质或者煤层在厌氧微生物降解过程中形成的生物气[17-18],后者是原油氧化环境下喜氧细菌作用形成[4]。
1.2 生物成因气分布特征
世界生物气集中分布在墨西哥湾、库克湾和西西伯利亚盆地[19]。西西伯利亚盆地及邻区早在20世纪80年代发现了34个生物气田,是生物气和热解气的混合气,乌连戈伊超大气田就是西西伯利亚盆地的生物气田[20]。加拿大、德国、意大利、哥伦比亚、日本和中国等国家也发现不同规模的生物气藏[21]。中国最集中且已经投入开发的原生生物气田主要分布在柴达木盆地东部三湖地区,目前已经发现涩北一号、涩北二号、台南、伊克雅乌汝、盐湖、驼峰山、台吉乃尔等生物气田[22]。长江沿岸及云南、安徽等小盆地有原生生物气显示,莺—琼盆地多为生物-热催化过渡带气或者生物气与过渡带气的混合,松辽、渤海湾、二连、江汉等盆地多分布过渡带气和原油菌解气,在煤系地层发育的鄂尔多斯、准噶尔盆地多有煤层菌解气分布[23](图1)。
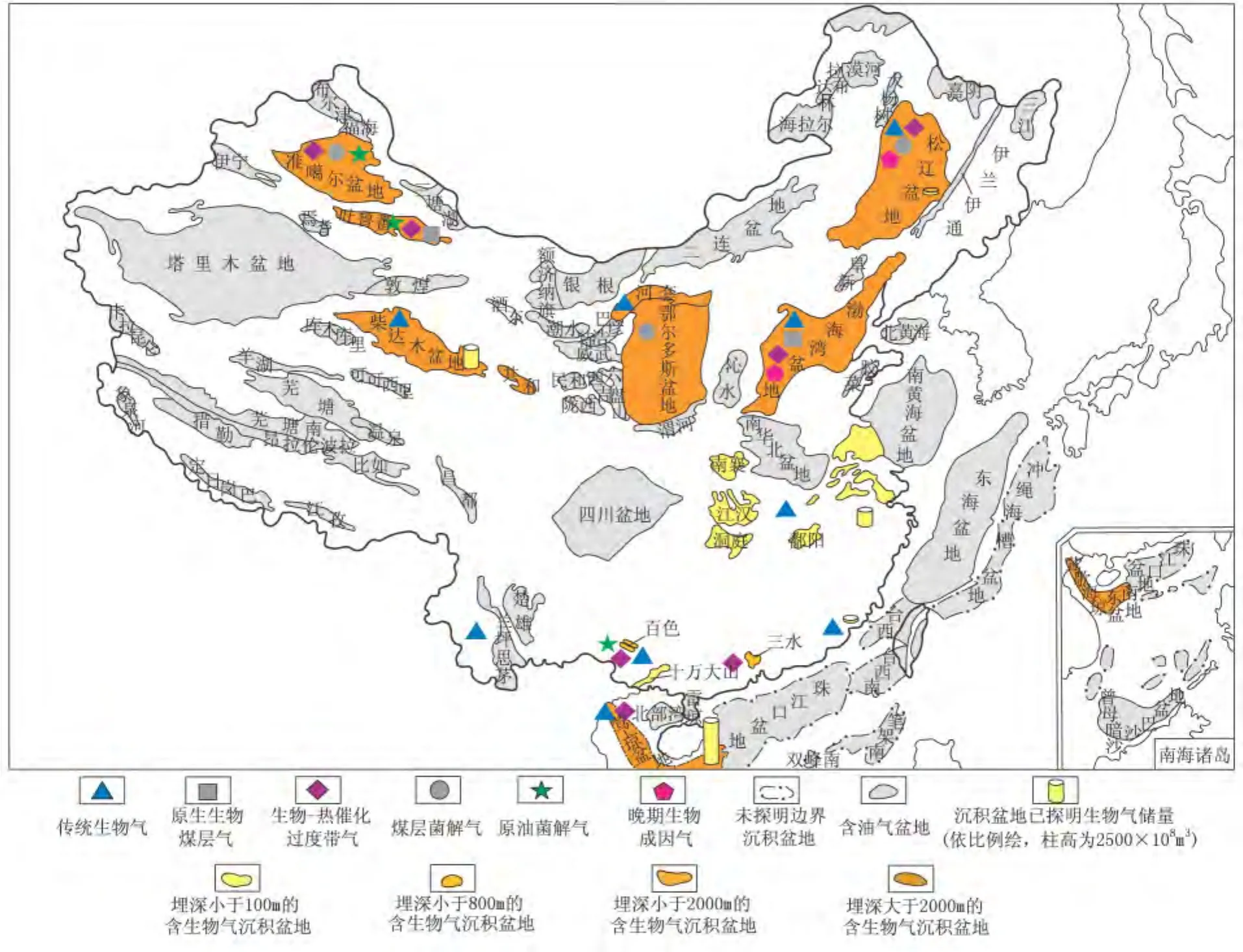
图1 中国主要生物气藏分布
生物气的分布具有时代新、埋藏浅的特征。世界生物气藏多存在于白垩纪、古近纪、新近纪和第四纪的三角洲、大陆架和陆相等环境,埋藏深度几十米到上千米,时代最老的是美国密执安盆地的中—晚泥盆世Antrim页岩和阿巴拉契亚盆地泥盆纪页岩的生物气[19]。中国生物气藏主要集中分布在第四系、新近系和古近系,尤其是第四系,柴达木盆地三湖地区生物气田主要分布于第四系中下更新统七个泉组和新近系上新统狮子沟组[24],深度多在 1700 m 以上[25]。
2 原生型生物气的形成
原生生物气和表生菌解气的生成过程都是在微生物降解作用下,通过CO2还原和乙酸发酵2种途径形成[26-28],只是生气时间不同和主要的厌氧细菌——甲烷菌消化物料存在差异,在成因机制上没有本质的区别[15]。
2.1 沉积地层中的微生物环境
沉积物自上而下形成了喜氧微生物带、硫酸盐还原带和生物甲烷生成带3种不同的微生物环境,各种生化环境都有占优势的微生物群体,这些微生物群体利用各自的环境,创造出有利于不同群体的新环境[27]。喜氧微生物带发育在水体中和沉积物剖面的上部,喜氧微生物利用溶解在水中的氧气进行喜氧呼吸,对有机物进行初步分解,当氧气消耗尽时,沉积环境进入厌氧微生物适宜生存的阶段。硫酸盐还原带只存在于海水和含硫酸盐的海洋沉积环境中,陆相沉积一般不含硫酸盐,当硫酸盐耗尽后,才进入生物甲烷生成带,硫酸盐还原作用和生物产甲烷作用二者呈渐变关系。因此,只有当沉积有机质越过喜氧微生物带及硫酸盐还原带后,才进入生物甲烷生成带,生成大量生物气。
2.2 生物成因气生成过程
生物气是有机质在微生物作用下经矿化生物化学过程的重要终极产物。生物和动植物体埋藏后,首先经过喜氧微生物的耗氧降解,有机物被分解成产甲烷菌能利用的底物,同时由于氧气被喜氧微生物消耗及兼性细菌的过渡,逐步演变为厌氧生态环境。在厌氧生态系统中,从聚合有机物到生物甲烷的形成过程主要分为3个阶段[5](图2)。
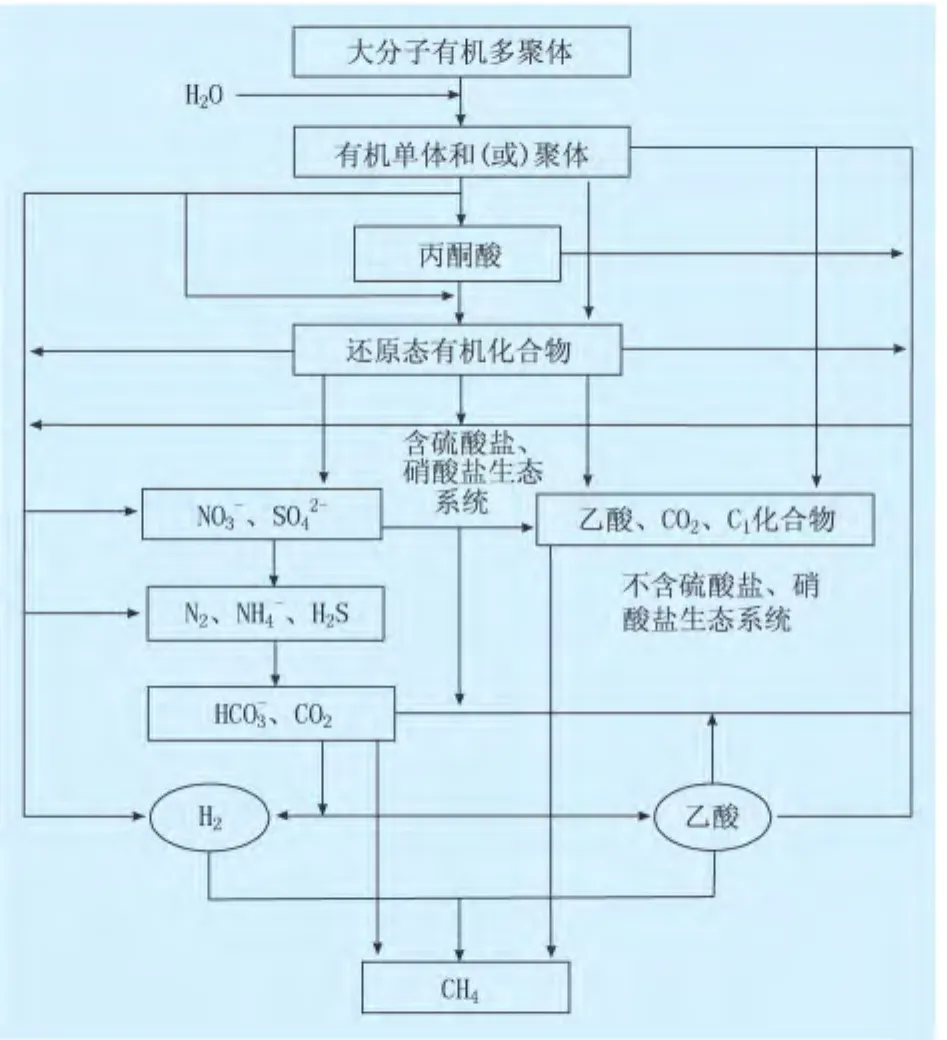
图2 厌氧生态环境中有机质降解生甲烷机理(据文献[29])
(1)第1阶段。有机大分子被发酵菌或水解菌分解为氨基酸、糖、脂肪酸、甘油等有机单体,而后水解为还原态有机化合物的丙酸、丁酸、乳酸和醇类等。
(2)第2阶段。在不含硫酸盐和(或)硝酸盐还原菌的生态环境下,还原态有机化合物被产氢菌和产乙酸菌等降解为乙酸、CO2及甲酸、甲醇、甲胺等和氢。有硫酸盐和(或)硝酸盐存在的地质系统中,还原态有机化合物优先被硫酸盐和(或)硝酸盐还原菌降解形成H2S,并抑制甲烷菌的产甲烷作用,只有硫酸盐还原菌消失后,还原态有机化合物才能进一步降解为CO2和乙酸。
(3)第3阶段。产甲烷菌在辅酶作用下,通过H2+CO2还原和乙酸发酵2种途径转化为甲烷[5]。
上述反应过程决定于沉积物中氧化剂的性质,游离氧分解后以金属氧化物(MnO2和Fe2O3)为主要氧化剂,硝酸盐还原后进入硫酸盐还原带,最后进入产甲烷菌还原带。当沉积物表面处于喜氧带或硫酸盐还原带时,有机质被大量氧化、消耗,不利于生物甲烷生成或导致甲烷过早形成,难以保存;只有当沉积物处于过渡性硫酸盐还原带之下或碳酸盐还原带,且该带厚度较大时,才有利于生物甲烷生成并保存[30]。深部微生物学研究认为,微生物可以栖息在从地表到4000 m或者更深的地方,这将要重新考虑硫酸盐还原带以下微生物甲烷形成的重要性[31-32]。
3 生物成因气形成演化特征
成岩作用阶段早期(生物化学作用阶段)有机质的演化特征,在文献[33-40]有机质成烃研究中有不同程度的描述,但并未进行深入研究。由于不同深度有机质的环境及主要微生物群种的不同,有机质被降解的过程以及形成的产物有明显的区别,使生物成因气源岩的演化过程和生物成因气的生成过程具有明显的阶段性。综合前人研究成果,结合典型生物气盆地生物气形成演化过程剖析,按照生物甲烷的产气率、微生物群落的配置及其活跃性,将生物气生成演化划分为3个阶段。
3.1 生物化学生气早期阶段
由于水体至沉积物表层(0到几十米)存在大量的氧气,有机质首先发生喜氧微生物的耗氧降解,这个过程的长短影响到有机质的保存和后期烃类的形成。有机质可被喜氧细菌的持续作用全部消耗;而当沉积物受到一定深度水体覆盖时,游离氧被消耗殆尽,抑制了喜氧细菌的活动。现代沉积研究结果[41-42]表明,在生物化学作用阶段有50%~80%的有机质被消耗。这个过程同时也创造了有机质被保存的贫氧环境,在厌氧沉积环境中,沉积物从上到下可划分为硫酸盐还原带、过渡带(即甲烷厌氧氧化带)和甲烷形成带3个特征带。硫酸盐还原带中的还原反应消耗过多的氢而抑制了甲烷的形成,过渡带中的氧化还原反应受甲烷形成菌-硫酸盐还原菌综合体影响[43],硫酸盐还原带中的硫酸盐下移到过渡带中,被从甲烷形成带上移的甲烷氧化消耗,甲烷形成带由发酵作用产生的大量氢有利于甲烷形成菌形成甲烷[44]。因此,硫酸盐还原带到甲烷形成带,硫酸盐急剧减少,甲烷迅速增加。生物模拟实验证实,柴达木盆地三湖坳陷0~200 m存在4类微生物菌群,尽管硫酸盐还原菌的存在不利于产甲烷菌的活动,但产甲烷菌仍有一定的降解作用,所形成甲烷在浅部盐层或其他盖层的遮挡下,于较高部位聚集形成局部气藏(图3)[24]。李明宅等[45]在埋深达1400 ~1700 m 的实验样品中仍检测到硫酸盐还原菌,在55℃时该样品中硫酸盐还原菌产H2S率高,抑制产甲烷菌的作用,说明在55℃时不产气;在65、75℃时,抑制作用解除后,才开始产气(表1)。一般情况下,硫酸盐还原菌的作用深度在300 m以浅范围内。
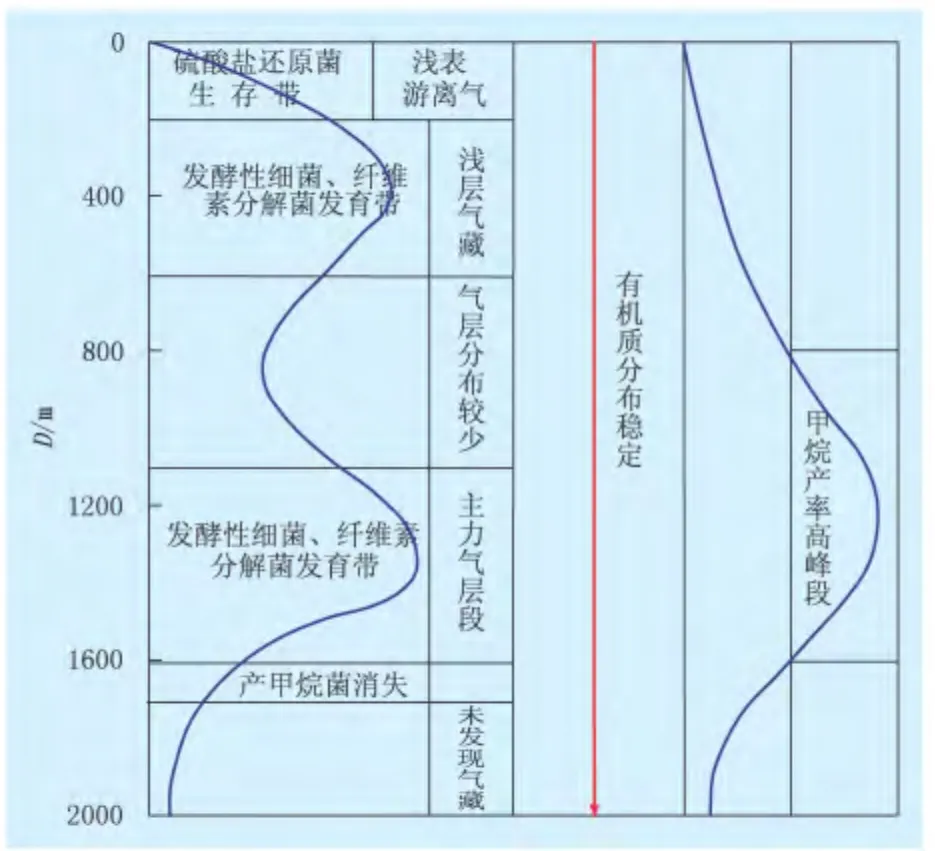
图3 第四系生物气生成演化模式图(据文献[24])
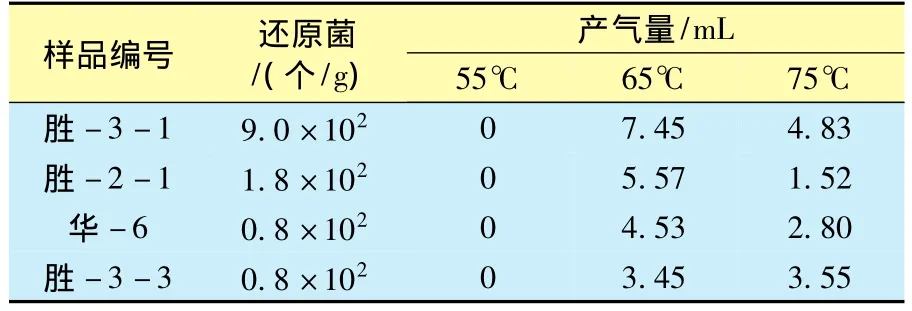
表1 硫酸盐还原菌与产气率关系(据文献[45])
3.2 生物化学生气高峰期阶段
随着沉积物埋深增加和温度升高,硫酸盐还原菌完全消失,环境条件逐渐适宜产甲烷菌群的生存。浅层发酵菌优先利用可溶有机质,发酵分解为还原态有机化合物,然后,在产氢菌和产乙酸菌的作用下分解形成乙酸、二氧化碳和氢等产甲烷菌可利用的最终底物,产甲烷菌不断利用移除底物的同时,促使有机质发酵水解并生成大量甲烷。生物模拟试验表明,在 35、55、65、75℃ 4个温阶,样品的平均产甲烷量分别为 5.65、5.63、3.98、2.55 mL,以35~55℃这一温度段的生气作用占主导地位,为生气高峰阶段[45]。柴达木盆地三湖坳陷深度超过200 m,硫酸盐还原菌消失,温度逐渐升高,该阶段适应的微生物菌群繁盛,微生物分解大量的有机组分,开始形成大量甲烷气;1000~1600 m,地层温度为35~65℃,此阶段适宜于本源细菌的生长,同时有机组分的成熟度增加,可供细菌降解的物质增多,微生物菌群数量增多,甲烷产率也达到最高值。而在600~1000 m深度,由于有机组分、温度、压力等诸多因素的改变,浅层微生物菌群已不适应此环境,微生物菌群数量有所下降,甲烷产率降低(图3)。
3.3 生物化学生气晚期阶段
当埋深增加,温度超过产甲烷菌群生存的极限,加之易被微生物利用的有机质在高峰期阶段已被大量降解消耗,可被微生物发酵利用形成产甲烷菌可利用底物的有机质逐渐减少,营养的缺乏以及高温的影响,产甲烷菌数量急剧减少,活跃性也大大降低,甲烷产率很低且随着深度增加逐渐减小。该阶段,包括木栓质体、树脂体、经细菌改造的陆源有机质、藻类和高等植物生物类脂物以及富硫大分子等一些特殊组成的有机质,通过低温生物化学和低温化学反应生成未熟油和低熟气[46]。柴达木盆地三湖坳陷1600~2000 m微生物菌群数量不断减少,1700 m以下已不存在产甲烷菌,至2000 m纤维素分解菌和发酵性菌消失,只在台南气田分布少量气层[24]。
4 生物气形成演化模式及影响因素
4.1 生物气形成演化模式
随着埋藏深度和温度的增加,生物气的生成演化呈现阶段性的特征(图4)。

图4 生物成因气生成演化模式
(1)生物化学生气早期阶段。具有低温、高硫酸盐的抑制等很多不利于生物气形成的条件,要么直接处于氧化带之下,由于缺乏良好的圈闭和盖层,加之成岩性差,形成的甲烷很容易散失;要么处于硫酸盐还原带,主要发生硫酸盐还原作用,抑制产甲烷菌的活动,只能生成少量的甲烷,使有机质得以保存。该阶段深度可从沉积物表面延伸到200~300 m左右,温度一般低于25℃,生物气产率比较低,在浅层不能形成有效的气藏。
(2)生物化学生气高峰期阶段。有机质埋深大约为300~1600 m,由于不同地区地质环境条件的不同,深度范围可能更宽。该阶段温度约为30~70℃,有机质未成熟,Ro一般小于0.4%。生物气产率和产量的主要控制因素是产甲烷菌和有机质的性质,其他因素如温度、压力、盐度和pH等都是通过影响产甲烷菌而间接影响甲烷的产率。有机质类型决定了微生物可利用的程度,而丰度决定最终的生成量。柴达木盆地虽然整体丰度比较低,但是高的沉积速率形成巨厚源岩厚度弥补了丰度的不足,形成储量巨大的生物气藏。
(3)生物化学生气晚期阶段。埋深超过1700 m,地温达到70℃以上,Ro达到0.4%以上。该阶段由于有低温热催化作用的参与,产物除了生物甲烷以外,还包括一些重烃气。甲烷含量一般达98%以上,C2—C4的重烃含量很少,绝大多数低于0.5%,干燥系数(甲烷含量与重烃总量比值)很高,C1-C3烃类气体成分比值 R通常大于1000[4],为典型的干气。
此演化模式的深度范围,不同的地质和环境条件会有一定的差异,取决于碳酸盐还原带的厚度以及生物菌群类型,柴达木盆地生物气生成下限深度至少达到2000 m,中下部(深度为700~1600 m)地层甲烷产率相对较高,是浅部地层的3~4倍。模拟实验证明,古细菌醇在2000 m的深度范围内仍然存在,说明2000 m并不是产甲烷菌的生存下限[16],只是环境条件已越来越不适合产甲烷菌生存,导致产甲烷菌数量减少,产甲烷量也相应逐渐减少,有机质低温热解催化产气开始起作用。
4.2 生物气形成的影响因素
生物气的形成是微生物新陈代谢的产物,取决于主要细菌产甲烷菌的生存和大量活动,影响甲烷菌生存和活动的根本因素在于提供细菌生存和繁衍的营养源(有机质)及维持较高活性的地质条件[19]。包括可溶有机质在内的丰富有机质[47]及半腐殖型和草本腐殖型有机质类型是形成生物气的物质基础[48-50],持续沉降和快速沉积是保证生物气生成的地质因素。绝对厌氧环境、还原条件(Eh﹤300)、低浓度硫酸根离子、适宜的温度(一般不高于80℃)使产甲烷菌得以大量活动,是生物气形成的前提。同时,较疏松的细粒沉积物中有机质富集且有较大孔隙,有利于细菌的生存和活动[50]。因此,以草本植物为主的母质快速沉积,低温、封闭的缺氧环境以及低的成岩演化阶段,是生物气生成的物质与环境条件;而发酵菌、厌氧菌、产甲烷菌等微生物菌群的发育与繁盛,则是生物气生成的动力条件。
5 结论
(1)按照生气时间和母质以及地质条件的不同,生物成因气可划分为2型7类,包括原生型的传统生物气、煤层生物气、生物-热催化过渡带气和次生型的原油菌解气、煤层菌解气、晚期生物成因气、浅层次生蚀变改造型气。主要分布于新近系和第四系中,埋深大多在2000 m以内。
(2)生物成因气是有机质在厌氧环境中微生物作用下,通过CO2还原和乙酸发酵2种途径形成的最终产物。
(3)有机质被降解的过程以及形成的产物具有明显的阶段性特征。生物化学生气早期,具有低温、高硫酸盐抑制等不利条件,生物气产率比较低;生物化学生气高峰期,发酵菌优先利用可溶有机质分解成产甲烷菌可利用的底物,同时产甲烷菌发酵水解底物生成大量甲烷,以35~55℃为最佳生气温度;生物化学生气晚期,地温达到70℃以上,不适宜产甲烷菌群生存及可利用底物减少,甲烷产率很低。
(4)生物气的形成取决于维持产甲烷菌生存和繁衍可以利用的营养源(有机质)及维持较高活性的地质条件,有机质丰度和类型、高沉积速率等地质环境及微生物发育程度成为影响生物气形成的主要因素。
[1]Claypool G E,Kvenvolden K A.Methane and other hydrocarbon gases in marine sediments[J].Annual Review of Earth and Planetary Sciences,1983,11(1):299 - 327.
[2]Kvenvolden K A.A review of the geochemistry of methane in natural gas hydrate[J].Organic Geochemistry,1995,23(11/12):177-189.
[3]张小军,陶明信,王万春,等.生物成因煤层气的生成及其资源意义[J].矿物岩石地球化学报,2004,23(2):166-171.
[4]李先奇,张水昌,朱光有,等.中国生物成因气的类型划分与研究方向[J].天然气地球科学,2005,16(4):477-483.
[5]丁安娜,王明明,李本亮,等.生物气的形成机理及源岩的地球化学特征——以柴达木盆地生物气为例[J].天然气地球科学,2003,14(5):402 -407.
[6]陈荣书,袁炳存.天然气地质学[M].武汉:中国地质大学出版社,1983:104-116.
[7]包茨.天然气地质学[M].北京:科学出版社,1988:14-24.
[8]戴金星.中国天然气地质学[M].北京:石油工业出版社,1992:6-15.
[9]张英,戴金星,李剑,等.我国生物气的地化特征与勘探方向[J].天然气工业,2009,29(9):20-23.
[10]徐永昌.我国80年代气体地球化学研究[J].沉积学报,1992,10(3):57 -68.
[11]杜尚明,胡光灿,李景明,等.天然气资源勘探[M].北京:石油工业出版社,2004:16-32.
[12]Scott A R,Kaise W R,Ayers W B Jr.Thermogenic and secondary biogenic gases,San Juan Basin,Colorado and New Mexico-implications for coal bed gas producibility[J].AAPG Bulletin,1994,78(8):1186 -1209.
[13]徐永昌.天然气成因理论及应用[M].北京:科学出版社,1994:102-118.
[14]邵珠福,钟建华,于艳玲,等.从成藏条件和成藏机理对比非常规页岩气和煤层气[J].特种油气藏,2012,19(4):21-24.
[15]李赞豪.具有广阔勘探前景的一种新型浅层天然气——油层、煤层厌氧菌解再生生物气[J].石油实验地质,1994,16(3):220 -229.
[16]帅燕华,张永昌,苏爱国,等.生物成因天然气勘探前景初步分析[J].天然气工业,2006,26(8):1-4.
[17]孙俊民.煤层气的成因及地球化学特征[J].焦作工学院学报,1998,17(4):245 -248.
[18]李晶莹,陶明信.国际煤层气组成和成因研究[J].地球科学进展,1998,13(5)467 -473.
[19]林春明,李艳丽,漆滨汶.生物气研究现状与勘探前景[J].古地理学报,2006,8(3):317-330.
[20]何家雄,夏斌,张启明,等.南海北部边缘盆地生物气和亚生物气资源潜力与勘探前景分析[J].天然气地球科学,2005,16(2):167 -174.
[21]贾承造,赵文智,魏国齐,等.国外天然气勘探与研究最新进展及发展趋势[J].天然气工业,2002,22(4):5-9.
[22]关洪军,朱大奎,张道伟.柴达木盆地三湖地区第四系地下水流场与生物气藏成因分析[J].地质论评,2007,53(4):491 -497.
[23]李景明,魏国齐,陈孟晋.中国天然气地质理论研究新进展[J].中国石油勘探,2002,7(1):8-16.
[24]梅建森,康毅力,张永高,等.柴达木盆地生物气源岩评价及勘探方向[J].天然气工业,2007,27(9):17-20.
[25]魏国齐,刘德来,张英,等.柴达木盆地第四系生物气形成机理、分布规律与勘探前景[J].石油勘探与开发,2005,32(4):84 -89.
[26]陈英,戴金星,戚厚发.关于生物气研究中几个理论及方法问题的研究[J].石油实验地质,1994,16(3):209-219.
[27]关德师,戚厚发,钱贻伯,等.生物气的生成演化模式[J]. 石油学报,1997,18(3):31 -36.
[28]史占祯.渤海湾盆地及其外围的生物气研究[J].天然气工业,2002,22(5):11 -16.
[29]Zehnder A J B.Biology of anaerobic micro organisms[M].New York:John Wiley and Sons,1988:2 -5.
[30]黄保家,肖贤明.莺歌海盆地海相生物气特征及生化成气模式[J].沉积学报,2002,20(3):462-468.
[31]张水昌,赵文智,李先奇,等.生物气研究新进展与勘探策略[J].石油勘探与开发,2005,32(4):90-96.
[32]Ward J A,Slater G F,Moser D P,et al.Microbial hydrocarbongases in the Witwatersrand Basin,South Africa:implications for the deep biosphere [J].Geochimica et Cosmochimica Acta,2004,68(15):3239 -3250.
[33]Hilary Irqwin,et al.Isotopic evidence for source of diagenetic carbonates formed during burial of organic-rich sediments[J].Nature,1977,269(5265):209 - 213.
[34]Tissot B P,Welte D H.Petroleum formation and occurrence[M].Berlin,DEU:Springer- Verlag,1978:160 -253.
[35]Rice D D,Claypool G E.Generation,accumulation and resource potential of biogenic gas[J].AAPG,1981,65(1):5-25.
[36]张义纲,陈焕疆.论生物气的生成和聚集[J].石油与天然气地质,1983,4(2):160 -170.
[37]维索茨基.天然气地质学[M].戴金星,等译.北京:石油工业出版社,1986:153-173.
[38]王大锐,宋岩.碳同位素在生物气勘探中的示踪作用[J].石油勘探与开发,1992,19(4):47 -51.
[39]姜乃煌,宋孚庆,任冬芬,等.甲烷菌发酵阶段划分[J]. 石油勘探与开发,1993,20(4):39-43.
[40]李明宅,张洪年,郜建军.生物气的生成演化模式和初次运移特征[J].石油实验地质,1995,17(2),147-155.
[41]黄第藩,等.陆相有机质演化和成烃机理[M].北京:石油工业出版社,1984:109.
[42]中国科学院兰州地质研究所,等.青海湖综合考察报告[R].北京:科学出版社,1979:257-259.
[43]Hoehler T M,Alperin M J,Daniel B A,et al.Field and laboratory studies of methane oxidation in anoxic marine sediment:evidence for a methanogen-sulfate reducer consortium[J].Global Biogeochemical cycles,1994,8(4):451-463.
[44]Kuivila K M,Murray J W,Devol A H.Methane production,sulfate reduction and competition for substrates in the sediments of Lake Washington[J].Geochimica et Cosmochimica acta,1989,53(2):409 -416.
[45]李明宅,张洪年,刘华,等.生物气模拟试验的进展[J]. 石油与天然气地质,1996,17(2):117-122.
[46]王铁冠,钟宁宁,侯读杰,等.中国低熟油的几种成因机制[J].沉积学报,1997,15(2):75-83.
[47]张英,李剑,张奎,等.柴达木盆地三湖地区第四系生物气源岩中可溶有机质丰度及地质意义[J].地质学报,2007,81(12):1716 -1722.
[48]张英,王晓波,李谨,等.不同类型有机质生物产甲烷模拟实验研究[J].石油实验地质,2009,31(6):633-636.
[49]康晏,王万春,任军虎.生物气生成的地球化学因素分析[J].矿物岩石地球化学通报,2004,23(4):350-354.
[50]关德师.控制生物气富集成藏的基本地质因素[J].天然气工业,1997,17(5):8 -12.
