高脂膳食和跑台运动对雄性大鼠腓肠肌腺苷酸活化蛋白激酶 /乙酰辅酶A羧化酶信号通路和膜蛋白脂肪酸转位酶蛋白含量的影响
2015-10-15张云丽娄淑杰
张云丽, 娄淑杰
(1. 上海体育学院 运动科学学院, 上海 200438; 2. 聊城大学体育学院, 聊城 252059)
高脂膳食和跑台运动对雄性大鼠腓肠肌腺苷酸活化蛋白激酶 /乙酰辅酶A羧化酶信号通路和膜蛋白脂肪酸转位酶蛋白含量的影响
张云丽1,2, 娄淑杰1
(1. 上海体育学院 运动科学学院, 上海 200438; 2. 聊城大学体育学院, 聊城 252059)
目的 探讨高脂膳食和8周有氧耐力运动对大鼠腓肠肌腺苷酸活化蛋白激酶(AMPK)/乙酰辅酶A羧化酶(ACC)信号通路和膜蛋白脂肪酸转位酶CD36含量的影响。方法 建立营养性肥胖大鼠模型,并随机分为肥胖安静组(OC组)和肥胖运动组(OE组),另设普通饲料安静组(NC组)和普通饲料运动组(NE组)。运动干预结束后检测腓肠肌AMPKα、p-AMPKα、ACC、p-ACC和膜蛋白CD36的蛋白水平。结果 1) NE组与NC、OE及OC组相比,p-AMPKα的蛋白水平显著升高(P<0.01); OC组p-AMPKα的蛋白水平显著低于NC组(P<0.01)。2) OC组p-ACC蛋白水平显著低于NC组(P<0.01); OE组p-ACC的蛋白水平显著高于OC组(P<0.01)。3) NE组与NC、OE及OC组相比,OC组与NC组相比,膜蛋白CD36含量均无显著性变化(P>0.05)。结论 1) 运动可改善高脂膳食引起的AMPK/ACC信号通路障碍。2) 运动对体质量不同大鼠p-ACC蛋白水平的影响存在差异。3) 腓肠肌膜蛋白CD36的含量并没有伴随AMPK/ACC信号通路的激活或抑制而发生显著变化。
跑台运动; 高脂膳食; 肥胖; 腺苷酸活化蛋白激酶(AMPK); 乙酰辅酶A羧化酶(ACC);脂肪酸转位酶CD36
腺苷酸活化蛋白激酶(AMPK)是调节细胞能量平衡的关键酶,激活AMPK将导致乙酰辅酶A羧化酶(ACC)受抑制,随后通过促进脂肪酸氧化、葡萄糖摄取等过程并抑制脂肪合成、蛋白质合成以及糖异生等途径来调节机体的能量平衡[1-3]。既往研究表明[4], 脂肪酸转位酶CD36广泛存在于脂肪酸代谢活跃的组织, 与脂肪酸的跨膜转运有关, AMPK/ACC信号通路和CD36均与骨骼肌脂肪酸氧化代谢相关,在肥胖的形成中具重要作用,但高脂膳食和运动对AMPK/ACC信号通路以及CD36的影响尚无定论。从检索到的文献看,目前有关高脂膳食和运动对AMPK/ACC信号通路和CD36蛋白含量影响的相关报道逐渐增多,但是由于动物模型、运动方案、组织样本选择以及检测亚基类型等的不同,造成研究结果并不一致,并且CD36蛋白含量与AMPK/ACC信号通路激活之间的关系尚不明确。作者前期研究表明[5],同一强度运动对相同性别、相同年龄的体质量正常大鼠和肥胖大鼠的脂肪组织量以及血清TG水平的影响存在差异性,那么这种差异性是否与AMPK/ACC信号通路以及CD36蛋白含量的不同有关?
针对上述问题, 本研究以普通饮食的体质量正常和高脂饮食诱导的肥胖大鼠为研究对象, 探讨高脂膳食和8周有氧耐力运动对大鼠腓肠肌AMPK/ACC信号通路以及膜蛋白CD36含量的影响。
1 材料与方法
1.1实验动物
7周龄清洁级雄性SD大鼠70只, 体质量190± 10 g,购自上海西普尔-必凯实验动物有限公司[SCXK(沪)2013-0016]。
1.2主要试剂及仪器
BCA蛋白浓度测定试剂盒(增强型)、BeyoECL Plus (超敏ECL化学发光试剂盒)及PVDF膜购于碧云天生物试剂公司; 兔单克隆抗体AM PK α、pAMPKα-Thr172, 兔多克隆抗体ACC、pACC-Ser79及辣根酶标记的羊抗兔IgG二抗、彩色预染蛋白质分子量标准均购于 Cell Signaling公司;兔多克隆抗体CD36购于Santa Cruz公司;兔多克隆抗体GAPDH购于杭州贤至生物科技有限公司;膜蛋白,核蛋白和胞质蛋白抽提试剂盒购于生工生物工程(上海)有限公司; 蛋白质电泳仪、蛋白质转移装置购于美国Biorad公司。
1.3模型构建及分组
普通饲料适应性喂养后,随机分为普通饲料组(n=20,C组)和高脂饲料组(n=50,H组)。正式实验第8周末,将普通饲料组大鼠随机分为普通饲料安静组(n=9,NC组,1只在实验过程中因病死亡)和普通饲料运动组(n=10,NE组); 选取高脂饲料组中体质量超过普通饲料组大鼠平均体质量加1.4倍标准差的大鼠, 并随机分为肥胖安静组(n=10,OC组)和肥胖运动组(n=8,OE组,2只不能完成运动被淘汰),继续高脂饲料喂养直至实验结束, 其余淘汰。基础饲料及高脂饲料均购自上海斯莱克实验动物有限责任公司(表1)。大鼠饲养于清洁级设施[SYXK(沪)2012-0003],每笼3~5只,自由摄食和饮水,动物房温度22±2℃,相对湿度50%± 10%,光照遵循12 h∶12 h明暗周期。
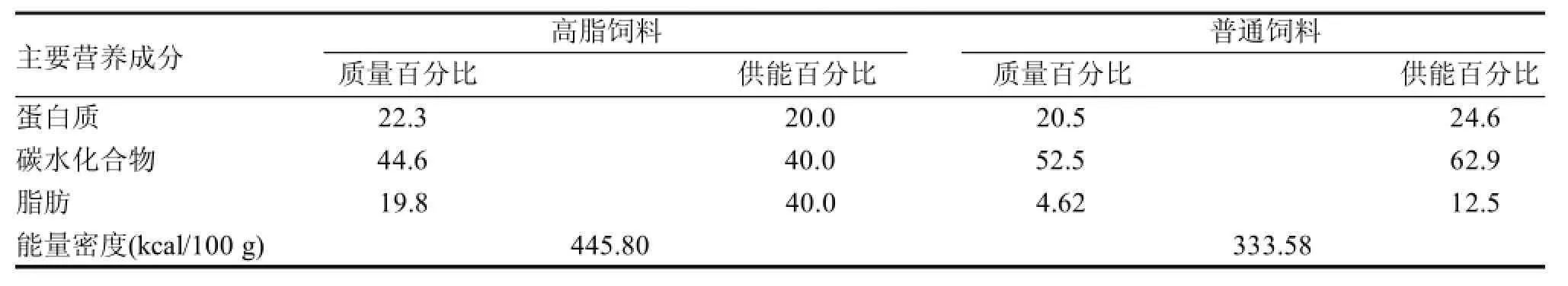
表1 饲料主要营养成分构成 %
1.3体质量监测
整个实验过程中,每周六上午固定时间称量大鼠体质量(第17周除外)。
1.4运动方案
NC组和OC组不运动。NE组和OE组经1周适应性运动后进行8周坡度0°跑台运动, 1 次/日,5 次/周。运动负荷安排如下: 按11 m/min, 10 min/d开始运动, 之后每日以1 m/min增加运动速度, 5 min/d增加运动时间, 直至跑速为18 m/min,运动时间为40 min/d。然后维持跑速为18 m/min,运动时间为40 min/d,直至实验结束。
1.5标本采集与储存
末次运动结束后24 h,禁食过夜,每组大鼠中随机选取 6只,常规麻醉,断头处死,迅速分离左侧腓肠肌,冻存于液氮中,后转至-80℃冰箱保存待测。
1.6Western blotting法检测腓肠肌蛋白表达
称取腓肠肌组织100 mg,用蛋白抽提试剂盒依次提取胞质蛋白(用于检测AMPKα、p-AMPKα-Thr172、ACC、p-ACC-Ser79)和膜蛋白(用于检测CD36)。BCA法检测样品蛋白浓度,用裂解液将各组样品的蛋白浓度调平后,加入相应量蛋白上样缓冲液(5×),沸煮10 min,进行SDS-PAGE电泳,后转移至 PVDF膜上。然后用5%的脱脂牛奶室温摇床封闭1 h,接着依次与相应一抗4 ℃孵育过夜。TBST洗膜3次,每次5 min。室温孵育二抗1 h后,TBST洗膜3次,每次5 min,后用ECL化学发光剂在X线下曝光、显影。Gel Doc EZ凝胶成像仪将X光片扫描成图像并保存为电脑文件后,用Image J软件进行图像分析。
1.7数据统计
2 结果
2.1实验动物数量分析
第8周末分组前共纳入大鼠70只,第8周末分组时共纳入大鼠40只,后续实验中1只因病死亡,2只不能完成运动被淘汰。37只大鼠进入体质量结果的分析。运动干预结束后随机选取24只大鼠进行蛋白表达的结果分析。
2.2不同组别大鼠体质量变化
运动干预前, 高脂膳食组大鼠的体质量自第2 周起就始终显著高于普通饲料组大鼠(P<0.01), 表明高脂膳食可引起大鼠肥胖。运动干预过程中, 正常运动组大鼠的体质量在第14周、15周、16周出现显著下降(P<0.01、P<0.05、P<0.05), 而肥胖运动组大鼠的体质量自第10周起就始终显著低于肥胖对照组大鼠(P<0.01)(图1)。提示运动干预引起正常大鼠体质量适度下降, 但显著降低肥胖大鼠的体质量。
2.3不同组别大鼠骨骼肌AMPKα、p-AMPKα-Thr172、ACC和p-ACC-Ser79蛋白表达
肥胖大鼠骨骼肌AMPKα和ACC含量均无显著性变化(P>0.05, P>0.05), p-AMPKα-Thr172和p-ACC-Ser79表达均明显降低(P<0.01, P<0.01); 运动干预对正常和肥胖大鼠骨骼肌AMPKα和ACC含量均无显著性影响(P>0.05, P>0.05), 但正常和肥胖大鼠骨骼肌p-AMPKα-Thr172的含量明显升高(P<0.01), 对正常大鼠骨骼肌p-ACC-Ser79含量无明显影响, 但肥胖大鼠骨骼肌p-ACC-Ser79的含量明显升高(P<0.01)(图2)。
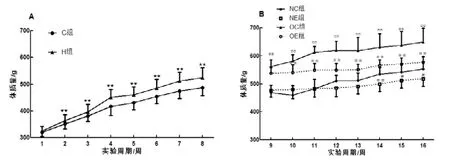
图1 各组大鼠体质量变化曲线
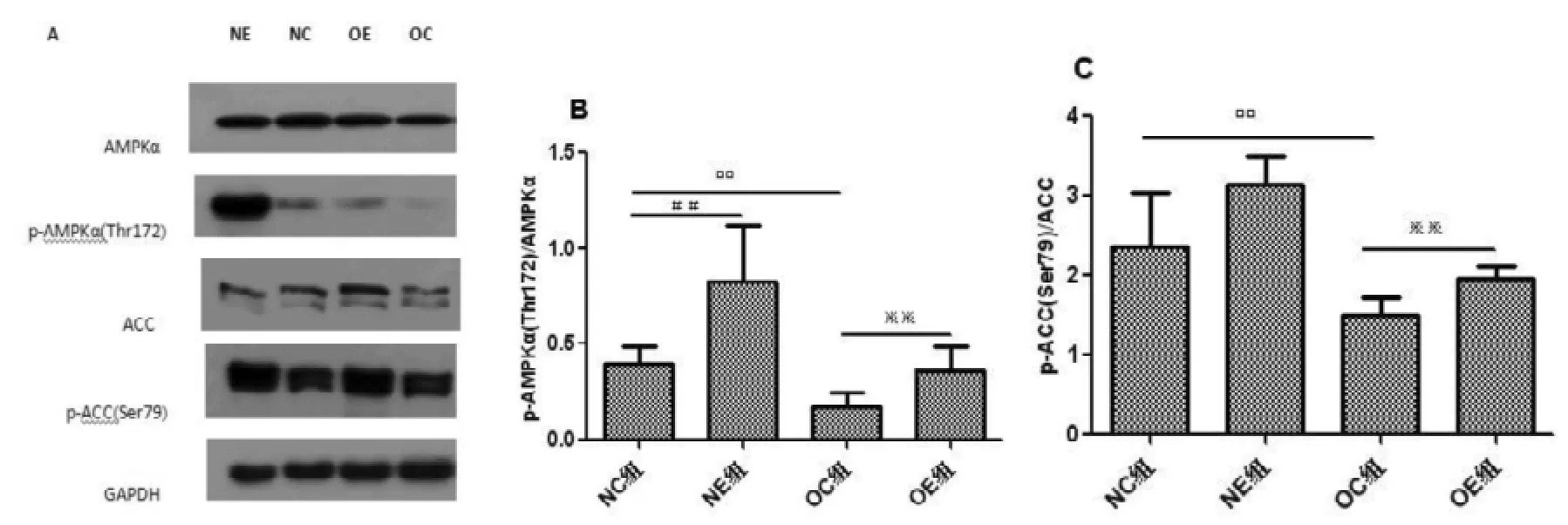
图2 不同组别大鼠腓肠肌A-MPKα、p-AMPKα-Thr172、ACC和p-ACC-Ser79蛋白表达变化情况
2.4不同组别大鼠腓肠肌膜蛋白CD36表达
肥胖大鼠骨骼肌膜蛋白CD36表达无显著性变化(P>0.05); 运动干预对正常大鼠和肥胖大鼠骨骼肌膜蛋白CD36表达均无显著性影响(P>0.05, P>0.05)(图3)。
3 讨论
本研究采用高脂膳食构建的肥胖大鼠体质量明显高于普通膳食大鼠,成功建立肥胖大鼠模型。
3.1肥胖大鼠腓肠肌AMPK/ACC信号通路障碍
在哺乳动物中, AMPK的活性与细胞能量水平密切相关,AMPK的下游蛋白ACC是脂肪酸代谢的限速酶,在脂肪酸的分解或者合成代谢过程中起着重要作用。AMPK和ACC的活性均受营养状态的调节。有研究显示[6], 长期高脂膳食诱导的肥胖或胰岛素抵抗大鼠骨骼肌中AMPK总蛋白含量及磷酸化水平均有显著的减少。Liu等[7]研究表明, 5个月的高脂膳食诱导大鼠骨骼肌AMPKα蛋白含量、p-AMPKα蛋白水平以及AMPKα 2 mRNA水平显著降低, 此外p-ACC的蛋白水平和p-ACC mRNA水平亦显著降低。但孙婧瑜等[8]研究显示, 8周高脂膳食对小鼠骨骼肌AMPK和ACC的蛋白磷酸化水平没有显著影响。导致不同研究结果的原因可能与高脂膳食的构成和喂养周期以及实验动物的种属差异有关。
与Liu等[7]研究相一致,本研究中长期高脂膳食引起p-AMPKα和p-ACC的蛋白水平均出现显著下降(P<0.01),说明高脂膳食可以降低AMPK/ACC信号通路的激活,引起骨骼肌ACC活性增强,进而使骨骼肌脂肪合成作用增强而脂肪酸氧化作用受到抑制,这与高脂膳食引起大鼠体质量增加,进而导致肥胖有关。
3.2运动对大鼠腓肠肌AMPK/ACC信号通路影响
当运动引起机体能量消耗增加时,细胞内AMPK的活性增加,使ACC磷酸化增加而活性降低,进而减少脂肪酸合成并提高脂肪酸的氧化速度,以适应运动对能量增加的需求[9]。孙婧瑜等[8]研究表明,8周自主跑轮运动的小鼠,其骨骼肌AMPK的蛋白磷酸化水平显著增加, 同时ACC的蛋白磷酸化水平也伴随AMPK的激活而显著增加。本研究结果显示, 不论是高脂饲料组还是普通饲料组,8周中等强度的有氧耐力运动均可引起p-AMPKα的蛋白水平增加,但与以往研究不同的是, 本研究显示运动对体质量不同大鼠p-ACC蛋白水平的影响存在差异, 8周中等强度的有氧耐力运动引起肥胖大鼠p-ACC的蛋白水平显著增加(P<0.01), 但对体质量正常大鼠的p-ACC蛋白水平却无显著影响(P>0.05)。本研究选取SD大鼠作为研究对象,采用被动跑台运动方式,而孙婧瑜等[8]以C57BL小鼠为研究对象,采取自主转轮运动方式。实验动物和运动方式不同可能是导致研究结果出现差异的原因。作者以往研究表明[5],8周中等强度的有氧耐力运动可有效减少肥胖大鼠的脂肪组织量,但对体质量正常大鼠无明显影响,其原因可能与运动对肥胖大鼠和体质量正常大鼠的p-ACC蛋白水平的影响不同,进而造成脂肪酸氧化水平的差异所致。但是,运动对体质量不同大鼠p-ACC蛋白水平产生差异性影响的深层次原因仍有待进一步探讨。
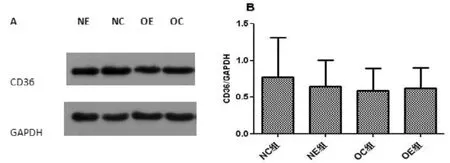
图3 不同组别大鼠腓肠肌膜蛋白CD36表达
3.3肥胖大鼠腓肠肌膜蛋白CD36表达
人类和动物模型的研究表明, 疾病(例如肥胖的进展, 胰岛素抵抗和II型糖尿病)的发病机理与脂肪酸转运体有关联。脂肪酸的跨膜转运涉及多种膜蛋白, 其中CD36最受研究者的关注。Cameron-Smith等[10]研究表明, 短期高脂膳食能够增加受试者脂代谢水平及骨骼肌CD36的总蛋白含量。而孙婧瑜等[8]研究表明,8周的高脂膳食导致骨骼肌CD36总蛋白表达水平显著下降, 分析原因可能与高浓度的脂肪酸引起骨骼肌CD36蛋白泛素化水平增加有关[11]。可见,高脂膳食对骨骼肌CD36总蛋白的影响尚无定论,甚至存在相反的研究结果。
Ring等[12]研究表明,胞质内的CD36不能有效转运脂肪酸。Bonen等[13]通过对肥胖、超重、偏瘦及Ⅱ型糖尿病人群进行调查,显示4组人群骨骼肌CD36总蛋白含量并无差异性,但是肥胖和Ⅱ型糖尿病人群骨骼肌细胞膜上FAT/CD36含量较其他两组均有差异性。但与上述研究不同,本研究结果显示,长期高脂膳食对大鼠腓肠肌膜蛋白CD36的表达水平并无显著性影响(P>0.05)。有研究表明,除质膜外,CD36还可能存在于细胞内的囊泡膜和线粒体膜上[14,15],而本研究所使用的蛋白抽提试剂盒只能提取总膜蛋白,无法获得纯化的质膜蛋白和线粒体膜蛋白,这可能是导致长期高脂膳食对腓肠肌膜蛋白CD36的表达水平无显著性影响的一个原因,提示在某些实验条件下仅仅分离胞质蛋白和膜蛋白是不够的,在今后的研究中需要进一步改进实验条件,以确定高脂膳食对质膜和/或线粒体膜CD36含量的影响。
3.4运动对大鼠腓肠肌膜蛋白CD36表达的影响
Schenk等[16]研究表明, 耐力训练可以增加骨骼肌CD36的总蛋白表达,从而增加脂肪酸的氧化代谢,而Kiens等[17]通过对有两年以上耐力训练经历的受试者进行检测,却发现长期的耐力训练并不能提高股外侧肌CD36的总蛋白表达水平。Chabowski等[18]研究表明, AMPK的激动剂AICAR在激活心肌细胞 AMPK的同时, CD36的基因及蛋白表达水平均出现显著增加。本研究结果显示, 8周中等强度的有氧耐力跑台运动对普通饲料和高脂饲料大鼠腓肠肌膜蛋白CD36含量均无显著性影响(P>0.05), 提示运动在激活AMPK/ACC信号通路的同时并没有增加膜蛋白CD36的含量。那么这是否意味着8周中等强度的有氧耐力跑台运动一定不会改变骨骼肌细胞的脂肪酸摄取呢?首先, 骨骼肌组织中多种蛋白被证明具有脂肪酸转运促进作用, 比如CD36、脂肪酸结合蛋白(FABP)和脂肪酸转运蛋白(FATP等)[19],因此8周中等强度的有氧耐力运动能否影响其他转运体是未知的。其次,被动扩散和蛋白介导的脂肪酸运输可能均有助于细胞的脂肪酸摄取。有研究者指出[20],脂肪酸以扩散形式进行跨膜转运可能是独立发生的或是蛋白介导脂肪酸转运过程一部分,膜脂质双层的特性和脂肪酸结合膜蛋白均会影响细胞对脂肪酸的摄取。这也许可以解释为什么在某些细胞系脂肪酸转运体CD36的增加并不伴有脂肪酸摄取的增加[21,22]。可见,无论是高脂膳食还是运动对CD36的影响及其机制的研究仍需要大量的工作进行验证。
本研究提示,8周中等强度的有氧耐力运动可改善高脂膳食引起的大鼠腓肠肌AMPK/ACC信号通路障碍,并且相同强度的有氧耐力运动对相同性别和月龄的体质量正常和肥胖大鼠腓肠肌p-ACC的蛋白水平的影响存在差异性。此外, 本研究并没有发现大鼠腓肠肌膜蛋白CD36的含量伴随AMPK/ACC信号通路的激活或抑制而发生显著变化。在以后的研究中,将进一步确定高脂膳食和有氧耐力运动对质膜和/或线粒体膜CD36含量的影响。
[1]Carling D. The AMP-activated protein kinase cascade-a unifying system for energy control[J]. Trends Biochem Sci,2004, 29(1):18-24.
[2]Kahn BB, Alquier T, Carling D, et al. AMP-activated protein kinase: ancient energy gauge provides clues to modern understanding of metabolism[J]. Cell Metab, 2005, 1(1):15-25.
[3]Long YC, Zierath JR. AMP-activated protein kinase signaling in metabolic regulation[J]. J Clin Invest, 2006, 116(7):1776-1783.
[4]Holloway GP, Luiken JJ, Glatz JF, et al. Contribution of FAT/ CD36 to the regulation of skeletal muscle fatty acid oxidation:an overview[J]. Acta Physiol (Oxf), 2008, 194(4):293-309.
[5]张云丽, 蔡明, 李静静, 等. 8周有氧耐力运动对正常和肥胖大鼠摄食量、身体成分和血脂的影响[J]. 实验动物与比较医学, 2014, 34(4):294-298.
[6]Lessard SJ, Rivas DA, Chen ZP, et al. Tissue-specific effects of rosiglitazone and exercise in the treatment of lipid-induced insulin resistance[J]. Diabetes, 2007, 56(7):1856-1864.
[7]Liu Y, Wan Q, Guan Q, et al. High-fat diet feeding impairs both the expression and activity of AMPKα in rats’ skeletal muscle[J]. Biochem Biophys Res Commun, 2006, 339(2):701-707.
[8]孙婧瑜, 孙易, 丁树哲. 不同干预方式对小鼠骨骼肌AMPK/ ACC信号通路及CD36蛋白含量的影响[J]. 沈阳体育学院学报, 2014, 33(2):74-79.
[9]Cao S, Li B, Yi X, et al. Effects of exercise on AMPK signaling and downstream components to PI3K in rat with type 2 diabetes[J]. PLoS One, 2012, 7(12):e51709.
[10] Cameron-Smith D, Burke LM, Angus DJ, et al. A short-term,high-fat diet up-regulates lipid metabolism and gene expression in human skeletal muscles[J]. Am J Clin Nutr, 2003, 77 (2):313-318.
[11] Smith J, Su X, El-Maghrabi R, et al.Opposite regulation of CD36 ubiquitination by fatty acids and insulin: effects on fatty acid uptake[J]. J Biol Chem, 2008, 283(20):13578-13585.
[12] Ring A, Le Lay S, Pohl J, et al. Caveolin-1 is required for fatty acid translocase (FAT/CD36) localization and function at the plasma membrane of mouseembryonic fibroblasts[J]. Biochim Biophys Acta, 2006, 1761(4):416-423.
[13] Bonen A, Parolin ML, Steinberg GR, et al. Triacylglycerol accumulation in human obesity and type 2 diabetes is associated with increased rates of skeletal muscle fatty acid transport and increased sarcolemmal FAT/CD36[J]. FASEB J,2004, 18(10):1144-1146.
[14] Keizer HA, Schaart G, Tandon NN, et al. Subcellular immunolocalisation of fatty acid translocase (FAT)/CD36 in human type-1 and type-2 skeletal muscle fibres[J]. Histochem Cell Biol, 2004, 121 (2):101-107.
[15] SebastiánD, Guitart M, García-Mart ínez C, et al. Novel role of FATP1 in mitochondrial fatty acid oxidation in skeletal muscle cells[J]. J Lipid Res, 2009, 50 (9):1789-1799.
[16] Schenk S, Horowitz JF. Coimmunoprecipitation of FAT/ CD36 and CPT1 in skeletal muscle increases proportionally with fat oxidation after endurance exercise training[J]. Am J Physiol Endocrinol Metab, 2006, 291(2):E254-E260.
[17] Kiens B, Roepstorff C, Glatz JF, et al. Lipid-binding proteins and lipoprotein lipase activity in human skeletal muscle:influence of physical activity and gender[J]. J Appl Physiol,2004, 97(4):1209-1218.
[18] Chabowski A, Momken I, Coort SL, et al. Prolonged AMPK activation increases the expression of fatty acid transporters in cardiac myocytes and perfusedhearts [J]. Mol Cell Biochem, 2006, 288(1-2):201-212.
[19] 周丽华, 束刚, 朱晓彤, 等. 肌肉脂肪酸转运膜蛋白及其相互关系[J].中国生物化学与分子生物学报, 2008, 24(11):992-1000.
[20] Luiken JJ, Koonen DP, Coumans WA, et al. Long-chain fatty acid uptake by skeletal muscles is impaired in homozygous,but not heterozygous, heart-type-FABP null mice[J]. Lipids,2003, 38(4):491-496.
[21] Eyre NS, Cleland LG, Mayrhofer G. FAT/CD36 expression alone is insufficient to enhance cellular uptake of oleate[J]. Biochem Biophys Res Commun, 2008, 370(3):404-409.
[22] Van Oort MM, van Doorn JM, Bonen A, et al. Insulin-induced translocation of CD36 to the plasma membrane is reversible and shows similarity to that of GLUT4[J]. Biochim Biophys Acta, 2008, 1781(1-2):61-71.
Effects of High-fat Diet and Treadmill Exercise on AMP-activated Protein Kinase (AMPK)/Acetyl-CoA Carboxylase (ACC) Signaling Pathway and Fatty Acid Translocase CD36 Protein Content in Rat Gastrocnemius Muscle
ZHANG Yun-li1,2, LOU Shu-jie1
(1. Shanghai University of Sport, Shanghai 200438, China;2. Liaocheng University, Liaocheng 252059, China)
Objective To investigate the effects of high-fat diet and 8 weeks aerobic endurance exercise (treadmill exercise) on AMP-activated protein kinase (AMPK)/acetyl-CoA carboxylase (ACC)signaling pathway and fatty acid translocase CD36 protein content in rat gastrocnemius muscles. Methods A rat model of nutritional obesity was established by feeding rats a high-fat diet, and then the obese rats were randomly divided into obese control group (OC group) and obese exercise group (OE group); In addition, a normal diet control group (NC group) and a normal diet exercise group (NE group)were set up. The exercise groups (OE and NE groups) were arranged by aerobic endurance exercise for 8 weeks. After the experiment, the levels of AMPKα, p-AMPKα, ACC, p-ACC and membrane protein CD36 in all groups were detected by Western blotting in gastrocnemius muscles. Results 1) Compared with the NC group, p-AMPKα protein levels had a significant increase in the NE group (P<0.01). p-AMPKα protein levels in the OE group were significantly higher than that in the OC group (P<0.01). p-AMPKα protein levels in the OC group were significantly lower than that in the NC group (P<0.01). 2) Compared with the NC group, p-ACC protein level in the OC group were significantly decreased (P<0.01). p-ACC protein levels had a significant increase in the OE group as compared with the OC group (P<0.01). 3) Compared between the NE group and NC group, the OE group and OC group, and the OC group and NC group, the content of membrane protein CD36 showed no significant change (P>0.05). Conclusion 1) The treadmill exercise could improve AMPK/ACC signaling pathway. 2) The impacts of exercise on p-ACC levels were different in rats with different body mass. 3) The membrane protein CD36 content in the rat gastrocnemius muscles did not significantly change whether AMPK/ACC signaling pathway was activated or inhibited.
Treadmill exercise; High-fat diet; Obesity; AMP-activated protein kinase(AMPK);Acetyl-CoA carboxylase (ACC); Fatty acid translocase CD36
Q95-33
A
1674-5817(2015)06-0441-07
10.3969/j.issn.1674-5817.2015.06.003
2015-07-25
国家自然科学基金(81572241); 上海市人类运动能力开发与保障重点实验室(上海体育学院)(NO.11DZ2261100); 上海体育学院研究生教育创新基金资助项目(xsxr2013030)
张云丽(1977-), 女, 博士研究生。E-mail: ylzhwl@foxmail.com
娄淑杰(1963-), 女, 教授, 博士研究生导师。E-mail: shujielou319@163.com
