橡胶草可作为产胶研究的模式植物
2015-10-14庄玉粉郭秀丽刘进平
庄玉粉++郭秀丽++刘进平
摘 要 对天然橡胶生物合成途径进行综述,并根据橡胶草和巴西橡胶树的生物学特点,提出将橡胶草作为研究橡胶生物合成的模式植物。橡胶草作为产胶模式植物,将会极大地推动天然橡胶生物合成与调控机理的研究,提高目前的科研产出。
关键词 天然橡胶 ;生物合成 ;橡胶草 ;巴西橡胶树 ;模式植物
分类号 S7 ;Q945.11
Russian Dandelion: An Excellent Model Plant
for the Study of Rubber Production
ZHUANG Yufen GUO Xiuli LIU Jinpin
(Hainan Key Laboratory for Sustainable Utilization of Tropical Bioresources /
College of Agronomy, Hainan University, Haikou,Hainan 570228)
Abstract The biosynthesis pathway of natural rubber was reviewed, and Russian dandelion (Taraxacum koksaghyz) was proposed as a model plant for studies of rubber biosynthesis based on the comparison of the biological characteristics of Russian dandelion and rubber tree (Hevea brasiliensis). We believe that it will greatly promote the research in the natural rubber biosynthesis and its regulation mechanism, and increase the research output for this area regarding Russian dandelion as a mode plant for the study of rubber production.
Keywords natural rubber ; rubber biosynthesis ; Russian dandelion ; rubber tree ; model plant
天然橡胶是世界上的四大工业原料之一,被广泛运用在国防、交通运输、工业、农业、机械制造、医药卫生和日常生活等多个方面,是国防和工业制造中不可或缺的原料。天然橡胶目前主要是从橡胶树上收获而得的,它的主要成分为聚异戊二烯(cis-1,4,-polyisoprene),是利用光合作用制造的同化物,经次生代谢途径而生物合成。虽然天然橡胶的合成研究已取得了不少进展,但由于巴西橡胶树自身的生物学特点,存在对天然橡胶生物合成研究的瓶颈,为了推动天然橡胶生物合成与调控机理的研究,提高目前的科研产出,探讨以巴西橡胶树替代作物橡胶草作为模式生物研究橡胶生物合成的可能性。
1 天然橡胶的生物合成
天然橡胶生物合成大体可分为两步:第一步是前体物合成;第二步是橡胶链的合成。
1.1 前体物合成
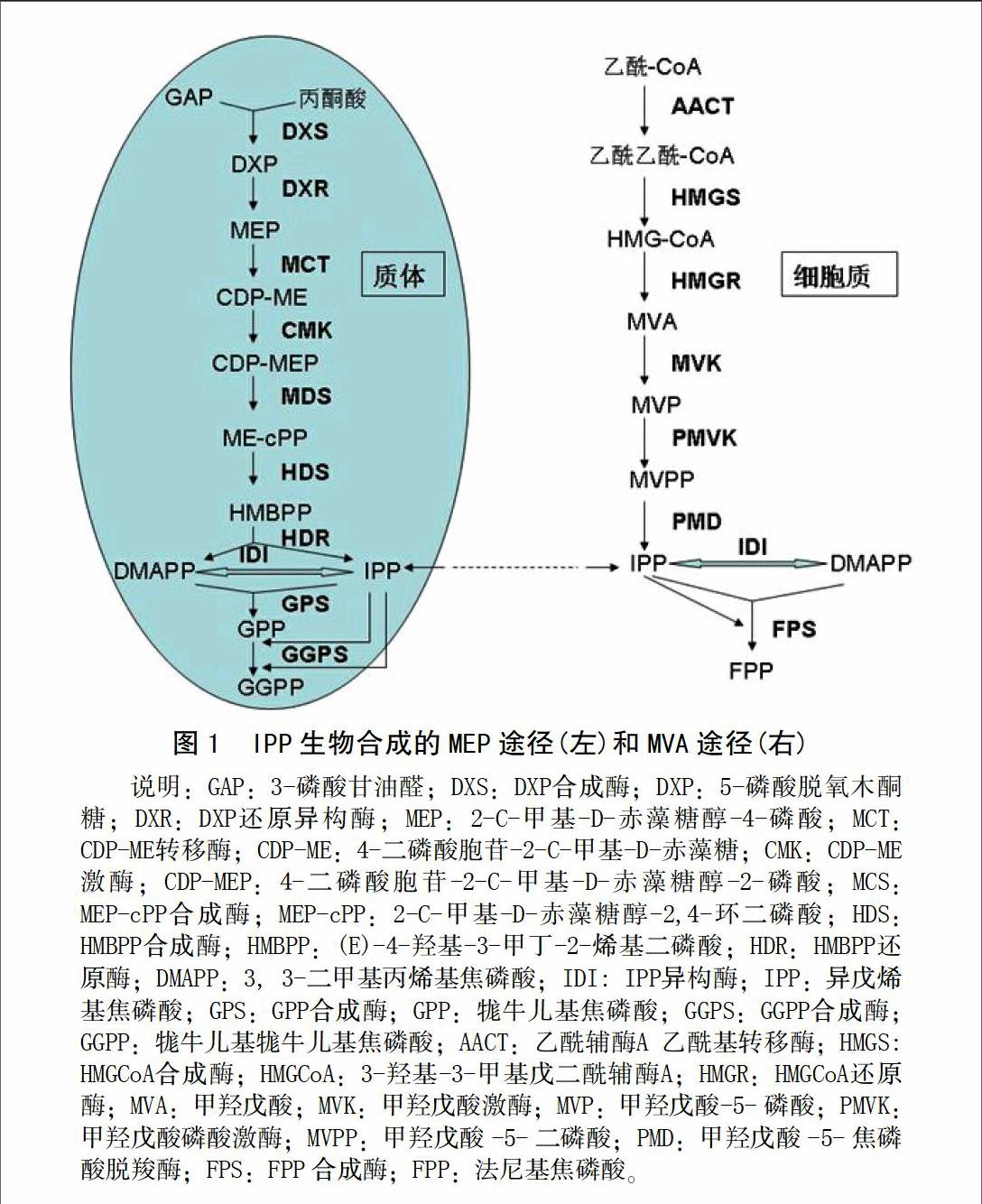
橡胶生物合成所需要的单体模块为异戊二烯焦磷酸(isopentenyl pyrophosphate, IPP或IDP),起始分子包括3,3-二甲基丙烯基焦磷酸(dimethylallyl pyrophosphate, DMAPP或DMADP)、牻牛儿基焦磷酸(geranyl pyrophosphate, GPP或GDP)、法尼基焦磷酸(farnesyl pyrophosphate, FPP或FDP)和牻牛儿基牻牛儿基焦磷酸(geranylgeranyl pyrophosphate, GGPP或GGDP)[1]。
橡胶前体物被认为是通过细胞质中的甲羟戊酸(mevalonic acid, MVA)途径合成的[1]。Sando等[2]曾利用位素示踪表明橡胶生物合成是通过MVA途径,而不是通过质体中的2-甲基-D-赤藓糖醇-4-磷酸(2-C-methyl-D-erythritol 4-phosphate, MEP)途径。但他们是利用橡胶35 d的实生苗进行示踪实验的,成熟的橡胶树胶乳合成是否也如此不得而知。鉴于越来越多的证据表明,这两条途径存在串扰(crosstalk)和共同的类异戊二烯前体物相互交换[3-4],因此也不能排除在橡胶合成过程中这两条途径存在互相作用和产物互换(图1)。在橡胶胶乳和叶片中发现5-磷酸脱氧木酮糖合成酶(1-deoxy-D-xylulose 5-phosphate synthase, DXS)表达说明,可能在乳管中存在MEP途径[5]。最近对橡胶树死皮树录组分析表明,合成IPP的这两条途径的关键酶基因都明显下调[6],显示天然橡胶合成中MVA和MEP途径可能存在某种方式的互作。
MEP途径主MVA途径和前体物都是来源于光合作用的产物——蔗糖。MEP途径的前体物3-磷酸甘油醛(glyceraldehyde phosphate,GAP)和丙酮酸为糖酵解产物,而MVA途径的前体物乙酰-CoA是经糖酵解和三羧酸循环途径生成的。
MEP途径中,DXS催化GAP和丙酮酸的酮醇偶联缩合生成5-磷酸脱氧木酮糖(1-deoxy-D-xylulose 5-phosphate,DXP)。DXP由5-磷酸脱氧木酮糖还原异构酶(DXP reducto-isomerase,DXR或IspC)催化形成MEP。MEP向二磷酸胞苷-2-C-甲基-D-赤藻糖[4-(cytidine-5′-diphospho)-2-C-methyl-D-erythritol或4-diphosphocytidyl-2-C-methyl-D-erythritol,CDP-ME]转变则由4-磷酸胞苷-2-甲基赤藓糖合成酶(CDP-ME synthase,MCT、CMS或IspD)催化。之后CDP-ME激酶(CDP-ME kinase,CMK或IspE)使CDP-ME磷酸化形成4-二磷酸胞苷-2-C-甲基-D-赤藻糖醇-2-磷酸(4-diphosphocytidyl-2-Cmethyl-D-erythritol-2-phosphate,CDP-MEP)。CDP-MEP环化形成2-C-甲基-D-赤藻糖醇-2,4-环二磷酸(2-C-methyl-D-erythritol-2,4-cyclodiphosphate,MEP-cPP)则由MEP-cPP合成酶(MEP-cPP synthase,MDS或IspF)催化。MEP-cPP形成(E)-4-羟基-3-甲丁-2-烯基二磷酸(4-hydroxy-3-methylbut-2-enyl diphosphate,HMBPP)由HMBPP合成酶或1-羟基-2-甲基-2-(E)-丁烯-4-二磷酸合成酶(HMBPP synthase,HDS或IspG)催化。然后HMBPP还原酶(HMBPP reductase,HDR或IspH)催化HMBPP形成IPP和DMAPP。IPP异构酶(IPP isomerase, IDI或IPI)可催化IPP和DMAPP相互转化。
MVA途径的第一步反应是乙酰-CoA在乙酰基转移酶(acetyl-CoA acetyltransferase, AACT)催化下,2分子乙酰-CoA缩合成乙酰乙酰-CoA(acetoacetyl coenzyme A)。乙酰乙酰-CoA经3-羟基-3-甲基戊二酸单酰辅酶A合成酶(3-hydroxy-3-methylglutaryl-coenzyme A synthase, HMGS)催化合成3-羟基-3-甲基戊二酸单酰辅酶A(3-hydroxy-3-methylglutaryl-coenzyme A, HMG-CoA)。之后经HMG-CoA 还原酶(HMG-CoA reductase, HMGR)催化还原成MVA。MVA经两步磷酸化过程,经甲羟戊酸激酶(mevalonate kinase, MVK)催化形成甲羟戊酸-5-磷酸(5-phosphomevalonate, MVP或MVAP),经甲羟戊酸-5-磷酸激酶(phosphomevalonate kinase, PMK或 PMVK)形成甲羟戊酸-5-焦磷酸(5-diphosphomevalonate, MVPP或MVAPP)。MVPP经甲羟戊酸-5-焦磷酸脱羧酶(mevalonate-5-pyrophosphate decarboxylase, PMD或MVD)脱羧形成IPP。IPP和DMAPP可经IDI相互转化。IPP在法尼基焦磷酸合成酶(farnesyl pyrophosphate synthase, FPS)的作用下与DMAPP合成FPP。
需要说明的是,除形成橡胶外,IPP、DMAPP、GPP、FPP和GGPP等可进一步形成异戊二烯单元组成的其他化合物及其衍生物,包括单萜类(如薄荷醇)、双萜类(如赤霉素、脱落酸、蓖麻烯)、倍半萜类(如衣壳醇)、异戊二烯基类(如细胞分裂素)、多聚异戊二烯醇(如多萜醇、酶Q、质体醌、类胡萝卜素、叶绿素)、固醇类(如菜油甾醇)等次生代谢产物[7]。
1.2 橡胶链的合成
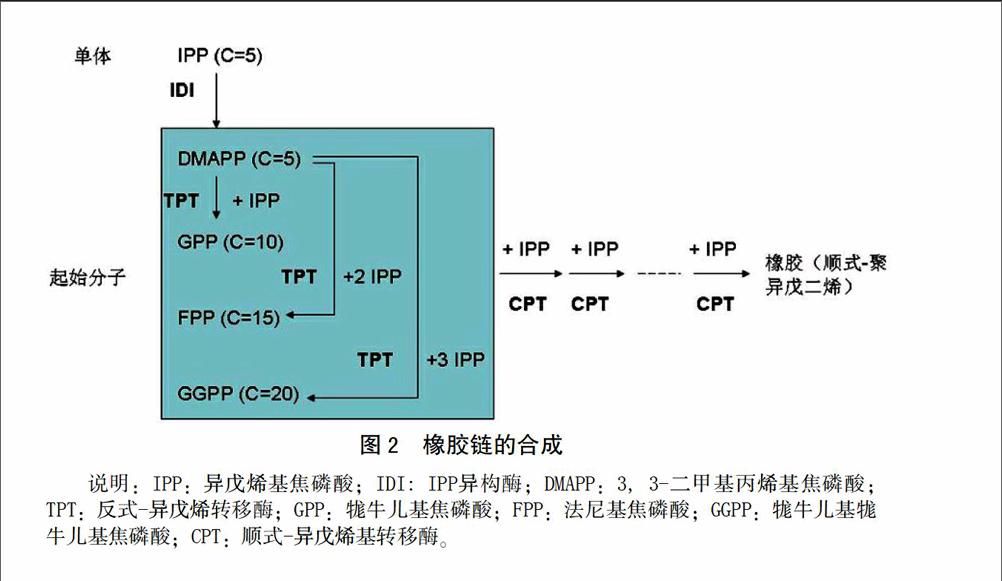
天然橡胶主要成份为聚异戊二烯(cis-1,4-polyisoprene),聚异戊二烯链在橡胶粒子表面合成。橡胶粒子存在于巴西橡胶树[Hevea brasiliensis (Willd. ex. A.Juss.) Muell. Arg]的胶乳细胞中。
橡胶链的合成中,起始过程需要1 分子反式构型的烯丙基焦磷酸(如DMAPP、GPP、FPP 或GGPP)作为起始分子或引物。反式-异戊烯转移酶(transprenyltransferase, TPT)催化IPP单体以反式构型头尾相接地掺入DMAPP,产生GPP、FPP和GGPP(图2)。DMAPP、GPP、FPP和GGPP等虽然都可以作起始分子,但以橡胶树等物种的体外橡胶生物合成中,FPP起始的橡胶合成最有效率。加上FPP是在细胞质中合成,因此在橡胶树中FPP是最有可能的起始分子[1,8-9]。
顺式-异戊烯基转移酶(cis-prenyltransferase, CPT)催化IPP单体以顺式构型头尾相接地添加到结合于起始分子上,形成长短不等的橡胶分子。橡胶链延伸过程需要二价金属阳离子Mg2+或Mn2+参与[8-9]。
由于橡胶粒子与天然橡胶合成紧密联系,分离到的橡胶粒子可以产生橡胶,同时破坏橡胶粒子则会破坏橡胶生物合成,因此表明橡胶粒子是橡胶转移酶活性必不可少的组分。橡胶粒子上的蛋白质数量和质量随不同物种而异。目前较为清楚橡胶树的橡胶粒子具有数量较多的蛋白。其中比重最大的是14-ku的橡胶延长因子(rubber elongation factor, REF)和22-ku的橡胶小粒子蛋白(small rubber particle protein, SRPP)[1]。
2 天然橡胶生物合成研究瓶颈
目前巴西橡胶树研究已取得了长足的进步,但存在较大的瓶颈,限制了对巴西橡胶树的深入研究。以橡胶树的橡胶生物合成为例,虽然大体的合成途径已经清楚,前述IPP生物合成的MEP途径、MVA途径及后继的橡胶链合成途径中的重要基因已经克隆[10],但MEP途径是否参与橡胶合成,IPP合成途径中的关键酶基因对橡胶合成的数量(橡胶产量)和质量(如橡胶链的长短)的影响和贡献并不清楚,对其分子调控机理也了解不多。对这些方面的任何进展都需要对相关基因进行克隆和遗传转化,通过对这些基因的(利用强启动子)过量表达和(利用反义基因)沉默表达来观察其对橡胶合成的影响。然而,橡胶树本身组织培养比较困难,遗传转化成功率极为有限,此外,即便遗传转化获得转基因植株,因为橡胶树为多年生木本植物,生长周期长,存在幼态和成熟两种状态,橡胶树需要在田间生长5~7 a才能割胶,等到能收获胶乳以确定对性状的影响也需要在5~7 a之后[11-12]。这些都是目前以巴西橡胶树为对象的天然橡胶生物合成的研究瓶颈。
3 巴西橡胶树替代植物研究现状
已报道有2 500种以上的植物可以产生橡胶,但绝大多数为热带植物。巴西橡胶树(Hevea brasiliensis Muell. Arg.)是目前商业化种植面积最大、产量最多的产胶植物,植胶区主要分布于亚洲、非洲和南美等热带及亚热带地区,其中亚洲橡胶产量占全球93%。由于巴西橡胶树主要产于热带地区,且易受南美叶疫病感染,而绝大多数发达国家依赖进口,所以欧盟、美国、澳大利亚等国相继开展了巴西橡胶树替代产胶植物研究。目前研究最多的2种替代产胶植物为银胶菊(英文名:Guayule,拉丁学名:Parthenium argentatum Gray)和橡胶草(又名俄罗斯蒲公英,英文名:Russian dandelion,拉丁学名:Taraxacum koksaghyz Rodin)。
国内已有作者对橡胶草的研究进展及作为巴西橡胶树替代作物研发进行了详尽的综述[13-16],这里主要就橡胶草作为模式生物研究橡胶生物合成意义和可能性进行论述,希望引起国内研究者的注意。
4 橡胶草作为模式生物研究橡胶生物合成的可能性
与多年生的木本植物巴西橡胶树相比,橡胶草有很多优势。首先,它为菊科菊苣族蒲公英属的多年生草本植物,在温带可以如菊苣一样种植,作为一种一年生植物(从播种到收获需1年)开发[11-12]。这样就可避免经历橡胶树慢长的幼态期之后才能对其胶乳产生进行评估。
其次,橡胶草组织培养和遗传转化都比较容易,获得的转化苗达到产胶状态所需时间短。转基因的橡胶草植株4个月就可以成熟,在移栽到田间的头两个月就能产生可检测数量的橡胶。而另外一种巴西橡胶树替代作物银胶菊虽然也是草本植物,但它需要在田间生长2年才能进行橡胶分析[11-12]。橡胶草遗传转化的这种便利性非常适合利用过量表达和沉默表达来研究天然橡胶生物合成途径中关键基因对橡胶合成的影响及其调控机制。
第三,橡胶草核型为2n=2x=16,基因组为1 450 Mb[10];巴西橡胶树2n=4x=36,具有双二倍体特征,其单倍体基因组大小约4×109 bp[17]。橡胶树产胶部位为树皮,橡胶草产胶部位为根,两者都有类似的乳管系统,但橡胶草不能如橡胶一样切割收获,必须将根搅碎后进行压榨或提取橡胶。而银胶菊则无乳管或相连的乳管网络,收获胶乳需要研磨植物组织以释放橡胶粒子[11-12]。对橡胶草和巴西橡胶树进行比较基因组学、转录组学和蛋白组学将获得更多产胶相关的分子信息。
总之,对橡胶草的研究,不仅是作为替代产胶植物本身,也将为巴西橡胶树乳管发育、橡胶粒子形成和橡胶合成研究提供重要的参考信息,因此,就象水稻作为单子叶禾本科类粮食作物的优良模式植物一样,橡胶草也是一种优良的产胶模式植物,预计未来对其研究产出将会有较大增长。
参考文献
[1] Puskasa J E, Gautriauda E,Deffieuxb A, et al. Natural rubber biosynthesis-A living carbocationic polymerization[J]. Progress in Polymer Science, 2006, 31 (6): 533-548.
[2] Sando T, Takeno S, Watanabe N, et al. Cloning and characterization of the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway genes of a natural rubber producing plant, Hevea brasiliensis[J].Biosci Biotechnol Biochem, 2008, 72 (11): 2 903-2 917.
[3] Rodríguez-Concepción M. Early Steps in Isoprenoid Biosynthesis: Multilevel Regulation of the Supply of Common Precursors in Plant Cells[J]. Phytochemistry Reviews, 2006, 5(1): 1-15.
[4] Rodríguez-Concepción M, Campos N, Ferrer A,et al. Biosynthesis of isoprenoid precursors in Arabidopsis[M]. In: Bach TJ, Rohmer M (eds.), Isoprenoid Synthesis in Plants and Microorganisms: New Concepts and Experimental Approaches, Springer Science+Business Media, New York, 2013: 439-456
[5] Ko J H, Chow K S, Han K H. Transcriptome analysis reveals novel features of the molecular events occurring in the laticifers of Hevea brasiliensis (para rubber tree)[J]. Plant Mol Biol, 2003, 53 (4): 479-492
[6] Liu J P, Xia Z Q, Tian X Y, et al. Transcriptome sequencing and analysis of rubber tree (Hevea brasiliensis Muell.) to discover putative genes associated with tapping panel dryness (TPD) [J]. BMC Genomics, 2015, 16: 398.
[7]李 莉,高凌云,董 越,等. 植物类异戊二烯生物合成相关酶基因研究进展[J]. 浙江师范大学学报(自然科学版),2008,31(4):461-466.
[8] 代龙军,曾日中. 产胶植物橡胶转移酶的研究进展[J]. 植物生理学报,2013,49(10):1 000-1 008.
[9] 邹 智,杨礼富,王真辉,等. 橡胶树中橡胶的生物合成与调控[J]. 植物生理学通讯,2009,45(12):1 231-1 238.
[10] Sando T, Takaoka C, Mukai Y, et al. Cloning and characterization of mevalonate pathway genes in natural rubber producing plant, Hevea brasiliensis[J]. Biosci Biotechnol Biochem, 2008, 72 (8): 2 049-2 060.
[11] Whalen M, McMahan C, Shintani D. Development of crops to produce industrially useful natural rubber[M]. In: Bach TJ, Rohmer M (eds.), Isoprenoid Synthesis in Plants and Microorganisms: New Concepts and Experimental Approaches, Springer Science+Business Media, New York, 2013: 329-345.
[12] Van Beilen JB, Poirier Y. Establishment of new crops for the production of natural rubber. Trends in Biotechnology[J]. 2007, 25(11): 522-529.
[13] 仇 键,张继川,罗世巧,等. 橡胶草的研究进展[J]. 植物学报,2015,50(1):133-141.
[14] 林伯煌,魏小弟. 橡胶草的研究进展[J]. 安徽农业科学,2009(37):5 950–5 951.
[15] 安 锋,林位夫,谢贵水,等. 国内外巴西橡胶树替代作物及技术研发现状[J]. 热带作物学报,2012,33(6):1 134-1 141.
[16] 赵平娟,安 锋,林位夫,等. 大力开展巴西橡胶树替代产胶植物及技术研发的建议[J]. 中国农学通报,2012,28(34):124-130.
[17] Roy C B, Nazeer M A, Saha T. Identification of simple sequence repeats in rubber (Hevea brasiliensis) [J]. Curr Sci, 2004, 87: 807-811.
