东北红豆杉光合特性研究
2015-09-17张永君
余 刚,邢 昊,张永君
(吉林省林业调查规划院,吉林 长春 130022)
光合作用是植物的基本生命活动之一,是植物吸收太阳能量、同化二氧化碳和水,制造有机物质、释放氧气的过程;是太阳辐射能进入生态系统并转化为化学能的主要形式,也是制约生态系统生物生产力的最重要的生理过程。植物光合作用是有机物质积累的基础,直接影响着生态系统水热平衡和物质循环过程,也是全球碳循环及其他物质循环的最重要环节。
光合作用是一个复杂的生物物理化学过程,受植物本身内部因素和外界环境因子影响。研究植物光合特性也主要从植物的生理因子如气孔导度、胞间CO2浓度等和生态因子如光照强度、二氧化碳浓度、温度、湿度等方面展开。目前对植物光合特性的研究主要有赵敏等在雅安市荥经县川黄柏规范化种植(GAP)基地,采用Li-6400 光合测定系统,以2 a 生黄蘖为研究对象,探讨了黄蘖幼树叶片在生长季后期净光合速率日变化[1]。陈晓强等选择4 个品种的桃树,对不同叶位的叶片光合作用进行研究[2]。罗树伟等利用Li-6400 光合作用测定仪,在长柄扁桃野生种质资源(种源)进行光合生理生态特性及影响因子研究[3]。江锡兵等采用Li-6400 光合测定系统对美洲黑杨与大青杨杂种3 个无性系的光合作用指标进行测定[4]。王旭军等用Li-6400 便携式光合作用测定系统对自然条件下城市园林树种乐昌含笑的光响应特性、CO2响应特性及光合作用日变化规律进行测定[5]。张朋伟等以浙江省常见的16 种园林植物为研究对象,利用Li-6400便携式光合作用测定系统测定其光合响应曲线,分析其在光能利用率(LUE)、CO2固定效率(CE)、水分利用率(WUE)方面的能力,评价其改善环境的效益[6]。李文华等对7 个主要造林树种,在2005 和2006 年的两个生长季,利用树干茎流仪、WP4 土壤-植物水势仪和Li-6400光合测定系统仪,研究了不同树木根层土壤水分、水势的动态变化,及植物叶片水势、蒸腾速率、水分利用率、光合特性和荧光特性[7]。季慧颖等应用Li-6400 对北京市引进的4 种乔木北美鹅掌楸、银槭、七叶树和水曲柳的光合特性及生态效益进行了研究[8]。
东北红豆杉(Taxus cuspidata S. et Z.),又名紫杉、赤柏松,是红豆杉科(Taxaceae)红豆杉属(Taxus)乔木植物,为第三纪孑遗的珍贵树种。红豆杉有“植物活化石”和“国宝”之称。是世界上濒临灭绝的天然珍稀抗癌植物,属国家Ⅰ级保护植物,至今已有250 万年的历史[9]。目前国内外学者对东北红豆杉做了大量研究,主要集中在生态群落、繁殖技术、化学医药、组织细胞培养等方面,而对东北红豆杉光合特性的研究却较少,张启昌等利用Li-6400 测定了不同CO2浓度条件下的东北红豆杉净光合速率、蒸腾速率、气孔导度、胞间CO2浓度及叶面饱和蒸气压亏缺,结果表明光合速率的显著增加是导致东北红豆杉水分利用效率随CO2浓度升高而增加的主要影响因素[10]。丁磊等在100%、70%、40%和10%四种光照条件下对东北红豆杉苗期生长及光合特性进行了研究,得出在100%、70%光照下,净光合速率日变化呈现“双峰型”曲线,有明显的“光午休”现象及引起“光午休”的主要环境因子和70%光照是东北红豆杉苗期生长的适宜光环境的结论[11,12]。
作者利用Li-6400 便携式光合仪对东北红豆杉光合特性进行测定,总结分析比较,掌握东北红豆杉的光合规律;研究内容分别为光合速率的日变化、不同光照强度的光合生理响应和叶绿素荧光特征。旨在为探讨其生长发育的一般规律、科学利用与有效保护这一种质资源提供基础数据。
1 研究地点、材料与方法
1.1 研究地点
研究地位于吉林市北华大学校园内。气候为北温带大陆气候,夏季温热多雨,冬季寒冷漫长,年平均气温4.5℃,1 月平均气温-17.9℃,最低温度-40.2℃;7 月平均气温22.9℃,最高温度33.6℃。平均降水量为668 mm,相对湿度为70%。无霜期130 d 左右。
1.2 研究材料和方法
采用美国Li-COR 公司生产的Li-6400便携式光合仪应用红外气体分析法对东北红豆杉进行测定,本试验选择3 雌株3 雄株为待测树,每次测定时选择同一树木正常生长枝条上的健康叶片作为待测叶。测定东北红豆杉的不同枝条(阳面、阴面)的净光合速率、气孔导度、气温、胞间CO2浓度、大气CO2浓度和光照强度等因子,当测量结果变异率小于0.05 时由红外气体分析仪记录,重复3 次,取平均值。
光合作用的日变化测定:选择在5 月中旬到6 月中旬晴天进行,从6:00 ~18:00 时,对阴阳两面的待测叶每隔2 h 测定1 次,由仪器记录东北红豆杉随时间变化的净光合速率。
对东北红豆杉光合速率影响因子进行相关分析。所选因子为气孔导度、胞间CO2浓度、蒸腾速率、叶面温度,样本室CO2浓度、样本室内气体相对湿度、样本室H2O 浓度、样本室内有效光合辐射。用SPSS 软件对因子变量的相关性进行分析。

光合作用的光响应测定:利用Li-6400 自动曲线测定功能,选择红蓝光源,设定一系列光合有效辐射(PAR)梯度,分别为:0 μmol·m-2·s-1、50 μmol·m-2·s-1、100对湿度85%5%。改变光照强度以后,最少稳定时间设置为120 s,由红外气体分析仪自动记录。采用经典的Farquhar 模型来进行拟合,统计方法采用SPSS 统计软件,利用其中的非线性统计分析模块。

式中:A—净光合速率;
Amax—最大净光合速率;
Q—表观量子效率;
K——曲角;
RD—光下呼吸速率;
light—有效光合辐射。
叶绿素荧光参数测定:使用Li-6400 光合作用测定系统的荧光叶室。测定前,叶片先于暗中适应20 min,测定初始荧光(F0),再照射饱和脉冲(1 000 μmol·m-2·s-1),测定暗适应下最大荧光(Fm)、光化学量子效率(Fv/Fm)等;然后将光脉冲调至1 000 μmol·m-2·s-1,叶片充分光适应30 min,测定光下最小荧光(F0')、光下最大荧光(Fm')、稳定荧光(Fs)、激发能捕获效率(Fv'/Fm')、φPsⅡ实际光量子效率(PhiPS2)、光化学猝灭系数(qP)、非光化学猝灭系数(qN)等。所有测定指标由仪器自动记录。测定6 个植株,每叶片记录3 次,结果取平均值。
以上各测定结果使用Excel 对数据进行整理,使用SPSS 统计软件对数据进行相关分析、方差分析及多元回归分析。
2 结果与分析
2.1 东北红豆杉光合速率日变化
从图1 中可以看出(由于操作原因,每天测量时雄株记录较雌株晚半小时),东北红豆杉雌雄株之间的光合速率日变化有着明显不同,雌株呈单峰形状,雄株呈双峰形状。雌株在8:00 ~10:00 之间光合速率随时间不断增加,到11:00 左右达最大0.99 μmol·m-2·s-1,且光合速率高峰在11:00 ~15:00 之间持续,15:00以后则逐渐降低直至光合作用停止。雄株在8:00 ~10:00 之间光合速率随时间也不断上升,在 10:30 左右达到第一个峰值1.23 μmol·m-2·s-1,峰 值 比 雌 株 稍 早 达到,且峰值比雌株大。在10:30 ~13:50 间,雄株呈凹形,在14:00 左右达第二个峰值1.13 μmol·m-2·s-1,由此可看出雄株有明显的午休现象。随后逐渐降低。另外,雄株光合速率在14:30 以前比雌株略高,表明雄株在同样光合条件下,对光的吸收和利用较雌株高。而在14:30 以后,雄株的光合速率下降较雌株快,表明雄株对光照强度的要求高于雌株。
2.2 东北红豆杉光合速率影响因子分析

图1 雌雄株光合速率日变化Fig.1 Photosynthetic rate diurnal variation of female and male plants
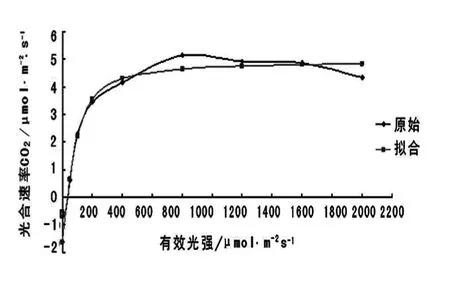
图2 东北红豆杉光响应曲线Fig.2 Photoresponse curve of Taxus cuspidata

表1 Bivariate(双变量)相关分析Tab.1 Bivariate correlation analysis
从表1 中可以看出,光合速率与样本室内气体相对湿度无显著相关性,与其他几个因子相关系数都有显著性,与蒸腾速率、样本室内有效光合辐射相关系数较大,与样本室CO2浓度和胞间CO2浓度呈负向相关。气孔导度与蒸腾速率相关性较大,相关系数高达0.98,与样本室内有效光合辐射也显著相关。叶面温度与样本室内有效光合辐射也显著相关,相关系数为0.709。综上可知,东北红豆杉的光合速率与蒸腾速率、有效光合辐射、气孔导度、叶面温度呈显著正相关,与样本室CO2浓度、胞间CO2浓度呈显著负相关。
2.3 东北红豆杉对不同光照强度的光合生理响应

表2 非线性回归统计参数估计Tab.2 Parameter estimation of nonlinear regression statistics
根据表2 各参数的估计值,K 为0.729,Amax为6.593,Q 为0.053,RD为1.642。
代入Farquhar 模型的理论公式求出拟合值,作出拟合曲线如图2 所示。
从图2 中可以看出,拟合后的曲线可以看出当光合有效辐射为0 μmol·m-2·s-1时,光合速率为负值;在0 ~400 μmol·m-2·s-1时,光合速率随光合有效辐射的增加而显著增加,在400 μmol·m-2·s-1以后,光合速率不再随光合有效辐射而增加,曲线几近与横坐标轴平行,表明在400 μmol·m-2·s-1左右为光饱和点。
将光合速率与光合有效辐射强度在0 ~200 μmol·m-2·s-1之间进行线性回归,求出线性方程,代入最大净光合速率(Amax),即可求出东北红豆杉的光饱和点(LSP),令光合速率为0,即可求出光补偿点(LCP)。

表3 简单线性回归常数项和系数的估计及检验Tab.3 Estimation and test of simple linear regression constant term and coefficient

图3 雌雄株光化学量子效率Fv/Fm和激发能捕获效率Fv'/Fm'Fig.3 The photochemistry quantum efficiency Fv/Fm and the excitation energy capture efficiency Fv'/Fm'of female and male plants
从表3 中可以看出,参数估计常数项为-0.179,P 值为0.847 大于0.05,则常数项无统计学意义。系数估计值为0.013,P 值为0.048,有统计学意义。
建立的线性回归方程为:
A= -0.179 +0.013·light
其中:A 为净光合速率,light 为有效光合辐射。
将光合速率最大值Amax和0 值代入上述方程 有:光 饱 和 点 (LSP)= 520. 923 μmol·m-2·s-1,光补偿点(LCP)=13.769 μmol·m-2·s-1。
LSP 和LCP 是植物利用光强能力的重要指标[13],LSP 高的植物被认为能更有效地利用强光,LCP 低的植物被认为能更有效地利用弱光[14],由所求出的光补偿点可以看出东北红豆杉对弱光能有效利用、光饱和点可以看出其对有效光合辐射要求不高,520 μmol·m-2·s-1左右即能饱和。另外,光补偿点低且光饱和点相应也低的植物具有很强的耐阴性[8],由所求的LSP 和LCP 可知,从理论数据上得出东北红豆杉具有很强的耐阴性。
2.4 东北红豆杉叶绿素荧光特征
叶绿素荧光主要反映了光合机构光系统II(PSII)的生理生化变化[15],从图3 中可以看出,雌雄株的最大(潜在)光化学量子效率Fv/Fm(也被称为开放的PSII 反应中心的能量捕获效率)没有明显的差异,表明雌雄株暗适应后的能量捕获效率几乎是相同的。而雌雄株之间的激发能捕获效率Fv'/Fm'差异相对明显,表明在光下开放的PSII 反应中心雌株的激发能捕获效率较雄株的高,能捕获较多的激发能。
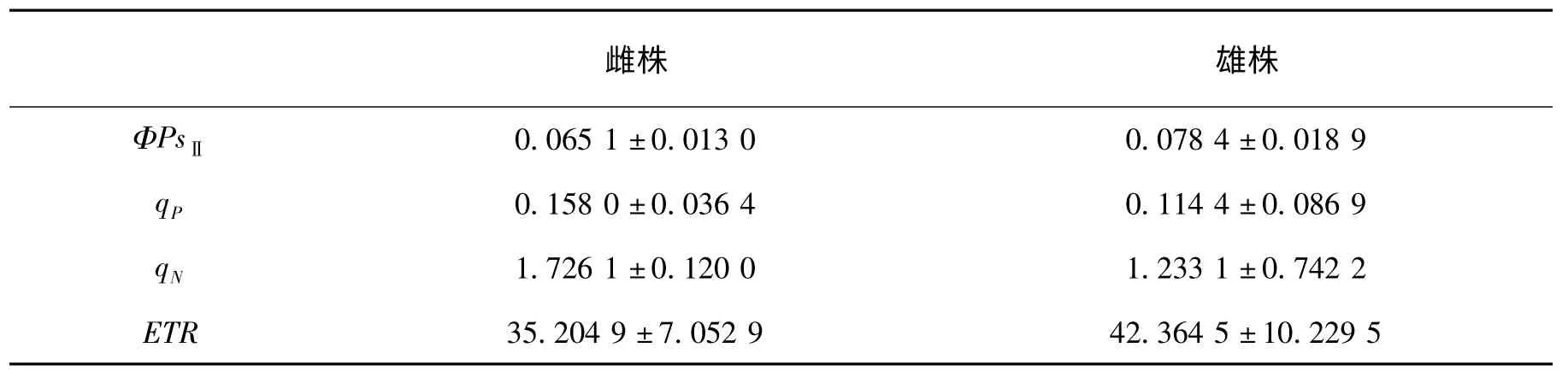
表4 东北红豆杉雌雄株叶绿素荧光参数比较Tab.4 The chlorophyll fluorescence parameters comparison of female and male plants of Taxus Cuspidata
从表4 中可以看出,雄株的光化学量子效率ΦPsⅡ是0.078 4±0.018 9,比雌株的0.065 1±0.013 0 略大,表明雄株被用于光化学途径激发能占进入PSⅡ总激发能的比例较多,反映了雄株比雌株光化学反应效率略高。雌株的光化学淬灭系数qP为0.158 0±0.036 4,较雄株的0.114 4±0.086 9 大,反映了雄株PSⅡ反应中心中开放程度没有雌株的大。非光化学淬灭系数qN雌株是1.726 1±0.120 0,比雄株的1.233 1±0.742 2 稍大,表明雌株热耗散比雄株大,反映了雄株用于热耗散的光能较小,其光能利用率较雌株的略高。还可以看出电子传递速率ETR 雄株的为42.364 5±10.229 5,比雌株的35.204 9±7.052 9 大。

表5 东北红豆杉雌雄株荧光参数的方差分析Tab.5 Fluorescence parameter variance analysis of female and male plants of Taxus cuspidata
从表5 中可以看出,P 值均大于0.05,表明雌雄株之间的荧光参数差异不显著。从图3、表4、表5 的分析可以得出东北红豆杉雌雄株之间的荧光参数在数值上雄株较雌株略大,但数值之间不具有显著的差异性,表明东北红豆杉雌雄株之间叶绿素荧光特征没有明显差异,光合机构光系统Ⅱ(PsⅡ)的变化基本是相同的。
3 结论
东北红豆杉雌株与雄株之间的光合速率日变化有着明显的不同,雌株日变化曲线呈单峰形,雄株呈双峰形,有明显的午休现象。
拟合出东北红豆杉对不同有效光合辐射的光响应曲线,并由此曲线求出其光饱和点为520 μmol · m-2· s-1,光 补 偿 点 为13.769 μmol·m-2·s-1,从数据理论上证明东北红豆杉具有很强的耐阴性。
雌雄株之间的叶绿素荧光参数最大的(潜在)光化学量子效率Fv/Fm基本相同,而激发能捕获效率Fv'/Fm'雌株较雄株略高。但对所测叶绿素荧光参数所做的方差分析来看,雌雄株之间的各参数Fv/Fm、Fv'/Fm'、qN、qP、φPsⅡ、ETR 并无显著性差异。因此,雌雄株之间叶绿素荧光特征无差异,光系统Ⅱ(PsⅡ)的生理生化变化基本是相同的。
[1]赵敏,叶萌,严晓宇,等.黄柏幼树光合特性研究[J].林业科技开发,2007,21(5):43-46.
[2]陈晓强,姜卫兵,俞明亮,等.桃不同叶位光合特性研究[J].江苏农业科学,2007,(5):104-105.
[3]罗树伟,郭春会,张国庆,等.沙地植物长柄扁桃光合特性研究[J].西北农林科技大学学报(自然科学版),2010,38(1):125-132.
[4]江锡兵,李博,张志毅,等.美洲黑杨与大青杨杂种无性系苗期光合特性研究[J].北京林业大学学报,2009,31(5):151-154.
[5]王旭军,吴际友,廖德志,等.主要城市园林树种乐昌含笑光合特性研究[J].西南林学院学报,2008,28(5):26-30.
[6]张朋伟,余树全,伊力塔,等.不同类型园林树木光合特性研究[J].农业科技与信息(现代园林),2010,(12):55-59.
[7]李文华.陕北黄土区主要造林树种蒸腾耗水及光合特性研究[D].北京:北京林业大学,2007.
[8]季慧颖.北京四种引进植物光合特性及生态效益研究[D].北京:北京林业大学,2010.
[9]刘彤.天然东北红豆杉种群生态学研究[D].哈尔滨:东北林业大学,2007
[10]张启昌,赵影,其其格,等.东北红豆杉枝叶对不同浓度CO2的光合生理响应[J]. 北华大学学报,2006,7(1):66-70.
[11]丁磊,胡万良,孔祥文,等.遮荫对东北红豆杉苗期生长及光合特性的影响[J].吉林林业科技,2008,37(1):1-4.
[12]孙义男,郑昕. 穆棱东北红豆杉国家级自然保护区景观斑块特征分析[J]. 森林工程,2014,30(4):46-49.
[13]曹军胜,刘广全.刺槐光合特性的研究[J].西北农业学报,2005,14(3):118-122,136.
[14]朱万泽,王金锡,薛建辉.台湾桤木引种的光合生理特性研究[J].西北植物学报,2004,24(11):2012-2019.
[15]孙红英,曹光球,辛全伟,等. 香樟8 个无性系叶绿素荧光特征比较[J].福建林学院学报,2010,30(4):309-313.
