米槁净光合速率与生理生态因子的关系
2015-09-10王军才廖小锋刘济明等
王军才 廖小锋 刘济明等


摘要: 为了探明米槁(Cinnamomum migao H W Li )光合作用日变化特征及其与生理生态因子间的关系,采用 Li-6400 光合测定仪对米槁叶片进行生理生态指标测定,并运用多元逐步回归和通径分析相结合的方法分析了净光合速率与生理生态因子间的关系。结果表明:米槁净光合速率(Pn)日变化为典型的双峰曲线,第1峰[HJ1mm][峰值 10 18 μmol/(m2·s)] 出现在10:00左右,第2峰[峰值7 93 μmol/(m2·s)]出现在15:00左右,有明显的光合“午休”现象,气孔因素是导致“午休”的主要原因。通过多元逐步回归分析,得出米槁净光合速率最优回归方程为:Pn=21 257+100 982Gs+0168Gi+0 067Tr-0 056Ca(R2=0 921)。综合回归分析与通径分析的结果,得出影响米槁叶片净光合速率的主要生态因子是大气CO2浓度(Ca)和空气相对湿度(RH),主要生理因子是气孔导度(Gs)和胞间CO2浓度(Ci)。
关键词: 米槁;光合作用日变化;净光合速率;生理生态因子;回归分析;通径分析
中图分类号: S718 45 文献标志码:A
文章编号:1002-1302(2015)08-0256-04
光合作用在植物界甚至全球生态系统物质与能量循环中起极其重要作用,它是外因、内因共同综合作用的结果 [1]。光合作用是作物产量和品质构成的决定性因素,同时又是一个对环境条件变化十分敏感的的生理过程 [2-3]。米槁(Cinnamomum migao H W Li )为樟科樟属植物,树高可达 25 m,主干发达,侧枝较细弱,树冠近球形,树皮粗糙茶褐色,具纵向裂纹,内层及断面棕红色,分布于贵州省、云南省和广西壮族自治区等地 [4],垂直分布范围为海拔300~1 000 m [5]。米槁性甘、辛,味温,具有温中散寒、理气止痛的功效,主治胃痛、腹痛、风湿关节炎、胸闷和呕吐等症状 [6],是近年来贵州省拟重点发展的民族特色药材,以米槁为主要原料开发出的新药有心胃止痛胶囊、心胃止痛软胶囊、米槁心乐滴丸等 [7]。目前国内外对米槁的研究还很缺乏,主要集中在药用成分分离与鉴定 [8]、生物活性与药理特性 [9-10]、栽培技术与病虫害防治 [11-12]等方面,虽然取得了一定成绩,但产业化发展的技术基础依然薄弱,其中光合作用与其影响因子关系的研究报道鲜见。为此,以常规栽培的米槁一年生苗木为试验材料,对其光合作用日变化进行了研究,期望揭示米槁光合作用中净光合速率与生理生态因子间的关系,为米槁苗木培育、栽培管理提供科学依据和理论指导。
1 材料与方法
1 1 试验地概况
试验地位于贵州省贵阳市贵州大学南校区林学院苗圃内,平均海拔1 020 m,年平均气温为15 6 ℃,年极端最高温度为39 1 ℃,年极端最低温度为-7 8 ℃,最热月平均气温为24 ℃,最冷月(1月)平均气温4 1 ℃。年均降水量为 1 129 5 mm,年均空气相对湿度为71%,年均阴天日数为 240 6 d,年均日照时数为1 144 6 h。一年中气候温和,光照充足,无霜期较长。试验盆栽用土为黄泥土,呈微酸性,pH值为5 5~6 5,土壤肥沃。
1 2 供试材料
1 2 1 试验苗木 试验材料为米槁一年生实生苗。该苗木原位于2013年3月从贵州省罗甸县逢亭镇逢亭村米槁育苗基地移栽至贵州大学林学院苗圃温室大棚,种植于规格一致的塑料花盆(底部内径23 cm,盆口内径27 cm,盆高23 cm)内,而后进行精心的水肥管理,长势旺盛。9月中旬,在该批次的米槁苗中,选取光照条件下长势一致、无病虫害的3株米槁作为试验植株,每株选取中部的3张成熟叶,标记固定以备测定。
1 2 2 仪器与设备 光合作用测定仪器为美国LI-COR公司生产的Li-6400光合测定仪。
1 3 试验方法
米槁光合日变化数据在2013年9月中旬晴天测定,此时米槁平均株高0 8 m左右,生长旺盛。测定从07:00—18:00每隔1 h测定1次,一次测定选取3株共9张叶片。叶片连体测量时使用透明叶室、开放气路,空气流速为500 mL/min,待相关参数稳定后(约2~3 min),记录净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、大气CO2浓度(Ca)、气温(Ta)、叶温(Tleaf)、光合有效辐射(PAR)、叶片蒸汽压亏缺(VpdL)等参数,每张叶片连续记录3次,最终取平均值。
1 4 数据处理与分析
利用Excel 2007对实测数据进行图表处理,逐步回归分析和通径分析用SPSS 19 0完成。
2 结果与分析
2 1 环境生态因子日变化
光合有效辐射(PAR)(图1)及气温(Ta)(图2)一天中的变化均呈先升后降的趋势;而图3中空气相对湿度(RH)呈现先下降后上升的趋势,大气二氧化碳浓度则上下波动且变化幅度较小,早间最高,之后迅速下降并维持在385 μmol/mol 左右,在17:00之后又缓慢上升。其中,光合有效辐射和气温的峰值均是在14:00出现,而空气相对湿度于16:00左右降低至最低值,而且光合有效辐射和空气相对湿度日变化幅度较大。由图3可知,空气相对湿度(RH)早上较高(约55%),16:00左右最低(仅26%);叶片蒸汽压亏缺(VpdL)日变化与RH基本相反。
2 2 生理因子日变化
2 2 1 净光合速率与蒸腾速率日变化 米槁光合曲线呈典型的双峰型(图4),具有明显的光合“午休”现象。其中,07:00—10:00 急剧上升,10:00左右出现第1个高峰,达到 10 18 μmol/(m2·s);10:00—13:00净光合速率持续下降,至13:00左右出现谷值,此时净光合速率仅为第1次高峰时的26 50%;而后又缓慢上升,于15:00左右出现第2个高峰,峰[CM(25]值为7 93 μmol/(m2·s);在保持短时间的高峰后,于15:00—18:00 净光合速率迅速降至最低。比较2个峰值发现,下午的峰值仅为上午峰值的77 89%,一天中净光合速率的极差达8 93 μmol/(m2·s)。
米槁蒸腾速率(Tr)日变化(图4)同样为双峰曲线,早间随着Ta、Tleaf、PAR的升高以及RH的降低而逐步升高,11:00达到第1个峰值,略晚于叶片Pn峰值时间;随后Tr下降,在13:00左右出现低谷值,此时蒸腾速率仅为 2 55 mmol/(m2·s),之后逐渐升高,在15:00左右达到第2个峰值,最后随着Ta、Tleaf、PAR的降低和RH的升高而迅速下降。
2 2 2 胞间CO2浓度、气孔导度及叶温 图5显示,米槁气孔导度(Gs)的日变化与净光合速率相似,全天呈现双峰型变化 (峰值分别出现在10:00及15:00左右),15:00之后随光强的减弱而迅速下降。胞间CO2浓度(Ci)早间较高,随光合作用的迅速降低,而后变幅减小,傍晚17:00之后又迅速升高。米槁叶温(Tleaf)的变化与外界Ta基本一致,10:00—17:00 均处于30 ℃以上,最高温度出现在14:00前后,约为39 01 ℃。
2 3 影响净光合速率的生理生态因子分析
净光合速率是植物光合作用水平的指示指标,在植物光合作用的气体交换研究中扮演重要角色,它受外部生态因子和内部生理因子的双重影响,不但各个因子对净光合速率的影响较为复杂,且各因子间也存在交互影响 [13]。为此,采用多元逐步回归分析和通径分析相结合的方法将相关生理生态因子对米槁净光合速率的影响情况进行解析。
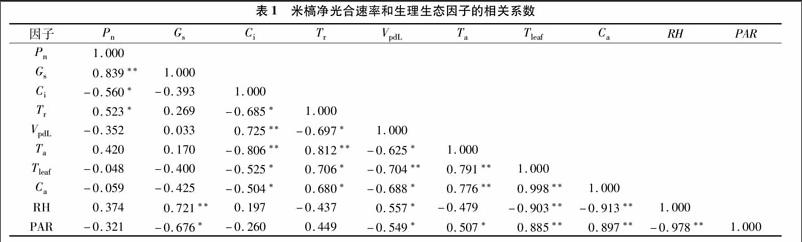
2 3 1 相关分析和多元逐步分析 从表1生理生态因子间的相关系数可以看出,Pn与Gs、Ci、Tr存在极显著或显著相关,而与PAR、Ta等因子相关性不显著。按照刘济明等 [14]的方法,把与米槁净光合速率Pn相关性显著的几个因子对Pn进行逐步回归分析,剔除部分变量,得到Pn对生理生态因子的回归模型:
y=21 257+100 982x1+0 168x2+0 067x3-0 056x4(R2=0 921)。 (1)
式中:y为Pn,x1为Gs,x2为Ci,x3为Tr,x4为Ca。
从式(1)可以看出,米槁净光合速率(Pn)主要受气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)以及大气CO2浓度(Ca)的影响,其他生理生态因子虽然也有影响,但在逐步回归分析中已被剔除。
2 3 2 通径分析 为进一步明确各生理生态因子对净光合速率的直接和间接影响,对测定结果进行通径分析,结果见表2。由表2可以看出,在生理因子中Gs、Ci、Tleaf、VpdL对Pn均起到了重要的作用,而在回归分析中并不重要的Tleaf[HJ1 9mm] 和VpdL也起到了重要的作用。 可能是因为这2个因子间存在着极强的负相关,使它们对Pn的影响在相关分析中被抵消,从表2中的间接效应亦可看出 [15]。生态因子中RH、Ta、Ca也起到了重要的作用,其中Ta的影响最大,但是由于Ta[JP+1]与其他因子间存在极强的正相关性,使它在相关分析中对Pn的影响被减弱。综合逐步回归分析和通径分析结果可知,影响米槁净光合速日变化的生理因子是气孔导度(Gs)和胞间CO2浓度(Ci),生态因子是大气CO2浓度(Ca)和相对湿度(RH)。
3 讨论
植物光合作用的日变化有单峰型、双峰型、严重型和平坦型4种类型 [16]。米槁叶片光合速率日变化呈双峰曲线,最高峰出现在10:00,为10 18 μmol/(m2·s),次高峰出现在 15:00,为7 93 μmol/(m2·s),午间有明显的“午休”现象。在高温强光天气下植物很容易出现“午休”现象,高温、强光和干旱土壤等条件引起的部分气孔关闭和光合作用光抑制是发生“午休”的主要原因 [17]。但对于不同的植物来说,引起光合速率“午休”的原因可能不同 [18-21]。关于引起光合速率产生“午休”的内在机制,大体上有2种解释:一种是气孔因素导致,由于午间太阳辐射增强,叶片细胞气孔阻力增大,CO2进入叶片受阻,胞间CO2浓度降低,光合速率下降,从而引起了植物出现“午休”现象 [22-24];另一种是由非气孔因素造成的 [25]。根据Farquhar等的观点 [26],判断气孔关闭是不是光合降低的原因,最重要的依据是胞间CO2浓度是否也同时降低,气孔和非气孔限制的界限是细胞间隙CO2浓度的变化方向,而不是气孔限制与非气孔限制值的相对大小。
本研究发现,在10:00以前,米槁叶片Ci随着Pn的增强而降低;在10:00—13:00时,Pn逐渐下降,Ci依然下降,同时发现Gs也随之下降,三者的变化趋势相一致,由此可以认为在此时间段内,Pn的变化主要是由气孔因素控制的。本试验同时发现米槁叶片Gs和Tr的日变化与叶片Pn日变化相似,表现出较强的正相关性。
植物光合作用是一个内外因子共同作用的复杂过程。在生产中,通过调节某个(某些)生理或生态因子提高树体光合作用时,要充分考虑调节因子对净光合速率所产生的直接和间接影响,以及因这些因子变化而导致其他相关因子发生变化所产生的负面影响 [27-28]。通过多元逐步回归分析和通径分析,结果表明影响米槁叶片Pn日变化的主要生态因子是大气CO2浓度和空气相对湿度,主要生理因子是气孔导度、胞间CO2浓度。由于减轻或避免“午休”现象是提高植物生长能力的一种方式 [29],因此在米槁苗木培育中,可考虑在日照过强、温度过高的午间,通过适当遮阴、加强通风等方式,减轻光合“午休”,促进生长发育,从而缩短苗木培育时间、尽早出圃。
参考文献:
[1] 姜小文,易干军,张秋明 果树光合作用研究进展[J] 湖南环境生物职业技术学院学报,2003,9(4):302-308,361
[2]孙存华,李 扬,贺鸿雁,等 藜的光合特性研究[J] 西北植物学报,2006,26(2):416-419
[3]林金科,赖明志,詹梓金 茶树叶片净光合速率对生态因子的响应[J] 生态学报,2000,20(3):404-408
[4]李永康 贵州植物志:第2卷[M] 贵阳:贵州人民出版社,1986:75
[5]赵 山,李鸿玉,邱德文,等 大果木姜子资源、生态调查——贵州、桂北及湘黔桂接壤区[J] 贵阳中医学院学报,1991(3):59-61,35
[6]李天祥,王静康 超临界CO2萃取大果木姜子精油的研究[J] 中药材,2003,26(3):178-180
[7]赵立春,邱明华,邱德文 超临界CO2萃取苗药大果木姜子果实挥发油化学成分研究[J] 环球中医药,2009,11(6):442-444
[8]武孔云,徐必学,梁光义,等 不同贮藏时间对米槁药材有效成分影响的比较研究[J] 时珍国医国药,2012,23(9):2 323-2 325
[9]孙学惠,隋艳华,邱德文 大果木姜子油对猫急性实验性心肌缺血的保护作用[J] 中国药学杂志,1995,30(6):341-344
[10] 桑维钧,李小霞,吴文辉,等 几种杀菌剂对米稿炭疽病菌的室内抑制试验[J] 青岛农业大学:自然科学版,2007,24(1):12-13
[11]江兴龙,潘俊锋,何茂琦 贵州米稿的栽培与病虫害防治技术[J] 林业调查规划,2005,30(4):104-108
[12]张小波,周 涛,郭兰萍,等 苗药大果木姜子挥发油成分变化及其地理分布[J] 生态学报,2011,31(18):5299-5306
[13]陈模舜,柯世省,倪琼琼 田间珊瑚树净光合速率及生态因子的日变化[J] 四川师范大学学报:自然科学版,2004,27(3):298-302
[14]刘济明,徐国瑞,廖小锋,等 罗甸小米核桃苗木光合作用特性[J] 西北农业学报,2012,21(3):148-152
[15]刘遵春,包东娥 “金光杏梅”叶片净光合速率与生理生态因子的关系[J] 西北植物学报,2008,28(3):564-568
[16]龙应忠,童方平,宋庆安,等 影响翅荚木种源苗期净光合速率的生理生态因子研究[J] 中国农学通报,2007,23(12):158-163
[17]Levitt J Rcsponse of plants to environment stress[M] New York:Academic Press,1972
[18]张江涛,刘友全,赵蓬晖,等 欧美杨无性系幼苗的光合生理特性比较[J] 中南林业科技大学学报,2007,27(4):8-11,22
[19]Aarnio E,Garr M,Hanohano J,et al Effects of drought on red maple(Acer rubrum) branch growth[J] Journal of Ecological Research,2002(4):31-36
[20]Bauerle W L,Whitlow T H,Setter T L,et al Abscisic acid synthesis in Acer rubrum L leaves - A vapor-pressure-deficit-mediated response[J] Journal of the American Society for Horticultural Science,2004,129(2):182-187
[21]谭会娟,周海燕,李新荣,等 珍稀濒危植物半日花光合作用日动态变化的初步研究[J] 中国沙漠,2005,25(2):262-267
[22]许大全 气孔的不均匀关闭与光合作用的非气孔限制[J] 植物生理学通讯,1995,31(4):246-252
[23]苏培玺,杜明武,张立新,等 日光温室草莓光合特性及对CO2浓度升高的响应[J] 园艺学报,2002,29(5):423-426
[24]路丙社,白志英,董 源,等 阿月浑子光合特性及其影响因子的研究[J] 园艺学报,1999,26(5):287-290
[25]张大鹏,黄丛林,王学臣,等 葡萄叶片光合速率与量子效率日变化的研究及利用[J] 植物学报,1995,37(1):25-33
[26]Farquhar G D,Sharkey T D Stomatal conductance and photosynthesis[J] Ann Rev Plant Physiol,1982,33:317-345
[27]唐建宁,康建宏,许 强,等 秦艽与小秦艽光合日变化的研究[J] 西北植物学报,2006,26(4):836-841
[28]黄春燕,吴 卫,郑有良,等 鱼腥草光合蒸腾特性及影响因素的分析[J] 西北植物学报,2006,26(5):989-994
[29]孙 嘏,刘洪剑,傅玉兰,等 江淮地区引种楸树光合生理特性的比较研究[J] 林业科学研究,2008,21(5):635-639
