籼粳交后代的籼粳分类及育性稳定性分析
2015-09-08林荔辉吴建梅林培清祁建民
林荔辉, 吴建梅, 林培清, 祁建民
(福建农林大学作物科学学院,福建 福州 35002)
籼粳交后代的籼粳分类及育性稳定性分析
林荔辉, 吴建梅, 林培清, 祁建民
(福建农林大学作物科学学院,福建 福州 35002)
以珍汕97B/秀水13的系列F7代株系为材料,在程氏籼粳分类与SSR分子聚类分析的基础上,鉴定了籼粳交后代株系在两种不同温度下的花粉黑染率与结实率的稳定性。结果表明,F7代株系的籼粳属性发生明显分离,以偏籼型比例最大,占47.7%;SSR分子聚类在遗传相似系数为0.54处将籼粳交后代大致分成籼、粳两大类群;SSR分子聚类结果与程氏籼粳分类结果基本吻合,相符度达85.4%。籼粳交后代花粉黑染率与结实率易受低温影响,育性下降,且变异系数大。根据育性表型可筛选出育性稳定的株系作为籼粳交育种材料。
水稻; 籼粳交; 程氏分类; 聚类分析; 育性稳定性
籼粳亚种杂种超亲优势强,有效利用其杂种优势已成为当前实现超级杂交稻育种目标的主要方向[1]。籼粳杂种一代不仅存在育性障碍,且结实率对环境变化较为敏感,温度是影响其育性稳定性的主要因素[2-4]。水稻灌浆结实的适宜温度一般在20-25 ℃左右,而籼粳杂种则需在更高的温度下方可正常孕穗结实,当温度<24 ℃就可能导致育性下降[5]。生产上常常因夏季温度突然降低而导致籼粳杂种明显减产,即使带有广亲和基因的籼粳杂种也易受影响[6],如广东、湖南、江苏、福建等省推广的亚优2号等均出现因低温导致结实率下降甚至不育,亚种间优势难以发挥[7]。因而,在籼粳亚种间杂种优势的利用上,既要考虑产量优势,还需兼顾其在不同地域的适应性与稳产性,而稳产的关键因素是结实率的稳定性[8]。
本研究选用偏籼型珍汕97B与偏粳型秀水13的杂交组合为育种群体,通过系谱法加代,筛选出株叶形态好、综合性状优良的系列F7代株系。在程氏籼粳分类法与SSR分子聚类相结合的基础上,分析了不同海拔高度与温度下籼粳类群的花粉黑染率与结实率的稳定性,为育种后代材料的选择与应用提供参考。
1 材料与方法
1.1 植物材料
以珍汕97B/秀水13的F2代为育种群体,通过系谱法育成性状稳定的41个F7代籼粳交后代。以5个籼型杂交稻(记为A5)、3个籼粳杂交稻(记为A6)、4个常规籼稻(记为A7)和3个常规粳稻(记为A8)为对照。除对照A6由三明市农业科学研究院提供外,其他材料均由福建农林大学作物科学学院提供。
1.2 程氏形态学指数籼粳分类
采用程氏形态指数分类法对F7株系进行籼粳属性分类[9]。根据分类分值判定各株系籼粳特性,即分值18-24为粳(记为A1),14-17为偏粳(记为A2),9-13为偏籼(记为A3),0-8为籼(记为A4)。
1.3 SSR分子标记聚类分析
根据McCouch et al[10]的SSR分子标记连锁图谱,在覆盖水稻整个基因组的SSR引物中,筛选出均匀分布在12条水稻染色体的76对SSR引物,对F7代株系进行聚类分析。SSR标记引物由上海生物工程技术服务有限公司合成。采用SDS方法提取水稻亲本与F7代株系DNA[11]。20 μL PCR反应体系为:(1)13.5 μL ddH2O;(2)2.0 μL引物;(3)2.0 μL Buffer(含Mg2+);(4)0.3 μL dNTPs;(5)0.2 μL Taq酶;(6)2.0 μL模板DNA。Taq酶、Buffer(含Mg2+)由福建农林大学作物科学学院配置。PCR反应设33个循环,每循环为:94 ℃变性5 min,94 ℃ 1 min、55 ℃ 1 min、72 ℃ 1min,最后72 ℃延伸10 min。PCR扩增产物采用6%非变性聚丙烯酰胺凝胶垂直电泳分离,根据引物片段大小调整电泳时间,经银染后显带分析。
1.4 育性稳定性鉴定
2009年7月1日、3日,分别在不同海拔高度下播种F7代株系与对照,其中顺昌县坂山村与莆田县西天尾镇的海拔高度分别为856、60 m(海拔每升高100 m,温度下降0.6 ℃,顺昌试点温度大约比莆田试点低4.8 ℃),以A5、A6、A7、A8为对照,考察籼粳交后代的花粉黑染率与结实率,分析A1、A2、A3、A4四类群在不同温度条件下的花粉黑染率与结实率差值,鉴定育性稳定性。待各株系全部抽穗后,分别于10月5日(低海拔)和6日(高海拔)早上8:00左右,每株系随机取3穗,每穗取当日5朵开放颖花,混合制片,采用1%I2-KI染色镜检,以黑染率的百分率考察育性,取平均值。在水稻成熟期,每份材料取5穗相同成熟度的稻穗考察结实率,取平均值。
1.5 数据统计
利用DPS软件分析籼粳交后代程氏形态学分类指数与6个性状间分值间的相关;采用Nei-Li类平均法进行SSR标记数据的聚类分析;并对比程氏形态学分类与SSR分子聚类分析结果。
2 结果与分析
2.1 程氏形态学指数分类
程氏形态学分类结果(表1)表明,对照常规粳稻、常规籼稻、籼型杂交稻以及籼粳杂交稻的程氏指数分值均对应于各自不同的类群。41个F7代株系的籼粳属性发生明显分离, 形态指数值呈连续分布。按籼型(0-8分)、偏籼型(9-13分)、偏粳型(14-17)、粳型(18-24)等4种类群划分,群体中出现了大量偏籼与偏粳的中间类型,籼∶偏籼∶偏粳∶粳的比例为7∶21∶10∶3,即21个株系偏籼型,占总株系的47.7%;而属性偏粳或粳的株系较少,分别为10个和3个,两者总和占总株系的31.7%,分离偏值可能与亲本秀水13程氏指数分值较低(分值仅14分)有关。从表1还可见,部分株系形态学明显偏粳,但分类却偏籼,反之亦然。上述偏差可能由于叶毛、稃毛等性状的表型分值具有较大的人为误差所致,这些株系的遗传背景则有待于采用更为精确的分子标记加以分析。
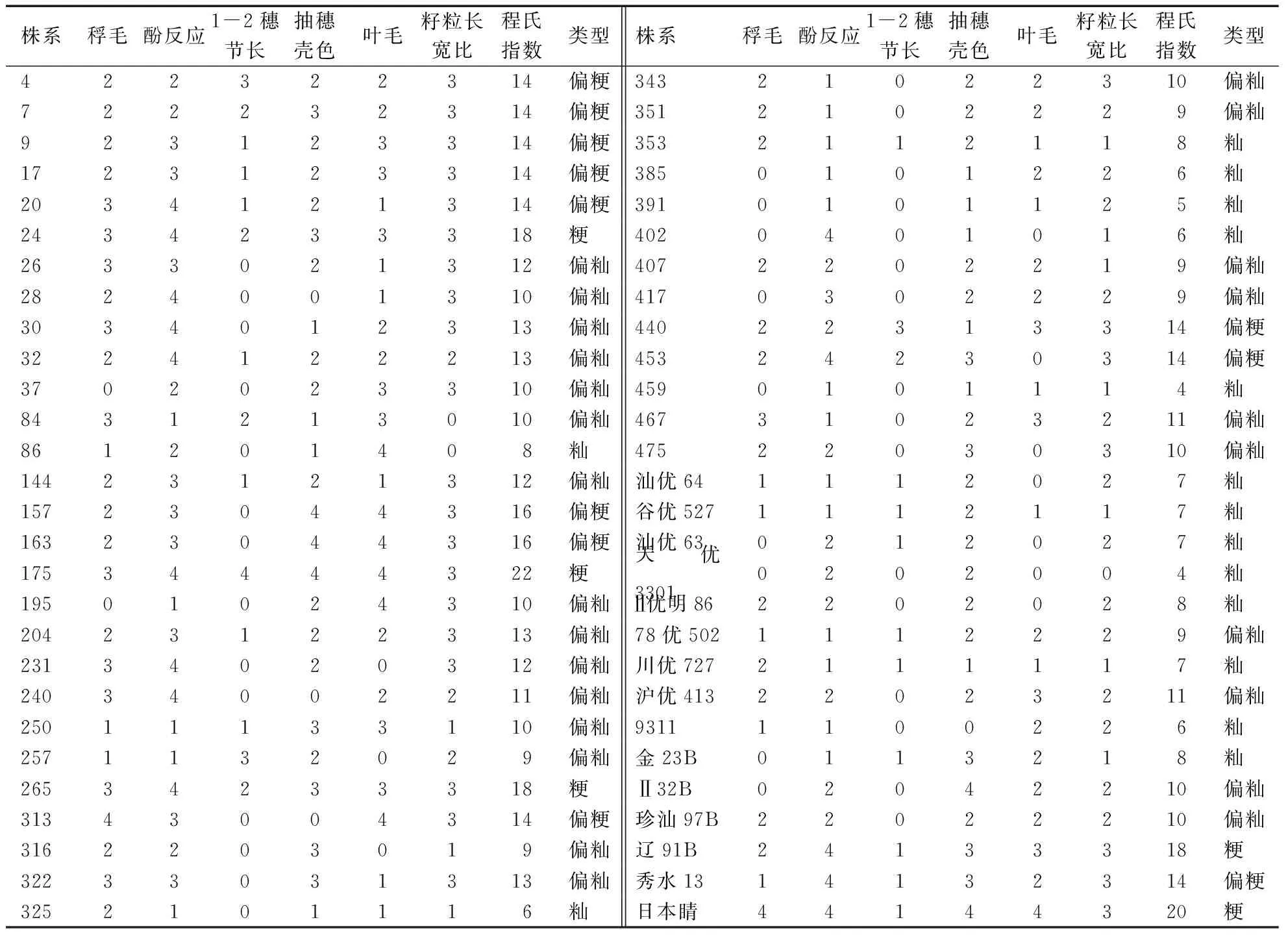
表1 珍汕97B/秀水13群体籼粳交后代的程氏形态学分类
41个株系和对照的程氏形态学指数与各性状分值的相关分析见表2。从表2可知,稃毛、酚反应、1-2穗节长、抽穗壳色、叶毛、籽粒长宽比等6个性状与程氏指数总分值之间均呈极显著正相关,表明程氏指数法中的6项判别性状指标都是区分籼粳类型的重要性状。其中与程氏形态指数相关系数最高的性状为籽粒长宽比和酚反应,相关系数达0.70**和0.69**,两性状对判断籼粳分类较为可靠。6个性状间除了酚反应与长宽比的相关系数为0.53**外,其他性状间的相关系数都较低,且大多性状间未达显著相关,表明上述6个判别性状在遗传上相互独立,其综合作用决定籼粳交后代的程氏分类指数与籼粳分类的表现。

表2 程氏形态学籼粳指数与6个性状分值间的相关分析1)
1)**表示差异达0.01显著水平。
2.2 SSR分子聚类分析
籼粳交后代株系经多代自交后,出现了明显的遗传变异。F7代株系的SSR分子标记聚类分析见图1。由图1可知,SSR标记能较好地区分粳稻和籼稻,且可直观表现出各株系间的遗传距离差异。41个F7代株系在遗传相似系数0.54处可分成两大类群,即粳稻类群(与秀水13同类群) 和籼稻类群(与珍汕97B同类群)。对比程氏形态学指数分类与SSR分子聚类结果,仅6个株系出现分类偏差,相符度达85.4%。其中在粳稻类群中,除了株系440(程氏籼粳分值14)聚类到籼稻类群中,其余12个粳或偏粳的株系均与秀水13聚类于同一类群,两种分类结果基本吻合;在粳稻类群中还聚类了26、28、30、144、204等5个偏籼型株系,上述株系的程氏籼粳分值分别为12、10、13、12和13,接近偏粳;酚反应分值分别为3、4、4、3、3,长宽比分值均为3分,两性状均倾向粳类群分值。故上述误差可能由于程氏分类时对叶毛、1-2穗节长、抽穗壳色等性状判别的人为误差所致。此外,鉴定结果也可能受外部自然环境影响产生一些微小的变化。综上分析表明,程氏形态学指数分类与SSR分子聚类对较为典型的籼粳特性分析是有效的,但对判断一些中间型材料的分类仍需加以完善。

图1 41个籼粳交后代的SSR分子聚类图
2.3 籼粳交后代育性稳定性分析
9月中下旬后,各供试材料均处于水稻的生殖生长期。逐日记载莆田低海拔试点的日平均温度变化,并依据两地温差(4.8 ℃)推算顺昌试点的高海拔温度变化情况(图2)。由图2可知,莆田试点在10月7日后出现<24 ℃的日平均温度, 11月2日后逐渐低于20 ℃;而在9月19日,顺昌试点日平均温度就出现<24 ℃,10月3日后已明显低于20 ℃。不同海拔间温度差异可有效鉴定出籼粳交后代的育性稳定性。
F7代株系类群程氏形态学指数平均分值分别为19.33(A1)、13.02(A2)、10.71(A3)和6.14(A4);而对照分值分别为6.60(A5)、9.00(A6)、8.50(A7)以及17.34(A8),类群内的变异系数变幅为11.95%-28.34%,籼粳分值变异大,说明籼粳交后代及对照具有典型性(表3)。

图2 2009年2种不同海拔高度的温度差异

类型籼粳分值CV/%低海拔黑染率/%CV/%结实率/%CV/%A119.33±2.3111.9595.67±4.935.1571.6±8.9012.43A213.02±3.6928.3487.80±26.0629.6776.24±22.0928.97A310.71±1.4813.8296.10±8.578.9275.86±12.1015.95A46.14±1.4623.7792.86±14.5315.7874.91±12.6816.93A56.60±1.5323.0397.00±3.934.0586.98±6.217.14A69.00±2.0022.2297.30±1.151.1875.87±6.438.48A78.50±1.9122.4798.75±0.500.5185.23±9.5611.21A817.34±3.0517.5998.67±0.570.5887.70±4.695.35类型高海拔黑染率/%CV/%结实率/%CV/%不同海拔下育性差值黑染率/%CV/%结实率/%CV/%A165.0±18.0327.7458.3±7.1512.2630.67±17.0155.4613.33±6.9552.14A278.97±23.7930.1366.86±20.7030.968.83±13.03147.569.38±11.31120.57A381.02±20.6025.4262.54±19.4231.0515.07±18.99126.0113.32±15.22114.26A485.21±17.5020.5361.44±12.8220.877.64±3.4244.7613.47±7.4555.31A591.01±6.527.1681.78±7.288.905.99±3.2454.005.20±2.1441.15A683.33±5.776.9262.57±6.6410.6113.97±5.1336.9113.3±11.6787.74A794.00±5.005.3282.55±9.3011.274.75±3.7779.372.68±0.269.70A891.67±2.893.1586.37±3.954.577.00±2.6437.711.33±0.8060.15
1)A1.粳型后代;A2.偏粳型后代;A3.偏籼型后代;A4.籼型后代;A5.籼稻杂交稻;A6.籼粳杂交稻;A7.籼稻常规对照;A8.粳稻常规对照。
从表3可见,所有供试材料在低海拔的花粉黑染率变幅为87.80%-98.75%,属正常可育,但籼粳交后代株系的花粉黑染率变异系数明显高于对照(A5-A8)。在高海拔条件下,籼粳交后代受低温影响的黑染率降幅明显高于对照,变幅为65.00%-85.21%,变异系数增大,育性稳定性差;而对照A5、A7、A8较为稳定,表明籼粳交后代易受低温影响,导致花粉黑染率明显降低。结实率变化趋势与花粉黑染率相似,低海拔下的籼粳交后代结实率均值明显高于高海拔下的结实率,两者差值变幅在9.38-13.47个百分点,变异系数在6.95%-15.22%之间;籼粳杂交稻(A6)差值为13.3个百分点,其他3组对照的结实率较为稳定,变幅仅为1.33-5.20个百分点,且变异系数小(0.26%-2.14%)。综上所述,籼粳交后代及籼粳杂交稻育性均易受低温影响,降幅大,育性稳定性明显低于对照(A5、A7、A8)。后代株系间在不同温度条件下花粉黑染率与结实率的稳定性也不同,其中7个株系的育性稳定性好。因而,这些株系是符合育种目标的中间材料,尤其是7、17、163等3个偏粳株系利用价值高。
3 讨论
籼粳间远缘品种杂交后代变异大,易创制出新的水稻资源,有效利用优异的中间材料,是实现杂交水稻超高产育种的有效途径。许旭明[12]研究表明,籼粳杂种优势利用存在双亲籼粳差异适度问题,即过大或过小的双亲遗传差异均不利于杂种优势的发挥。因此,判定与分析籼粳交后代的籼粳含量对育种材料的选择是必要的。采用偏籼型珍汕97B与偏粳型秀水13杂交群体的高代株系中,籼粳属性发生明显分离,形态指数值呈连续分布,多为偏籼型。这一结果与徐正进等[13]利用籼稻与偏粳型亲本杂交的F2代群体株系后代程氏指数变异结果相似。本研究中发现部分株系植株形态学偏粳型,但程氏形态学指数分类结果为偏籼型,SSR分子聚类分析则偏粳。由此表明,形态学标记只是一种表观的分类方法,在实际应用中可能存在一定人为误差,有效探讨其遗传基础,应采用SSR分子标记法加以鉴定。
低温可使水稻亚种间杂种的雌、雄配子发育受阻,花粉黑染率和结实率降低。亚种间开花期的安全日平均温度和适宜日平均温度一般比常规水稻的平均温度分别高2.0、1.2 ℃左右,因而,筛选鉴定亚种间杂种结实率的耐低温能力是籼粳交杂种优势利用的重要前提[14]。Li et al[15]对籼粳杂种育性的低温敏感性遗传分析表明,不同组合间的育性受低温影响程度大,且与其亲本的遗传背景密切相关,育性基因由一或几个基因共同控制,等位基因间的互作导致了在低温条件下的杂种不育性。李和标等[2]认为温度可以影响籼粳杂种的育性,当夏季日平均温度低于22 ℃时,亚种间杂种结实率明显降低。本研究中籼粳交后代育性也易受温度变化影响,与常规对照相比,低温条件下的水稻花粉黑染率与结实率明显降低,但也筛选出部分育性稳定性好的中间材料,并在籼粳杂交育种上取得明显进展[1]。当然,育性的耐冷性可能涉及诸多复杂的遗传机制和生理生化过程,如何有效克服籼粳交后代育性低温敏感性问题则有待进一步研究。
[1] 吴建梅,林荔辉,吴为人,等.水稻籼粳交亲籼型不育系的杂种优势利用[J].福建农林大学学报(自然科学版),2012,41(1):1-6.
[2] 李和标,李传国,陈忠明,等.籼粳杂种F1结实率稳定性研究[J].江苏农业学报,1995,11(3):7-11.
[3] 解平强,龙国炳,李卓吾,等.亚种间杂交稻F1代结实率的气候生态条件研究初报[J].湖南农业科学,1989(3):7-8.
[4] 段俊,梁承邺.亚种间杂交稻花粉育性和结实率的观察[J].广东农业科学,1996(1):7-9.
[5] 李祥洲,任昌福.水稻亚种间杂种一代籽粒充实的气温条件研究[J].作物学报,1996,22(2):247-250.
[6] LI H B, ZHANG Q, LIU A M, et al. A genetic analysis of low-temperature-sensitive sterility inindicajaponicarice hybrids[J]. Plant Breed, 1996,115:305-309.
[7] 杨杰,翟虎渠,王才林,等.水稻籼粳亚种间杂种低温花粉不育的QTL分析[J].遗传学报,2005,32(5):508-513.
[8] 陈立云.亚种间杂交稻结实率稳定性的研究[J].杂交水稻,2003,18(3):49-52.
[9] 程侃声.亚洲稻籼粳亚种的鉴别[M].昆明:云南科技出版社,1993:56-69.
[10] MCCOUCH S R, KOCHERT G, YU Z H, et al. Molecular mapping of rice chromosomes[J]. Theor Appl Genet, 1988,76:815-829.
[11] 吴建梅,林荔辉,林培清,等.水稻籼粳杂种育性QTL定位及其效应分析[J].热带作物学报,2014,35(2):246-252.
[12] 许旭明.水稻籼粳亚种间杂交衍生系遗传基础的研究[D].福州:福建农林大学,2009.
[13] 徐正进,李金泉,黄瑞冬,等.籼粳稻杂交后代亚种特性表现与分类研究[J].中国农业科学,2003,36(12):1571-1575.
[14] 吕川根,王才林,宗寿余,等.温度对水稻亚种间杂种育性及结实率的影响[J].作物学报,2002,28(4):68-73.
[15] LI H B, WANG J, LIU A M, et al. Genetic basis of low-temperature-sensitivesterility inindicajaponicahybrids of rice as determined by RFLP analysis[J]. Theor Appl Genet, 1997,95:1092-1097.
(责任编辑:陈幼玉)
Indica-japonicaclassification and fertility stability analysis ofindica×japonicacross progeny
LIN Li-hui, WU Jian-mei, LIN Pei-qing, QI Jian-min
(College of Crop Science, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China)
Based on Cheng′s morphological classification and SSR molecular clustering, the fertility stability of 41 F7lines derived from an original Zhenshan 97B (indica) × Xiushui 13 (japonica) cross was analyzed under two different temperatures. The results showed the following: 1) the F7lines were clearly segregating forindica-japonicaattributes, with a larger proportion (47.7%) beingindica-clinioustype; 2) the F7lines were divided intoindica- orjaponica-clinious groups at the genetic similarity coefficient value of 0.54; 3) there was a 85.4% agreement on results between SSR molecular clustering and Cheng′s index; and 4) the fertility of the F7lines, in terms of pollen stain rate and seed setting rate, was declined by exposure to low temperature with large coefficient of variation values. The fertility phenotype can be to screen for excellent breeding material inindica-japonicahybrid breeding program.
rice;indica-japonicahybrid; Cheng′s index; cluster analysis; fertility stability
2015-08-05
福建省教育厅资助项目(JA12096)。
林荔辉(1970-),男,副研究员,博士。研究方向:水稻遗传育种。Email:lihui9027@163.com。
S511
A
1673-0925(2015)03-0145-06
10.13321/j.cnki.subtrop.agric.res.2015.03.001
