水稻落粒性的分子生物学研究进展
2015-08-20宋颖娉宋立明
宋颖娉 宋立明
摘要:水稻是研究单子叶植物的模式作物,其落粒性的丧失是水稻被人工驯化的最为关键事件,近年来对水稻落粒性的研究取得了一系列进展,4个控制落粒性基因/主效QTL已经被克隆,并初步分析了相关基因的互作效应,为水稻人工驯化提供了重要依据。本文重点综述了水稻落粒性的分子机制研究进展,为水稻落粒性的遗传网络研究提供参考。
关键词:水稻;落粒性;驯化;分子机制;杂草稻
中图分类号: S511.01 文献标志码: A 文章编号:1002-1302(2015)07-0088-03
一般将亚洲栽培稻分为籼稻(indica)和粳稻(japonica) 2个亚种。关于亚洲栽培稻(以下简称栽培稻)的起源一直是一个困扰生物学家多年的难题,其中最主要的问题就是栽培稻是在哪里何时被驯化成栽培稻的[1-3]、水稻是一次性还是多次性从不同的祖先人工驯化而来的。与野生稻相比,落粒性的减弱、种子休眠性的降低、更紧凑的株型、花序变大且数目增多及种子变大是栽培稻驯化的综合特征。其中,落粒性的丧失是最重要的改变之一,这使得水稻成为易收获的作物[4]。如今,一般粳稻较难脱粒,籼稻居中,而野生稻[包括普通野生稻(Oryza rufipogon)及尼瓦拉野生稻(Oryza nivara)]易落粒[5]。
生产上中等难度落粒的品种最受欢迎,因为这样的品种对机械化操作最高效,也适用于手工收割和脱粒;容易落粒的品种,由于它们在收获中易引起减产而不受欢迎[6],所以在育种过程中,育种家倾向于选择中等落粒性的品种[7]。水稻落粒性的研究对揭示水稻进化机制及育种选择具有重要意义,水稻离区(abscission zone)发育和籽粒脱落的机制一直是分子生物学研究的热点之一。
1 水稻籽粒脱落的生理基础
水稻种子的落粒性受护颖和枝梗之间离层(abscission layer)所控制[8],离层在水稻抽穗前16~20 d就已形成发生。与周围枝梗和颖片细胞(大型厚壁细胞)组成不同,离层由 1~2 层小而圆的薄壁细胞组成[9]。当种子成熟时,离层细胞降解,使得水稻谷粒很容易地从母体植株上脱离下来。离层细胞可对激素产生响应,乙烯促进离层细胞脱离,而生长素则抑制脱离过程[10]。
不同的水稻品种具有不同的离层形态,这是导致水稻品种间落粒性存在差异的根本原因。野生稻的护颖和花梗间的接合处一般会形成离层,当中间薄壁细胞细胞壁发生降解时,成熟籽粒就会自由脱落,以保证种群繁殖[10]。而栽培稻的离层不完全形成,籽粒成熟后不会脱落,因而成熟种子可以得到有效收获。
2 水稻落粒性基因的发掘
水稻的落粒性是由多基因控制或由少数主效基因和多个微效基因共同控制的数量性状。利用各种遗传群体及其连锁图谱,许多学者对控制落粒性QTL 进行了定位,检测到了与落粒性相关的多个QTL。其中,在第1、3、4 染色体上检测到的QTL 的频率较高。这些控制落粒性的QTL 在野生稻(一年生或多年生)、杂草稻和栽培稻中都能检测到[2,5,11-14]。
同时,遗传学家也在积极寻找和创制水稻落粒性相关突变体,Ji 等通过使用化学诱变的方法在不落粒的粳稻品种Hwacheong中获得了1个易落粒突变体Hsh,发现Hsh 具有与尼瓦拉野生稻(O. nivara)类似发育良好的离层[15]。韩斌研究组使用一种巧妙的方法来发现新的水稻落粒调控基因,即将野生稻W1943的第4号染色体导入到栽培稻广陆矮4号背景下,构建了1个包含已知落粒基因SH4和qSH1的材料——SL4,该材料表现出极易落粒的性状。进一步对SL4进行射线诱变,筛选到2个完全不落粒的突变体shat1和shat2(与SH4等位),将其与已知的落粒调控基因联系起来,为水稻落粒性研究开启了新视野[16-17]。
3 水稻落粒性的分子生物学研究
到目前为止,在水稻中已经克隆了4个控制落粒性的基因/主效QTL,即SH4(SHA1、shat2)、qSH1、OsCPL1(sh-h)及SHAT1(SHATTERING ABORTION1)[4-6,12,17]。
Li 等利用籼稻品种和普通野生稻作亲本得到F2 群体,对水稻中的落粒性状进行QTL遗传分析,在水稻第4 染色体发现了1个贡献率为69%的主效QTL 位点,命名为SH4,该基因编码1个与Myb3同源的未知功能转录因子,该基因第1个外显子中的1个核苷酸替换(T替换了G)导致了在预测的Myb3 DNA 结合域上1个天冬酰氨替换赖氨酸,使得离层不能正常发育形成,从而使得表型从落粒变为了不落粒[4]。SHA1(shattering1)是SH4的等位基因,通过对96份粳稻和112份籼稻的检测,发现了它们均含有不落粒等位基因sha1,但在25 份野生的材料中均为野生型的SHA1[12]。后续研究证明对sh4的选择是一个独立的事件,即通过对该基因的一次选择而保留到水稻基因组上[18-19],支持了栽培稻不落粒的选择早于籼粳分化的理论。
尽管籼稻与粳稻均含有相同的SH4基因型,但这2个水稻亚种的落粒性却普遍有着显著的差别,通常粳稻难落粒而籼稻较易落粒,这暗示了水稻籼粳亚种间还存在其他主要控制落粒性的基因,qSH1就是其中的1个主效基因。qSH1基因编码1个BELl型同源盒基因,粳稻品种Nipponbare(日本晴)中qSH1基因5′调控区上的1个SNP(single nucleotide polymorphism)可引起离层无法形成,进而导致其落粒性丧失,qSH1能解释易落粒的籼稻品种Kasalath 与日本晴之间近70%的表型变异[5]。
Ji等从Hwacheong中获得易落粒突变体Hsh,命名为sh-h。sh-h基因位于水稻第7染色体,编码1个水稻carboxy-terminal domain (CTD) phosphatase类蛋白(OsCPL1),基因突变后功能失活促进离层的形成,导致易落粒。CTD磷酸酯酶在细胞分化中起重要作用,但是sh-h基因的功能及其与SH4、qSH1的互作机制还有待于进一步研究。
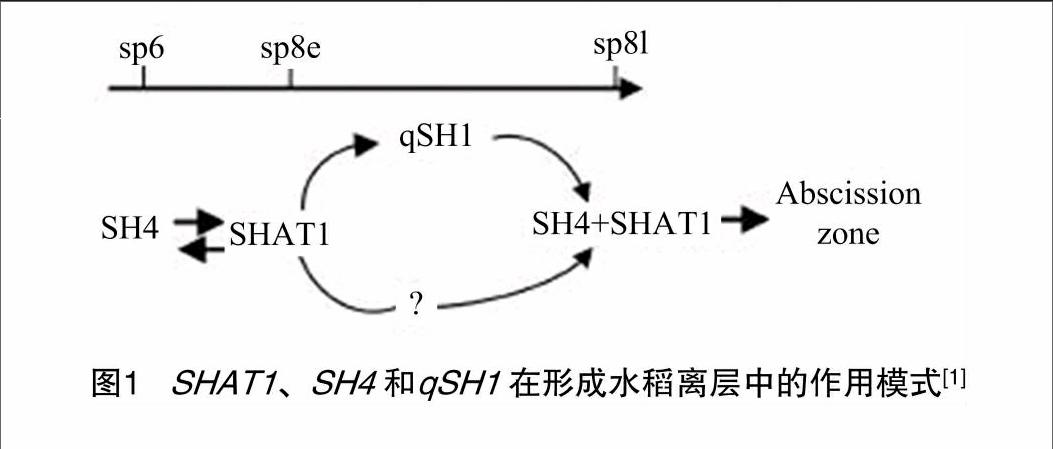
虽然SH4、qSH1在水稻的人工驯化过程中扮演着重要的角色,但是这2个基因的互作效应一直未能有较好的解释。韩斌课题组在一个包含已知落粒基因SH4和qSH1并表现出极易落粒的材料SL4诱变突变体中筛选获得1个极难落粒的材料shat1,SHAT1是一个未被人工选择的基因,类似于小麦的Q基因,SHAT1编码1个APETALA2转录因子[20],参与离层的发育,是落粒性必需基因。通过采用原位杂交等方法,研究人员第一次阐述了SHAT1、SH4和qSH1这3个基因的遗传关系:SHAT1 在离层的表达受SH4 正向调节,反过来SHAT1也起到维持SH4在离层表达的作用,qSH1作用于SH4和SHAT1下游,(还有可能有其他基因的参与)维持SHAT1和SH4在离层的持续表达,从而促进离层的形成(图1)[17]。
4 总结与展望
作物驯化是一个复杂而且漫长的过程[21-23],可以说作物的驯化史就是人类文明的进化史。随着考古的发现以及分子生物学的发展[23-25],越来越多关于水稻驯化的证据被发掘出来,最新研究已基本证明:栽培稻为单次起源[19],之后产生了籼粳分化,即野生稻首先在中国南方被驯化产生了粳稻,随后往北逐渐扩散。另一支则往南扩散,进入了东南亚,在当地与野生稻种杂交,经历了第二次驯化,产生了籼稻[26]。
20世纪以来,专业育种家的人工选择与有目的的基因重组加速了水稻进化的历史,特别是近30年来,随着科技的日新月异,以及在粮食危机的压力下,不同品种甚至亚种间的优异基因实现了高速的重组。然而,人们在有目的的筛选优良基因的同时,却导致了水稻基因型的单一化,当大规模的病害(或其他灾害)来临时,水稻变得十分脆弱。例如,随着劳动力的转移和科技的快速发展,轻简化的栽培方式兴起,导致易落粒的杂草稻繁殖加速,对水稻的生产带来诸多不利影响[27-28]。科学研究进一步对易落粒的杂草稻序列分析表明,在SH4位点,它们均含有与栽培稻相似的难落粒基因型(sh4)。这也带来了一个新问题:杂草稻具有与普通野生稻→亚洲栽培稻不相同的驯化历史?或者是除了目前已知的控制落粒性的基因/主效QTL外,还存在其他的主效基因[29-30]?
在双子叶模式植物——拟南芥中,已鉴定出多个与离区发育和器官脱落相关的基因,并已初步建立了拟南芥花器官脱落的调控网络[31]。水稻作为单子叶植物特别是禾本科作物的模式作物,到目前为止,只有4个对水稻离层发育所必需的落粒性基因被克隆。但是,它们之间的相互作用以及特别是OsCPL1基因的功能有待于深入研究,相关研究的匮乏难以解释水稻落粒性的多样性及复杂性。
最近,一个控制野生稻散穗的驯化基因SPR3(OsLG1)被克隆,SPR3编码1个SQUAMOSA 启动子结合蛋白(SBP)。研究发现该基因除了能够控制水稻的穗形外,还对落粒性有较大影响[32-33],这也更进一步说明水稻驯化是一个综合的过程,同时也预示着花器官发育相关基因对水稻的落粒性也起着调控作用。更多落粒性基因的发掘对更好地了解水稻落粒性的分子基础及现代育种具有重要的意义。
参考文献:
[1]Sang T,Ge S. The puzzle of rice domestication[J]. Journal of Integrative Plant Biology,2007,49:760-768.
[2]金 亮,卢 艳,包劲松. 水稻落粒性基因定位与克隆研究进展[J]. 分子植物育种,2009,7(2):393-397.
[3]区树俊,汪鸿儒,储成才. 亚洲栽培稻主要驯化性状研究进展[J]. 遗传,2012,34(11):1379-1389.
[4]Li C B,Zhou A L,Sang T. Rice domestication by reducing shattering[J]. Science,2006,311(5769):1936-1939.
[5]Konishi S,Izawa T,Lin S Y,et al. An SNP caused loss of seed shattering during rice domestication[J]. Science,2006,312(5778):1392-1396.
[6]Ji H,Kim S R,Kim Y H,et al. Inactivation of the CTD phosphatase-like gene OsCPL1 enhances the development of the abscission layer and seed shattering in rice[J]. The Plant Journal,2010,61(1):96-106.
[7]叶兴锋,徐林峰,施 聪,等. 中等落粒性的改良型超级稻 “协青早 A/M9308”应用价值评价[J]. 中国农学通报,2012,28(21):131-134.
[8]Patterson S E. Cutting loose. Abscission and dehiscence in Arabidopsis[J]. Plant Physiology,2001,126(2):494-500.
[9]Jin I D. On the formation and development of abscission layer in rice plants,Oryza sativa L.[J]. Jap J Crop Sci,1986,55:451-457.
[10]Roberts J A,Elliott K A,Gonzalez-Carranza Z H. Abscission,dehiscence,and other cell separation processes[J]. Annual Review of Plant Biology,2002,53:131-158.
[11]Cai H W,Morishima H. Genomic regions affecting seed shattering and seed dormancy in rice[J]. Theoretical and Applied Genetics,2000,100(6):840-846.
[12]Lin Z W,Griffith M E,Li X R,et al. Origin of seed shattering in rice (Oryza sativa L.)[J]. Planta,2007,226(1):11-20.
[13]Qin Y,Kim S M,Zhao X H,et al. Identification for quantitative trait loci controlling grain shattering in rice[J]. Genes & Genomics,2010,32(2):173-180.
[14]Sun J,Qian Q,Ma D R,et al. Introgression and selection shaping the genome and adaptive loci of weedy rice in northern China[J]. The New Phytologist,2013,197(1):290-299.
[15]Ji H S,Chu S H,Jiang W Z,et al. Characterization and mapping of a shattering mutant in rice that corresponds to a block of domestication genes[J]. Genetics,2006,173(2):995-1005.
[16]Hofmann N R. SHAT1,A new player in seed shattering of rice[J]. The Plant Cell,2012,24(3):839.
[17]Zhou Y,Lu D F,Li C Y,et al. Genetic control of seed shattering in rice by the APETALA2 transcription factor shattering abortionl[J]. The Plant Cell,2012,24(3):1034-1048.
[18]Zhang L B,Zhu Q H,Wu Z Q,et al. Selection on grain shattering genes and rates of rice domestication[J]. The New Phytologist,2009,184(3):708-720.
[19]Molina J,Sikora M,Garud N,et al. Molecular evidence for a single evolutionary origin of domesticated rice[J]. Proceedings of the National Academy of Sciences of the United States of America,2011,108(20):8351-8356.
[20]Simons K J,Fellers J P,Trick H N,et al. Molecular characterization of the major wheat domestication gene Q[J]. Genetics,2006,172(1):547-555.
[21]Doebley J F,Gaut B S,Smith B D. The molecular genetics of crop domestication[J]. Cell,2006,127(7):1309-1321.
[22]Vaughan D A,Lu B R,Tomooka N. The evolving story of rice evolution[J]. Plant Science,2008,174(4):394-408.
[23]Fuller D Q,Qin L,Zheng Y F,et al. The domestication process and domestication rate in rice:spikelet bases from the Lower Yangtze[J]. Science,2009,323(5921):1607-1610.
[24]Huang X H,Wei X H,Sang T,et al. Genome-wide association studies of 14 agronomic traits in rice landraces[J]. Nature Genetics,2010,42(11):961-967.
[25]樊龙江,桂毅杰,郑云飞,等. 河姆渡古稻DNA提取及其序列分析[J]. 科学通报,2011,56(增刊2):2398-2403.
[26]Huang X H,Kurata N,Wei X H,et al. A map of rice genome variation reveals the origin of cultivated rice[J]. Nature,2012,490(7421):497-501.
[27]戴伟民,宋小玲,吴 川,等. 江苏省杂草稻危害情况的调研[J]. 江苏农业学报,2009,25(3):712-714.
[28]梁帝允,强 胜. 我国杂草稻危害现状及其防控对策[J]. 中国植保导刊,2011,31(3):21-24.
[29]Thurber C S,Reagon M,Gross B L,et al. Molecular evolution of shattering loci in U.S. weedy rice[J]. Molecular Ecology,2010,19(16):3271-3284.
[30]Zhu Y Q,Ellstrand N C,Lu B R. Sequence polymorphisms in wild,weedy,and cultivated rice suggest seed-shattering locus sh4 played a minor role in Asian rice domestication[J]. Ecology and Evolution,2012,2(9):2106-2113.
[31]王 翔,陈晓博,李爱丽,等. 植物器官脱落分子生物学研究进展[J]. 作物学报,2009,35(3):381-387.
[32]Luo J J,Hao W,Jin J,et al. Fine mapping of Spr3,a locus for spreading panicle from African cultivated rice (Oryza glaberrima Steud.)[J]. Molecular Plant,2008,1(5):830-838.
[33]Ishii T,Numaguchi K,Miura K,et al. OsLG1 regulates a closed panicle trait in domesticated rice[J]. Nature Genetics,2013,45(4):462-465.
