冲突适应过程中的神经信息传递机制
2015-08-03赵晓月唐丹丹
赵晓月?唐丹丹
摘要:在认知控制研究领域中,冲突适应(conflict adaptation)涉及大脑对冲突进行连续的认知调整,它能反映人类有效控制冲突的基本机制。然而,现有研究还没有充分地揭示冲突适应的脑机制。本研究通过对被试完成字母Flanker任务时的脑电(electroencephalography,EEG)数据进行有效连通性分析,揭示了冲突适应过程中的神经信息传递机制。在观察任务中,当大脑传递冲突信号时,从左前额叶电极点到中前区电极点的连通性(β频带:20~21 Hz,400~650 ms)增强,以使大脑顺利完成与反应抑制相关的加工;在反应任务中,从中前区电极点到右前额叶电极点,congruency-Congruency条件比incongruency-Congruency条件的连通性(β频带:18~21 Hz,600~1,000 ms)更强。这些结果共同说明β频带的活性调整过程反映了大脑有效的执行控制过程,从而在行为上表现出由冲突观察所诱发的冲突适应。
关键词:冲突观察;冲突适应;侧边干扰任务;脑电;有效连通性分类号B84511引言
在认知控制领域中,冲突适应是人类大脑对连续的冲突情境进行有效控制的基础。它反映了一个由冲突所诱发的连续的行为调整过程,此行为调整便导致了优化的行为(Botvinick,Braver,Barch,Carter,& Cohen,2001; Botvinick,Nystrom,Fissell,Carter,& Cohen,1999; Gratton,Coles,& Donchin,1992)。采用一致性任务,如:Flanker任务(Eriksen & Eriksen,1974)、Stroop任务(Stroop,1935)和Simon任务(Simon,1969),冲突适应这一现象的认知和神经机制已被广泛地研究。如在字母Flanker任务中,刺激是由中间的靶刺激和两侧的干扰刺激组成,个体需要根据中间的字母(靶刺激)作出反应,同时忽略两侧的字母(干扰刺激)。根据靶刺激和干扰刺激的相容性,可将该任务划分为一致条件(congruent,简称C条件)和不一致条件(incongruent,简称I条件)两种条件。在一致条件下,靶刺激和干扰刺激相同(如“KKKKK”);在不一致条件下,靶刺激和干扰刺激不同(如“FFWFF”)。以反应时(RT)为行为指标,RT(I-C)即Flanker干扰效应(Eriksen & Eriksen,1974)。为研究冲突适应,在整个任务系列中,如果根据先前试次和当前试次的一致性(一致、不一致),将一致和不一致条件进一步划分为cC、cI、iC和iI试次(cC代表一致试次之后的一致试次,cI代表一致试次之后的不一致试次,iC代表不一致试次之后的一致试次,iI代表不一致试次之后的不一致试次),那么冲突适应则表现为冲突试次之后的干扰效应显著地小于一致试次之后的干扰效应,即RT(iI-iC) 目前,由Botvinick等(2001)提出的冲突监测理论能有力地解释冲突适应的认知神经机制。他们将冲突适应过程区分为对冲突出现的监测和对冲突的控制两个阶段,并且分别将冲突监测与前扣带回(anterior cingulate cortex,ACC)、冲突控制与背外侧前额叶(dorsal lateral prefronal cortex,DLPFC)相联系。根据该理论,在认知控制过程中存在着从ACC到DLPFC的联结环路(Nee,Jonides, & Berman, 2007; Nee, Wager, & Jonides, 2007)。大脑的冲突监测结构(ACC)和冲突解决结构(DLPFC)通过自上而下的信息交流来实施对行为冲突的控制和适应。先前试次中的冲突信息被ACC监测到之后,ACC迅速地将此信号传递给负责冲突控制的脑区DLPFC,DLPFC接收到冲突信号后,使大脑在当前试次出现之前便处于积极的准备状态。在当前试次中,当冲突再次出现时即能实现对冲突的有效控制,从而提升当前试次的行为表现(di Pellegrino,Ciaramelli,& Ladavas,2007; Egner & Hirsch,2005a,2005b; Floden,Vallesi,& Stuss,2011; Kerns,2006; Kerns et al.,2004; Liston,Matalon,Hare,Davidson,& Casey,2006; Matsumoto & Tanaka,2004; Silton et al.,2010; van Veen & Carter,2006)。最近的磁共振(functional magnetic resonance imaging,fMRI)研究结果进一步为冲突适应提供了高空间分辨率的证据(Sheth et al.,2012)。 由于冲突适应涉及连续而实时的认知调整(Kerns et al.,2004; Mansouri,Tanaka,& Buckley,2009),那么采用高时间分辨率(精确到毫秒)的脑电(electroencephalography,EEG)技术就可以揭示出冲突适应的精确时间进程。近年来,有效连通性分析(effective connectivity analysis)已被运用于人类的认知研究中(Cohen & Cavanagh,2011; Cohen & van Gaal,2013; Hwang,Velanova,& Luna,2010)。它为直接讨论不同脑区之间信息传递的因果关系提供了一个有效的方法(Friston,Harrison,& Penny,2003),其结果能揭示高水平的认知信息传递随时间变化的基本机制(Sauseng & Klimesch,2008)。有效连通性算法基于格兰格因果关系概念(Granger,1969),它能有力地评估神经元激活之间因果关系的方向和强度。一般地,有效连通性通过时间序列数据的多元自回归(multivariate autoregressive,MVAR)模型来检测,其结果通常表示为频域尺度,即局部定向相干性(partial directed coherence,PDC) (Baccala & Sameshima,2001)和直接传递函数(directed transfer function,DTF) (Kaminski & Blinowska,1991)。特别地,由于冲突适应涉及不同脑区之间的信息传递(例如:从ACC到DLPFC自上而下的信息交流)(Kerns et al.,2004),并且反映了人类大脑对认知的时间序列调整,那么对EEG数据进行有效连通性分析,其结果将揭示出这种序列调整的基本脑机制。
最近的研究以健康成人为被试,在时频域上发现,诱发的β振荡同时缩减了运动的速度和力量(Joundi,Jenkinson,Brittain,Aziz,& Brown,2012; Pogosyan,Gaynor,Eusebio,& Brown,2009)。也有研究显示,对于GO/noGO任务和停止信号任务(stop signal task),在成功的noGO试次和停止信号试次中,分别由人类下丘脑神经元(subthalamic nucleus,STN)和右侧额下回所触发的β频带的活性有所增长(Kühn et al.,2004; Swann et al.,2009)。这些结果共同表明,β振荡的能量调整是人类认知和动机系统中的一种常见机制,它的活性与反应抑制或冲突解决有关。
目前,在冲突观察范式中(Tang,Hu,& Chen,2013; Tang,Hu,Li,Zhang,& Chen,2013; 唐丹丹,刘培朵,陈安涛,2012; 唐丹丹, 陈安涛,2012),只有唐丹丹等人采用Stroop任务直接考察了冲突适应过程中不同脑区信息传递的因果关系。他们对时频EEG数据进行了有效连通性分析,其时频分布结果展示了由冲突观察所诱发的对冲突的适应。对于观察任务,从右前额叶(right-frontal scalp region)到后顶部(posterior parietal scalp region),不一致条件比一致条件的信息流动更强,该信息流反映在β频带(beta-band,20~21 Hz,300~500 ms和700~900 ms)上。此结果可能表明,右前额叶-后顶部β频带的激活与冲突监测有关:在观察任务中,与冲突监测相关的信息能被有效地从右前额叶传递到后顶部,以确保大脑为将要执行的反应任务做准备(Engel & Fries,2010)。对于反应任务,从中顶部(centro-parietal scalp region)到右前额叶,iI条件比cI条件的信息流动更强,该信息流反映在θ频带(theta band,6~7 Hz,180~330 ms)上。此结果可能表明,大脑对冲突信息的评估和对控制的调整可能与θ频带的活性调整有关。因此,冲突适应所涉及的冲突监测和冲突控制过程能被反映在不同脑区的有效连接上。
然而,在基于字母Flanker任务所设计的冲突观察范式中,现有研究还没有清楚地展示冲突适应的有效连通性机制。本研究采用冲突观察实验范式(Tang et al.,2013; Tang,Hu,Li,et al.,2013; 唐丹丹,等,2012; 唐丹丹,陈安涛,2012),考察冲突解决过程中不同脑区之间的信息传递机制。研究记录了15个正常成人被试在完成字母Flanker任务时的行为和EEG数据。其中,EEG数据通过EEGLAB (Delorme & Makeig,2004)预处理后,被进一步地进行有效连通性分析。此方法能有力地评估不同脑区之间信息传递的方向和强度(Hu,Zhang,& Hu,2012)。因此,本研究结果能在时频域上展示人类大脑解决冲突过程中的信息传递机制。
2方法
本研究的实验设计与唐丹丹和陈安涛(2012)等人的相同。为了进一步考察人类解决冲突过程中的神经信息传递机制,本研究采用有效连通性分析方法(Hu et al.,2012)对其EEG数据重新进行分析。
2.1被试
15名20~23岁的大学生(其中女生8名)自愿报名参加本次实验,平均年龄22.77岁(SD=0.70)。被试均为右利手,视力或矫正视力正常,在此之前没有参加过类似的实验,对本次实验目的毫不知情。实验完成后付给一定报酬。
2.2实验仪器与材料
实验采用计算机呈现刺激,计算机的显示器为联想LX-GJ556D,17寸彩显,分辨率为1024×768,颜色为真彩色,刷新率为85 Hz,屏幕背景为黑色。实验程序由E-prime编制运行,刺激呈现时间、反应时均由计算机自动记录。被试距屏幕的距离约为60 cm,在亮度适中的单间实验室里单独参加测试。刺激由白色的注视点(+或
瘙 毐 )及四类白色的大写字母组成(K、N、F、W)。字母用22号Times New Roman字体呈现,字母之间的空隙是一个空格的宽度。刺激的长和宽分别为5 cm和1.2 cm(视角:4.77 °×1.15 °)。每个试次中包含一个注视点和五个水平排列的大写字母,中间的字母是目标(靶)刺激,两侧的四个字母是干扰(分心)刺激,包含一致(如“KKKKK”)和不一致(如“FFNFF”)两种条件。
2.3实验程序
实验流程见唐丹丹和陈安涛(2012)的研究。首先在屏幕中央呈现300 ms的注视点,然后呈现时间间隔在300~500 ms内随机变化的空屏,之后呈现五个水平排列的大写字母,直到被试在1,500 ms内做出反应;反应之后呈现时间间隔在800~1,200 ms内随机变化的空屏,然后进入下一试次。为了使被试熟悉反应规则,在正式实验前,他们需完成一个包含64个试次(随机排列)的练习block,刺激和反应都与正式实验一致。正式实验包含6个block,每个block各包含145个试次。这些试次被伪随机地排列,使得从观察条件到反应条件转换时,cC、cI、iC和iI试次各80个,观察条件和反应条件各320个试次,并且前后两个试次中没有刺激重复和反应重复(避免特征整合效应)的试次。为了排除反应定势的影响,在刺激序列中随机地插入连续的观察试次或反应试次。
在整个任务中,如果呈现的注视点是“瘙 毐 ”,则提示被试将出现观察试次(观察刺激,但不需要作出反应);如果呈现的注视点是“+”,则提示被试将出现反应试次(观察刺激,并且做出反应)。实验要求被试采用主键盘,根据中间的字母(忽略两旁的字母)快速且准确地做出反应:如果中间的字母是K,则用左手中指按“1”键;如果中间的字母是N,则用左手食指按“2”键;如果中间的字母是F,则用右手食指按“9”键;如果中间的字母是W,则用右手中指按“0”键。完成每个block后休息两分钟,整个实验持续一小时左右。
2.4脑电记录
采用国际10~20系统扩展的64导电极帽,以Brain Products GmbH记录脑电信号,参考电极置于左侧乳突。同时,为了避免左右半球不对称性的情况,在右侧乳突上也放置电极,接地点在FPz和Fz的中点。左眼上下安置电极记录垂直眼电(VEOG),双眼外侧安置电极记录水平眼电(HEOG)。每个电极处的头皮电阻小于5 kΩ。滤波带通为0.01~100 Hz,采样频率为500Hz/导,离线式(off-line)叠加处理数据。
2.5EEG数据处理与统计分析
EEG数据预处理。EEG数据采用EEGLAB (Delorme & Makeig,2004)进行预处理。连续的EEG采用30 Hz的低通过滤,0.1 Hz的高通过率。EEG数据的分析时程从刺激呈现前500 ms到刺激呈现后1,000 ms,以先于刺激的时间间隔(-500 ms到0 ms)为基线校正数据。肌电(electromyographic,EMG)伪迹、放大器饱和所引起的伪迹以及其他伪迹(如,波幅超过±80 μV的伪迹)通过手动去除。眨眼和眼动伪迹用独立成分分析(independent component analysis,ICA)算法校正(Delorme & Makeig,2004; Jung et al.,2001; Makeig,Jung,Bell,Ghahremani,& Sejnowski,1997)。在所有数据集中,个别被移除的独立成分(independent components,ICs)包括一个大的眼电的电极贡献和前额叶的头皮分布。然后,再次进行基线校正。最后建新参考,即以所有电极的平均值为新参考。
定义空间感兴趣区域(spatial regions of interest,S-ROIs)。由于本研究在左前额叶电极点[left frontal region:(F1+F3)/2]、右前额叶电极点[right frontal region:(F2+F4)/2]和中前区电极点[centro-frontal region:(FCz+Cz)/2]观察到最明显的与任务相关的调节,所以这三个脑区被定义为S-ROIs。
有效连通性分析。采用时变的有效连通性(time-varying effective connectivity)(Hu,et al.,2012)评估大脑左前额叶、右前额叶和中前区之间信息流动的因果关系。最近Hu等(2012)发展了这种时变的有效连通性分析方法,本研究所采用的算法和Hu等人的算法相同。第一,采用时变的多元自回归(timevarying MVAR,tvMVAR)模型描绘单试次的大脑响应的演变,并且采用Kalman平滑确定tvMVAR模型。研究表明,Kalman平滑能准确地估计tvMVAR系数。第二,有效连通性模式在时频域上以时变的部分定向相干性(timevarying PDC,tvPDC)的形式展示出来。在观察任务中,对于一致条件和不一致条件,tvPDC值从Kalman基于平滑的tvMVAR系数估计中计算出来(Baccala & Sameshima,2001)。在反应任务中,本研究采用相同的算法分别计算cC,cI,iC,iI条件下的tvPDC值。第三,采用引导统计分析(bootstrapping statistical analysis)评估不同条件之间tvPDC值的显著性,此显著性水平的阈值设定为p<.01。再从1到30 Hz以1 Hz的步调对tvPDC求值,并且通过减法对tvPDC值进行基线校正,然后在每个被评估的频率上再除以被包含在基线时间间隔(从-400 ms到-100 ms)内tvPDC的平均值。第四,在观察任务中,采用时变的有效连通性评估左前额叶和中前区之间信息传递的因果关系。对于不一致条件和一致条件中所得的tvPDC值,本研究采用配对样本t检验比较二者的差异。在反应任务中,采用相同的方法评估右前额叶和中前区之间信息传递的因果关系。为评估冲突适应,本研究采用两因素(观察任务的一致性:一致、不一致;反应任务的一致性:一致、不一致)重复测量方差分析(analysis of variance,ANOVA)比较cC、cI、iC、iI条件之间tvPDC值(PDC100%)的差异,事后配对比较采用Bonferroni法校正。
3行为及EEG结果
3.1行为结果
为评估由冲突观察所诱发的冲突适应,对于RT,本研究实施了2 (观察任务的一致性:一致、不一致)×2(反应任务的一致性:一致、不一致)的两因素重复测量ANOVA。结果发现,观察任务的一致性主效应显著,F(1,14)=7.90,p<.05,η2=0.36;反应条件的一致性主效应显著,F(1,14)=55.41,p<.001,η2=.80;观察任务一致性与反应任务一致性的交互作用显著,F(1,14)=543,p<.05,η2=.28。事后检验仅仅比较iI和cI条件,以及cC和iC条件的RT。结果仅仅发现了iI条件和cI条件的RT差异,RTiI
对于错误率,本研究同样地实施重复测量ANOVA。由于观察任务的一致性与反应任务的一致性交互作用不显著,所以,在错误率上没有展示出显著的冲突适应,F(1,14)=1.65,p>.05,η2=0.11。其中,cC、cI、iC和iI条件下的错误率分别为380%、527%、540%和573%。
3.2观察任务中的神经信息传递过程
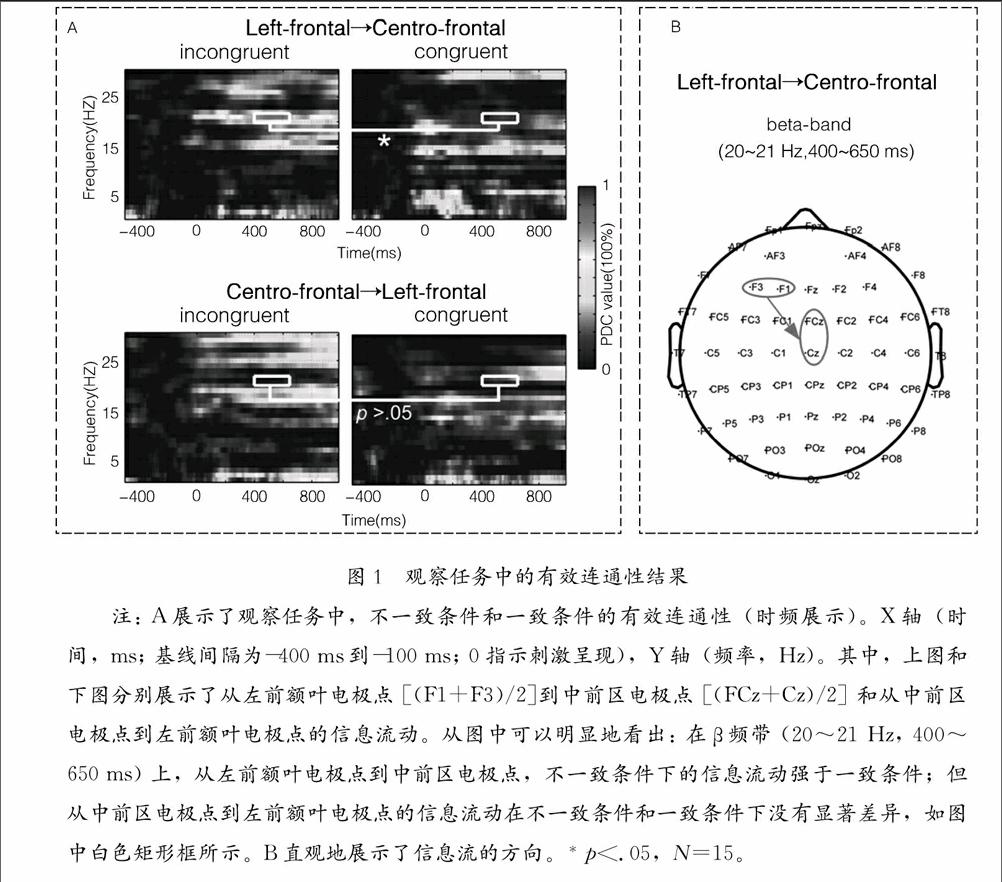
下页图1展示了观察任务中的有效连通性结果。对于观察任务,在不一致条件和一致条件下,分别采用时变的有效连通性确定了左前额叶电极点和中前区电极点之间信息传递的方向和强度:对相对于基线时间间隔(-400 ms到-100 ms,防止边缘效应)内的tvPDC值(p<.01; FDR校正; 自展检验,bootstrap analysis)求和。而在其他ROIs之间的有效连接上,没有观察到与观察任务有关的显著效应,ps>.05。图1A展示了左前额叶电极点和中前区电极点之间显著增加的tvPDC值(Milde et al.,2010)的时频分布。从左前额叶电极点到中前区电极点,不一致条件的有效连通性(β频带:20~21 Hz,400~650 ms)显著强于一致条件的有效连通性,t(14)=2.17,p<.05。其中,不一致条件和一致条件下的tvPDC值(平均值±标准差,PDC100%)分别为:0.65±0.87和0.11±0.51。采用相同的分析方法,本研究在其他频带上没有发现不一致条件和一致条件之间tvPDC值的显著差异,ps>.05。从中前区电极点到左前额叶电极点,不一致条件和一致条件之间的有效连通性(β频带:20~21 Hz,400~650 ms)差异都不显著,p>.05。图1B直观地展示了信息流传递的方向:神经信息有效地从左前额叶电极点传递到中前区电极点。图1观察任务中的有效连通性结果
注: A展示了观察任务中,不一致条件和一致条件的有效连通性(时频展示)。X轴(时间,ms; 基线间隔为-400 ms到-100 ms; 0指示刺激呈现),Y轴(频率,Hz)。其中,上图和下图分别展示了从左前额叶电极点 [(F1+F3)/2]到中前区电极点 [(FCz+Cz)/2] 和从中前区电极点到左前额叶电极点的信息流动。从图中可以明显地看出: 在β频带 (20~21 Hz, 400~650 ms) 上,从左前额叶电极点到中前区电极点,不一致条件下的信息流动强于一致条件;但从中前区电极点到左前额叶电极点的信息流动在不一致条件和一致条件下没有显著差异,如图中白色矩形框所示。B直观地展示了信息流的方向。
3.3反应任务中的神经信息传递过程
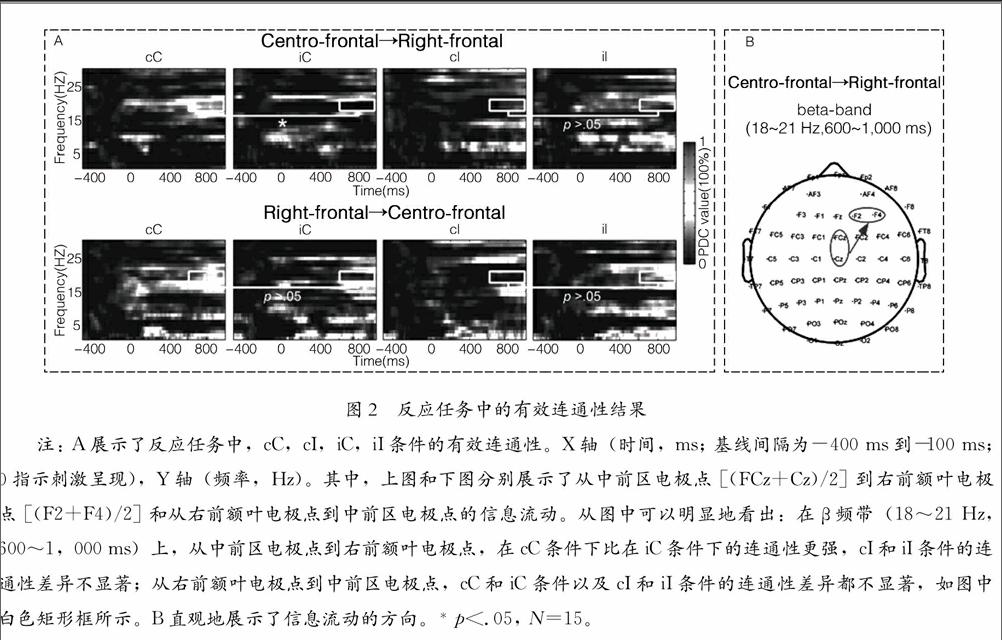
下页图2展示了反应任务中的有效连通性结果。图2A展示了右前额叶电极点和中前区电极点之间显著增加的tvPDC值(FDR校正) (Milde et al.,2010)的时频分布。从中前区电极点到右前额叶电极点,只有β频带(18~21 Hz,600~1,000 ms)展示了与冲突适应相关的效应。其中,观察条件的一致性主效应边缘显著,F(1,14)=4.06,p=.06,η2=.23;反应条件的一致性主效应不显著,F(1,14)=1.49,p>.05,η2=.10。重要的是,观察条件与反应条件的一致性展示了显著的交互作用,F(1,14)=9.38,p< .01,η2=.40。其中cC、cI、iC和iI 四种条件下的tvPDC值(平均值±标准差,PDC100%)分别为:0.67±091、0.09±0.32、0.02±0.33和0.19±0.44。为评估冲突适应,事后检验只比较cC和iC条件、cI和iI条件之间tvPDC值的差异(Cohen & Cavanagh,2011; Larson,Kaufman,& Perlstein,2009; Tang,Hu,Li,et al.,2013)。结果显示:cC条件的tvPDC值显著大于iC条件的tvPDC值,p<.05;cI条件和iI条件的tvPDC值没有显著差异,p>.05。然而,从右前额叶电极点到中前区电极点,两因素重复测量ANOVA没有展示任何显著性结果,ps>.05。另外,在其他ROIs之间的有效连接上,没有观察到与反应任务有关的显著效应, ps>.05。图2B直观地展示了信息流传递的方向:神经信息有效地从右前额叶电极点传递到中前区电极点。图2反应任务中的有效连通性结果
注: A展示了反应任务中,cC,cI,iC,iI条件的有效连通性。X轴(时间,ms; 基线间隔为-400 ms到-100 ms; 0指示刺激呈现),Y轴(频率,Hz)。其中,上图和下图分别展示了从中前区电极点 [(FCz+Cz)/2] 到右前额叶电极点 [(F2+F4)/2] 和从右前额叶电极点到中前区电极点的信息流动。从图中可以明显地看出:在β频带(18~21 Hz,600~1,000 ms)上,从中前区电极点到右前额叶电极点,在cC条件下比在iC条件下的连通性更强,cI和iI条件的连通性差异不显著;从右前额叶电极点到中前区电极点,cC和iC条件以及cI和iI条件的连通性差异都不显著,如图中白色矩形框所示。B直观地展示了信息流动的方向。
4讨论
本研究的有效连通性结果为冲突适应过程提供了不同脑区之间信息传递的直接证据。冲突监测可能主要发生在左前额叶,冲突适应则主要发生在右前额叶;从冲突监测到冲突适应,中前区起到了调控作用。这与以脑损伤病人和动物为被试的研究结果一致(di Pellegrino et al.,2007; Markela-Lerenc et al.,2004; Posner & DiGirolamo,1998; Roelofs,van Turennout,& Coles,2006),这些研究发现前额叶在冲突监测过程中起重要作用,中前部(ACC)主要负责对冲突进行控制和调整。本研究结果也与近来的一项事件相关电位(ERPs,event-related potentials)研究结果一致,Tang,Hu,Li等(2013)采用相似的实验设计发现,在右前额叶,持续电位(sustained potential,SP)的波幅调整反映了大脑对冲突的适应。
时频域的研究证实,β频带的活性与当前的认知激发状态成反比(Engel & Fries,2010):β频带的激活与反应抑制有关,其失活与反应激活或反应准备有关(Pasttter, Hanslmayr, & Buml,2008,2010)。并且β频带的失活反映了实际的或想象中的运动活动(Kilner,Bott,& Posada,2005; Pasttter et al.,2008; Tzagarakis,Ince,Leuthold,& Pellizzer,2010)。一些研究也发现β频带与大脑对反应的加工有关(Alegre et al.,2003; Kaiser,Birbaumer,& Lutzenberger,2001; Kilner et al.,2005; Pasttter et al.,2008; Tzagarakis et al.,2010; Zhang,Chen,Bressler,& Ding,2008)。近来的一项研究在人类下丘脑神经元植入深度大脑刺激电极(deep brain stimulation electrodes,DBS),记录了被试完成色-词Stroop任务时的神经活动。结果发现,由人类下丘脑神经元所触发的β振荡与反应抑制有关,并且在β频带范围内的能量调整对于反应选择是至关重要的(Brittain et al.,2012)。在不一致条件下,β活性短暂增长,这就推迟了行为反应,以至于大脑解决了冲突。
本研究的结果显示,在观察任务中,大脑左前额叶将监测到的冲突信息(β频带:20~21 Hz,400~650 ms)传递到中前区,使之调整认知资源,以实施对冲突的控制。这一过程共持续250 ms,这一时间间隔正好与大脑从冲突的监测到实施对冲突的控制所需的最短时间相对应(Müller & Rabbitt,1989; Notebaert,Gevers,Verbruggen,& Liefooghe,2006)。另外,本研究也发现,在不一致的观察条件下,β频带的活性更强,说明在不一致条件下,大脑执行更强的反应抑制加工以成功地对冲突进行控制,这一控制过程主要通过左前额叶到中前区自上而下的信息交流来实现(见图1)。
在反应任务中,从中前区电极点到右前额叶电极点,cC条件比iC条件的有效连通性(β频带:18~21 Hz,600~1,000 ms)更强,但是cI条件与iI条件之间没有显著差异。这说明冲突适应也能体现在当前的一致试次上,这与冲突适应模型一致(Freitas,Bahar,Yang,& Banai,2007; Gratton et al.,1992; Lamers & Roelofs,2011; Ullsperger,Bylsma,& Botvinick,2005)。对于iC条件,由于被试在先前的观察任务中监测到冲突信息,那么大脑已经集中了更多的资源加工靶刺激而非侧边(Flanker)刺激,所以缩小了注意范围(Paquet,2001)。而在其后的反应任务中,在一致条件下,由于没有冲突出现,那么注意范围的缩小将不利于任务的完成。然而,对于cC条件,由于被试在先前的观察任务中没有监测到冲突信息,那么大脑将拓展注意范围,并且更多地加工侧边(Flanker)刺激;在其后的反应任务中,在一致条件下,由于没有冲突出现,那么注意范围的拓宽将更利于任务的完成。另外,对于iI和cI条件,尽管大脑在观察任务中的注意范围不同,但是大脑能根据反应任务所面临的冲突情景及时地调整认知资源加工靶刺激,从而顺利解决当前的冲突,所以cI条件与iI条件之间的有效连通性强度没有显著差异。然而,在Stroop任务中,Tang等(2013)发现了从中顶部电极点到右前额叶电极点显著的有效连通性,即iI条件比cI条件的信息流动更强,该信息流反映在θ频带(6~7 Hz,180~330 ms)上。本研究没有观察到这一结果,这可能是由于任务类型不同造成的,因此还需要将来的研究进一步证实。
综上所述,本研究认为:在被试完成字母Flanker任务时,大脑左前额叶能监测到观察任务中的冲突信息,并且将冲突信息有效地传递到中前区;在反应任务中,一方面中前区能根据观察条件的一致性环境灵活地调整注意资源以有效地完成反应任务,另一方面它再向右前额叶传递控制信息,使之及时解决冲突(Floden et al.,2011; MacDonald,Cohen,Stenger,& Carter,2000),从而表现出由冲突观察所诱发的冲突适应(见图2)。
5结论
大脑左、右前额叶和中前区的信息流动能影响β频带的活性。在观察任务中,当从左前额叶到中前区传递冲突信息时,β频带的活性增强,并且这种信息流动影响了反应任务中的信息流动强度和方向。比起iC条件,连续的一致性情境(cC条件)导致大脑分配更多的资源加工Flanker刺激,那么大脑便从中前区向右前额叶传递更少的冲突信息,所以β频带的激活更强。上述这些联合的效应导致由冲突观察所诱发的冲突适应。
