应用ROC分析猕猴掌跖骨性差
2015-07-17赵哲王凤产胡凤霞田华香赵晓进
赵哲+王凤产+胡凤霞+田华香+赵晓进

摘 要 为用ROC分析猕猴掌跖骨的性差大小,取成年猕猴掌骨标本44例(14雄,30雌),跖骨标本45例(14雄,31雌).选择掌跖骨长度和骨干横径2个变量进行测量.数据分析采用SPSS 20.0软件.统计处理采用逻辑回归和ROC分析.结果表明:两侧掌跖骨长度性差较大,ROC曲线下面积(AUC)变化范围为0.917~0.994;侧别之间的性别差异较小;掌跖骨长度的性差明显大于骨干横径性差;ROC分析显示5根掌跖骨的长度性差大小不同,两侧的掌骨性差明显大于跖骨.结果提示:猕猴掌跖骨性差显著,掌跖骨的长度变化主要受基因控制,骨干横径主要受到环境因素的影响.掌跖骨性差模式可能与人类明显不同,可能很好地反映了遗传与环境因素的交互作用.
关键词 猕猴;掌跖骨;性差;ROC分析
中图分类号 Q954.61 文献标识码 A 文章编号 1000-2537(2015)03-0029-05
由于灵长类骨骼化石非常稀少,目前主要是根据对现生灵长类的骨骼进行研究,然后将其结果应用于灵长类化石的研究中[1].利用部分骨骼变量来推断个体性别的技术日趋成熟,包括使用一些较小的骨骼,例如锁骨[2]、跟骨[3]、掌骨和跖骨[4-5].
本文研究目的是通过逻辑回归模型和ROC分析方法来比较猕猴掌跖骨的各个变量之间、左右侧别之间、5根掌跖骨之间以及掌骨与跖骨之间的性差大小.目前有关掌跖骨的性差研究国外资料较多[6-9],而国内资料较少[4-5];采用单因素和多元回归分析较多[10-12],采用ROC分析和逻辑回归分析较少[6];掌骨研究较多[11-12],跖骨研究较少[5].本研究可积累非人灵长类骨骼性差数据,丰富国内非人灵长类掌跖骨性差资料,为灵长类化石性别鉴定或种属识别奠定基础.
1 材料和方法
猕猴掌跖骨标本来自河南省北部的太行山猕猴自然保护区.
成年掌骨标本44例(14雄,30雌),跖骨标本45例(14雄,31雌).标本的性别是已知的,年龄根据牙齿萌发和掌骨骨骺愈合情况来确定.所有标本是左右成对.选择健康无病、形态发育正常的掌跖骨标本进行测量.选择掌跖骨的最大长和骨干横径进行测量[4,6],测量工具采用0~150 mm电子数显卡尺,精度:0.01 mm.骨骼标本保存在河南师范大学生物标本馆.
为了便于描述,掌跖骨的缩写字母分别用MC和MT表示,根据它们的排列顺序从拇指到小指依次标识为MC1(MT1),MC2(MT2),MC3(MT3),MC4(MT4)和 MC5(MT5).掌跖骨最大长为LG,掌跖骨中间横径为MW.这里选择MW是因为根据资料它们可以作为骨干粗壮度的指标[13],左右侧别标识是在掌跖骨缩写字母前面加上“L”或“R”.例如,缩写字母为:LG(LMC3),表示左侧第三掌骨的最大长.
所有测量数据采用SPSS 20.0进行分析.
ROC分析通常用于临床疾病诊断.ROC曲线能很容易地查出测试组的临界值[14].可将各个测试组的ROC曲线绘制到同一坐标中,分别计算各个测试组的ROC曲线下面积(AUC),哪一组的AUC最大,说明哪一组的诊断价值最佳.在比较掌骨和跖骨的性差大小时,首先用5根掌骨长度和5根跖骨长度做二元逻辑回归,将得到的回归结果做ROC分析.便于整体上比较掌跖骨的性差大小.
2 结果
2.1 两侧掌跖骨性差
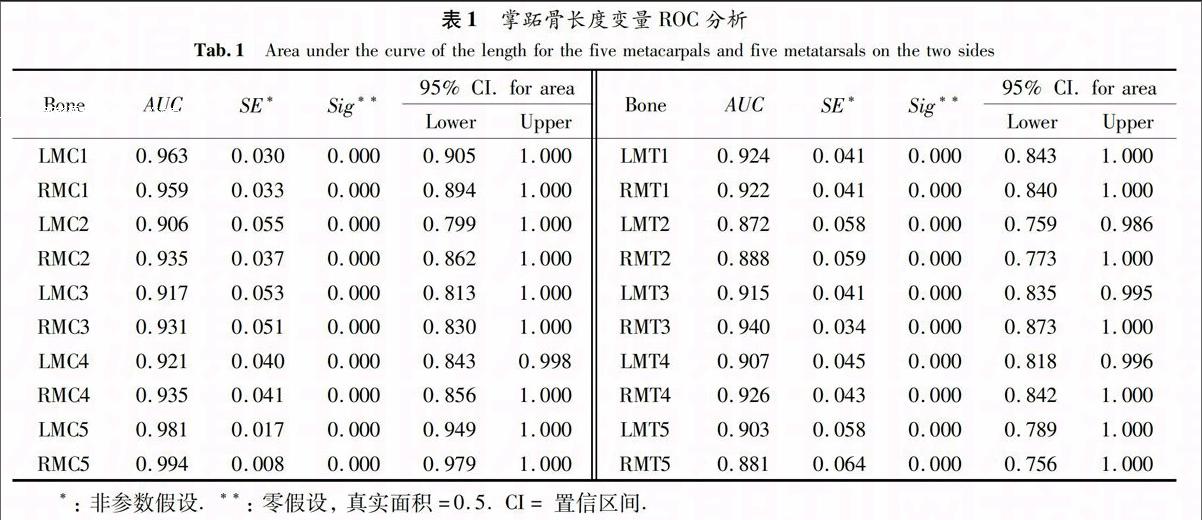
5根LG(MC)性差的侧别比较见表1(左半部分)和图1(上半部分).结果表明:总体上ROC曲线下的面积(AUC)较大,变化范围为0.906~0.994,侧别之间的AUC差别较小.5根LG(MT)性差比较见表1(右半部分)和图1(下半部分).结果表明:总体上比掌骨性差小一些,AUC的变化范围为0.872~0.940;除了RMT3(AUC=0.940)外,都是MC的AUC大于MT的AUC.掌跖骨长度的性差在左右两侧之间差异较小.
2.2 掌跖骨LG与MW性差
由于左右侧掌跖骨变量的差异较小,统计采用左右侧掌跖骨变量合并计算.比较结果见图2(A,B).很明显无论是掌骨还是跖骨,LG的性差明显大于MW.其他的掌跖骨的LG与MW比较结果与MC3和MT3类似,不再赘述.
2.3 5根掌跖骨性差
用ROC分析5根LG(MC)的性差大小,结果见图2C.LG(MC5)的性差最大(AUC=0.968),最小的是LG(MC2)(AUC=0.917).5根LG(MT)性差大小见图2D.LG(MT3)的性差最大(AUC=0.926),最小的是MT2(AUC=0.881).图3 左侧(A)和右侧(B)掌跖骨长度性差ROC分析(虚线:掌骨,实线:跖骨)
2.4 掌骨与跖骨性差
将5根掌骨LG和5根跖骨LG逻辑回归结果进行ROC分析.结果显示无论是左侧还是右侧,掌骨性差明显大于跖骨性差(见图3).本文得到的AUC较大主要是由于采用了5根掌骨LG和5根跖骨LG多元逻辑回归的结果.
3 讨论
结果显示太行山猕猴掌跖骨变量性差显著,证明通过逻辑回归和ROC分析可以有效地识别个体的性别.许多文献支持此结果[9,15],如McFadden等[15-16]报道了狒狒、大猩猩和黑猩猩掌跖骨相对长度的性差.结果证明掌跖骨长度的性差主要受遗传基因的控制,同时也受出生前的胚胎发育环境和出生后性激素释放的影响.
3.1 两侧掌跖骨性差
ROC分析结果显示侧别之间的AUC没有显著性差异.原因可能是:(1)两侧掌跖骨发育是否对称取决于运动类型和日常行为活动的影响,两侧掌跖骨LG无性差,说明在日常的行为习惯上没有差异.这个结果与Barrio等[17]报道的雅典人和Kusec等[18]报道的克罗地亚人掌骨性差研究结果一致.他们两侧掌骨具有很高的对称性,推测他们两手的使用是一致的.(2)猕猴在本质上不存在人类所特有的利手现象[19],所以人类与猕猴两侧掌跖骨性差模式可能不同.有资料显示人类右手具有更大的性差[20].Alicioglu等[13]认为由于人类存在利手现象,所以在研究掌骨时应严格限制在左手,目的是尽量减少一些行为活动对性差的影响.(3)在一些大型的猿类是否在种群水平上存在利手现象仍存在较多的争议[20].Sarringhaus等[19]研究了黑猩猩的的MC2是否存在侧别差异.结果表明MC2虽然是右侧大于左侧,但是没有达到统计学意义.(4)采用不同的统计方法和不同的掌跖骨变量结果可能有所不同,但是总体上掌跖骨侧别差异较小,不会像人类那样显著.掌跖骨的形态主要取决于力量控制和精确协调,包括会受到同侧和交叉肢体长骨的影响[20].(5)本文结果提示采用掌跖骨进行性别鉴定时,如果发现一侧骨骼增生、变形或缺失,可以采用另一侧的骨骼来替代,一般不会影响统计结果.另外,根据文献报道,为了增加统计结果的稳定性和样本数目,可以在统计时将两侧的掌跖骨标本进行合并[6].
3.2 掌跖骨LG和MW性差
许多研究结果表明掌跖骨的不同变量性差不同[4-6].Scheuer等[10]对英国人掌骨上不同变量的性差大小进行研究,结果显示掌骨MW的性差最大,其次是MC底宽和头宽,LG的性差最小.Khanpetch等[6]用ROC分析研究了泰国人的掌骨性差.结果显示MW(LMC3)性差较大(AUC=0.843).LG(LMC3)性差较小(AUC=0.790).本文研究结果显示掌骨LG性差明显大于跖骨MW.提示人类和猕猴掌跖骨变量的性差模式可能不同.结果分析:(1)Sarringhaus等[19]报道人类和非人灵长类掌骨的形态存在明显差异,这是由行为活动的差异造成的,例如,运动类型、行走姿态、抓握精度及准确度等.(2)骨骼长度的生长主要受遗传基因的影响,掌骨MW等变量反映了掌骨的粗壮度特征,主要受到后天行为活动的影响[11].(3)掌跖骨的LG和MW都存在一定程度的性差,都可以单独使用来辅助鉴定个体的性别,如果结合一起使用,性别判别效果会更好[4-5].(4)一般情况下,掌跖骨LG具有生长周期较短、容易测量及性差显著的特点,很少受到环境机械刺激或年龄因素的影响,所以在人类研究中也是性别判别的最佳变量和最常用变量.(5)人类与猕猴掌跖骨变量性差模式不同,主要归结于遗传、环境和行为活动等因素的交互作用.
3.3 5根掌跖骨性差比较
总体上5根掌跖骨之间的性差是不同的,但是差别较小.猕猴掌跖骨长度性差从大到小的排序:MC5,MC1,MC4,MC3,MC2;MT3,MT1,MT4,MT5,MT2.这与人类5根掌跖骨之间性差大小的报道不同[20],甚至与某些非人灵长类的掌跖骨研究结果也不同.McFadden等[15]报道了大猩猩和黑猩猩掌跖骨长度统计,5根掌跖骨性差从大到小排序分别是:MC2,MC5,MC3,MC4,MC1;MT5,MT4,MT2,MT3,MT1.总体上黑猩猩的掌跖骨性差明显小于大猩猩.他们同时指出,无论是大猩猩还是黑猩猩,可能都和人类一样,掌跖骨在早期生长发育时明显受到性激素的调控[15].Case等[20]根据美国人掌跖骨标本得出结果显示LG(MC3)性差最大,其次是LG(MC2),最小的是LG(MC1);他们认为:(1)LG(MC1)性差较小可能与他们的绝对值较小有关,另外可以解释为在LG(MC1)的使用中性别之间没有明显差异;(2)掌跖骨LG性差显著,可以有效地识别个体的性别,但是不一定是性别鉴定的唯一指标或最佳选择.以前许多文献都证明了这一点,例如人类(包括一些非人灵长类)远侧段指骨的性差常常大于掌骨[20].人类掌骨性差最大的是MW[20];另外掌骨LG性差明显小于掌骨体积的性差[22];无论对于人类还是非人灵长类,在用掌跖骨变量进行性别判别时,常常是LG和其他粗壮度变量结合使用,效果更好[20].(3)掌跖骨的绝对LG性差与他们相对LG性差大小相比,相对值性差可能会比绝对值性差具有更大的应用价值[15-16].因为相对长度的性差不受个体形态大小或变量大小的影响,而且很有可能与胚胎早期发育和一些疾病的发生有密切的联系.文献显示出生前胚胎环境比出生后环境因素对骨骼长度的影响更小.但是如果出生前环境中性激素水平异常,就会导致掌跖骨长度早期发育出现异常[8].
3.4 掌骨与跖骨性差比较
总体上,猕猴掌骨性差大于跖骨.Case等[20]对欧美人掌跖骨长度的性差进行研究,结果显示掌骨性差大于跖骨,但未解释产生这一差异的原因.McFadden等[16]对狒狒的掌跖骨LG和质量采用效应值来计算性差大小,结果发现掌骨LG的效应值是3.0~3.7,跖骨LG的效应值是4.1~4.9.表明非人灵长类跖骨性差大于掌骨.如果采用相对LG进行性差比较,则LG(4MT)∶LG(5MT)性差最大.Tague[9]报道了17种非人灵长类掌跖骨LG的性差,发现一些灵长类的掌跖骨LG性差非常显著,也有一些种属掌跖骨LG没有性差.他们报道了食蟹猴(Macaca fascicularis)掌跖骨LG的性差大小,结果显示5根掌跖骨的性差都有统计学意义(P<0.001).5根掌骨LG与所对应的5根跖骨LG性差几乎相等.赵晓进等[23-24]研究显示猕猴掌跖骨LG性差模式不同,认为主要与猕猴掌跖骨的生活习性、使用频率、活动范围和灵活程度等有关.还有一种掌跖骨性差大小不同的解释是由于跖骨的形态具有明显的弓形结构,所以用X射线测量时,没有像测量掌骨LG那么准确[5].本文采用直接测量法,且跖骨的弯曲程度并不是很明显,所以不支持上述解释.掌跖骨性差模式不同可能是猕猴掌跖骨的性差特征之一.综上所述,作者认为影响掌跖骨性差大小的因素主要有统计方法、种属差异、运动类型、体位、行为习惯和使用频率等.
总之,研究灵长类掌跖骨的性差可能会为动物种属之间的相互关系提供丰富信息,促进掌跖骨在其他灵长类和哺乳动物研究中更深入地应用.
参考文献:
[1] 吴汝康.灵长类的性别二态问题[J].人类学学报, 1987,6(3):255-261.
[2] 宋 洁,王凤产,毛晓静,等. 太行山猕猴锁骨性差[J].河南师范大学学报, 2014,42(1):129-132.
[3] 赵俊杰,赵晓进,党晓云,等. 太行山猕猴跟骨性别判别[J]. 解剖学杂志, 2009,32(5):687-689.
[4] 胡海洋,赵晓进,周 迪,等.太行山猕猴掌骨性别判别分析[J].人类学学报, 2013,32(2):226-232.
[5] 闫鑫甜,张弯弯,徐小解,等.猕猴蹠骨性别逐步判别分析[J].解剖学杂志, 2012,35(5):670-674.
[6] KHANPETCH P, PRASITWATTANSEREE S, CASE DT, et al. Determination of sex from the metacarpals in a Thai population[J]. Forensic Sci Int, 2012,217(1-3):2291-2298.
[7] EI MORSI D A, AL HAWARY A A. Sex determination by the length of metacarpals and phalanges: X-ray study on Egyptian population[J]. J Forensic Leg Med, 2013,20(1):6-13.
[8] MCFADDEN D, BRACHT M S. Sex difference in the relative lengths of metacarpals and metatarsals in gorillas and chimpanzees[J]. Horm Behav, 2005,47(1):99-111.
[9] TAGUE R G. Variability of metapodials in primates with rudimentary digits: Ateles geoffroyi, Colobus guereza, and Perodicticus potto[J]. Am J Phys Anthropol, 2002,117(3):195-208.
[10] SCHEUER J L, ELKINGTON N M. Sex determination from metacarpals and the first proxinal phalanx[J]. J Forensic Sci, 1993,38(4):769-778.
[11] MANOLIS S K, ELIOPOULOS C, KOILIAS C G, et al. Sex determination using metacarpal biometric data from the Athens Collection[J]. Forensic Sci Int, 2009,193(1-3):130.e1-130.e6.
[12] SMITH S L. Attribution of hand bones to sex in population groups[J]. J Forensic Sci, 1996,41(3):469-477.
[13] ALICIOGLU B, YILMAZ A, KARAKAS H M, et al. Sex determination by interarticular distance of metacarpals and phalanges: a digital radiologic study in contemporary Turkish people [J]. J Anat, 2009,3:14-20.
[14] 范莎莎,李爱民,李 荣,等.IL-17在肝癌患者血清中表达增高对肝癌诊断的研究[J].湖南师范大学自然科学学报, 2014,37(3):19-23.
[15] MCFADDEN D, BRACHT M S. Relative lengths of fingers and toes in human males and females[J]. Horm Behav, 2002,42(4):492-500.
[16] MCFADDEN D, BRACHT M S. The relative lengths and weights of metacarpals and metatarsals in baboons (Papio hamadryas)[J]. Horm Behav, 2003,43(2):347-355.
[17] BARRIO P A, TRANCHO G L, SANCHEZ J A. Metacarpal sexual determination in a Spanish population [J]. J Forensic Sci, 2006,51(5):990-995.
[18] KUSEC V, SIMIC D, CHAVENTRE A, et al. Age, sex and bone measures of the second, third and fourth metacarpal (Island of Pag, SR Croatia, Yugoslavia) [J]. Coll Antropol, 1988,12(2):309-322.
[19] SARRINGHAUS L A, STOCK J T, MARCHANT L F, et al. Bilateral asymmetry in the limb bones of the Chimpanzee (Pan troglodytes) [J]. Am J Phys Anthropol, 2005,128(4):840-845.
[20] CASE D T, ROSS A H. Sex determination from hand and foot bone lengths [J]. J Forensic Sci, 2007,52(2):264-269.
[21] KRISHAN K, KANCHAN T, SHARMA A. Sex determination from hand and foot dimensions in a North Indian population[J]. J Forensic Sci, 2011,56(2):453-459.
[22] ESHAK G A, AHMED H M, ABDEL GAWAD E A M. Gender determination from hand bones length and volume using multidetector computed tomography: A study in Egyptian people[J]. J Forensic Leg Med, 2011,18(6):246-252.
[23] 赵晓进,王训练,党晓云, 等. 太行山猕猴掌骨和蹠骨长度比率的性别差别[J]. 解剖学报, 2009,40(6):993-996.
[24] 赵晓进,赵俊杰,王 耿,等. 太行山猕猴掌骨和蹠骨长度变量性别判别分析[J]. 人类学学报, 2009,28(1):88-94.
(编辑 王 健)
