甘肃白龙江干旱河谷主要灌木种群生态位研究
2015-07-12陈国鹏黄旭东
王 飞,郭 星,陈国鹏,黄旭东
(甘肃省白龙江林业科学研究所,甘肃 兰州 730070)
甘肃白龙江干旱河谷主要灌木种群生态位研究
王 飞,郭 星,陈国鹏,黄旭东
(甘肃省白龙江林业科学研究所,甘肃 兰州 730070)
采用2种生态位宽度指数、生态位重叠指数以及生态位相似比例指数公式分别计算并分析了甘肃白龙江干旱河谷主要灌木18个种群的3种指数。2种生态位宽度计算公式的测度结果表明:除小叶香茶菜、河朔荛花、多花胡枝子、小叶石积木、川甘亚菊和荆条的生态位宽度较大外,大多数种群的生态位宽度较小,反映其群落结构的单调性。具有较高生态位宽度的物种一般具有较高的生态位重叠,但并非生态位宽度小的物种其重叠值也就小,其生态位重叠值的大小与生态位宽度不呈明显相关性。利用生态位相似性比例结果表明:小于0.1的种对有117对,占总种对的76.5%,说明种群对环境的需求有较大的差异。灌木种群生态位的研究可用于指导白龙江干旱河谷植被的保护和恢复。
灌木种群;生态位宽度;生态位重叠;生态位相似性比例;甘肃省白龙江干旱河谷
生态位研究是近代理论生态学的一个主要内容,生态位宽度和重叠的测定是一个非常复杂的问题[1],是生态学中的一个重要概念,是种群生态研究的核心问题[2]。生态位是指在自然生态系统中一个种群在时间、空间上的位置及其与相关种群之间的功能关系[3],生态位理论已被广泛地用于研究植物种间关系、群落结构、群落演替、生物多样性、物种进化及濒危物种评价等方面[4~7],成为解释自然群落中物种共存与竞争机制的基本理论。生态位宽度表示种群生长过程中综合利用资源的能力、利用资源多样化的程度和竞争水平。生态位宽度和种间生态位重叠被认为是物种多样性及群落结构的决定因素,反映该种群对资源的利用能力及其在群落或生态系统中的功能和位置,也反映了其所在群落的稳定性[8~9]。
干旱河谷是我国西南地区山地的特殊类型,在同区域山地垂直带中,干旱河谷带是相对脆弱的地带,是长江上游山地生态环境最脆弱,也是在山区整治中最关键和最困难的一种特殊地域类型[10]。在计算各个种群的生态位宽度和种群间的生态位重叠时,绝大多数研究都以群落调查中的每个取样地为“资源状态”,以各个种群在不同取样地中的个体数目[1]、重要值[11]等为种群的数量特征。灌丛是白龙江干旱河谷地区稳定的植被类型,在植被恢复中具有重要的地位和作用,对地区主要物种种群生态位的研究还尚未见报道。种群的资源利用能力是种群分布与群落演替的内在动力,因此通过对干旱河谷主要灌木种群生态位的研究,能够明确不同灌木种类对环境的生态适应特点,了解各种群在群落中的地位和作用以及种群间的相互关系,对植被资源保护、可持续利用和植被的恢复重建等具有重要意义[12]。
1 研究地区概况
白龙江发源于甘、青、川三省交界处的郎木寺附近,南以岷山与四川分界,西接积石山高原,北以迭山与黄河水系一级支流洮河分水,向东南流至四川昭化汇入嘉陵江,全长约600 km,为长江水系北部的主要二级支流。流域整体位于青藏高原东北边缘,白龙江流经的地域广、落差大,河道全长576 km,其位于东径102°46′~104°52′,北纬33°04′~35°09′,属典型的西北高山地形,温度、降水具有明显的垂直梯度和水平差异[13]。由于坡陡谷深、降水量偏少、峡谷地貌对太阳辐射的反复折射和地面强烈辐射造成的增温作用及焚风效应,加之樵采、垦荒、放牧、筑路等人为干扰的强度和频度相对较大,在沿江海拔2100 m(迭部县尼傲乡)以下至760 m(文县口头坝乡)和主要支流(阿夏沟、岷江、拱坝河、北峪河、羊汤河、白水江)两岸的河谷及浅山地带,形成以气候干暖少雨、植被稀疏残败、地形破碎、土地石漠化及岩漠化为主要特征的干旱河谷景观,泛称为干热河谷。其年均温一般为13~16.5℃,局部河段为6.7℃或20℃,最暖月平均气温17~23℃,最冷月平均气温-2.5~1℃,无霜期148~280 d,年平均降水量300~500 mm,年平均蒸发量1 740~2 122 mm,年平均蒸发量是年平均降水量的3~5倍,全年干燥度1~4。植被主要由旱中生小叶落叶具刺灌木及耐旱草本(禾草为主)植物组成.根据气候要素并参考植被特征,白龙江干旱河谷主要为干暖河谷类型,局部为干温河谷类型[14~15]。
2 研究方法
2.1 样方设计
取样于白龙江干旱河谷地区样地海拔793~2 106 m之间,共计73个样方,每个样方地点的选择兼顾不同的群落类型,所选取的样方具有代表性,能反映白龙江干早河谷植被的主要类型,生境条件等。
植被调查:
(1)群落灌木层设置 5 m× 5 m样方,记录样方内植物名称、经纬度、海拔高度、坡度、坡向等因子,测量每一物种高度、基径、株丛数、多度、盖度。
(2) 草本层取样,在灌木样方四角和中央各取一个1 m×1 m草本样方,记录样方内植物名称,测量每一物种高度、基径、株丛数、多度、盖度等因子。
2.2 数据分析
2.2.1 重要值(IV) 采用IV=RA(相对多度)+RF(相对频度)+RP(相对优势度)计算种群的重要值[16]。其中乔木树种以胸高断面积表示相对优势度,灌木、草本和层间植物以相对盖度表示相对优势度。

式中:B(sw)i是种i在r个资源状态下的生态位宽度,Pij是种i对资源位j的利用与该种对全部资源位的利用之比值,nij是种i在资源位j上的相对优势度,Ni是种i在r个资源位上的相对优势度之和,其标准化形式为:B'(sw)i=1/lgs×B(sw)i,其中s为种群数, 以Simpson指数的倒数来测定种的生态位宽度。其计算公式为:

其中:Pij和r的含义同上,其标准化形式为:B'(L)i=(B(L)i-1) /(r-1)。
2.2.3 生态位重叠的计测公式[20]为:

式中,Lih为种物i重叠物种h的生态重叠指数,Lhi为种物h重叠物种i的生态重叠指数。
2.2.4 生态位相似比例计测公式[21]:
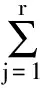
式中:Cih为种i与种h之间的生态位重叠值,即种i与种h利用资源的相似性程度,域值也为[0,1];Phj=nhj/Nh。
3 结果与分析
3.1 重要值
重要值是一个重要的群落定量指标,并常用于比较不同群落间某一物种在群落中的重要性[22],据甘肃白龙江干旱河谷的94种74个样地调查资料,重要值排名前18位的种群重要值占总重要值的70.87% (表1),本文以这18个种群作为甘肃白龙江干旱河谷主要种群进行生态位宽度、重叠和相似性的研究。

表1 甘肃白龙江干旱河谷主要种群的重要值
3.2 主要灌木种群生态位宽度
组成灌丛的种群生态位是组成灌丛群落各种群适应所处环境和不断利用各种环境资源(水分、养分等)的结果。生态位大小反映了种群对环境适应能力或对资源的利用程度。生态位宽度越大,物种对生态因子适应幅度就越大,在群落内的分布幅度也就较大[23]。各种群的生态位宽度见表2。
对白龙江干旱河谷主要种群2种生态位宽度计算公式的测度结果及标准化结果基本一致,表明大多数种群的生态位宽度较小、生态幅度较窄。采用Levins公式中的Shannon-W iener指数测定种的生态位宽度生态位宽度最大的5个种小叶香茶菜、河朔荛花、多花胡枝子、小叶石积木和川甘亚菊,以Simpson指数的倒数来测定种的生态位宽度最大的5个种小叶香茶菜、河朔荛花、多花胡枝子、荆条和川甘亚菊,样方调查统计资料也表明这6个种群的个体数量和样方频度要明显大于其他种群,在群落各个层次中都占较大优势,反映出它们具有较宽的资源利用谱,对资源的利用处于领先地位,在创造和维持干旱河谷植物群落内部环境中起着重要作用,对环境变化的生态适应性较强,其数量多,分布广泛且较为均匀,对环境资源的利用最为充分,竞争能力最强。这与实际调查的情况基本相吻合。以这些物种为优势种群的群落在干旱河谷是较为稳定的生态系统类型。

表2 白龙江干旱河谷主要种群的生态位宽度值
生态位宽度较小的物种,对资源的利用能力较弱,竞争能力也较弱。2种生态位宽度计算公式的测度基本一致。采用Levins公式中的Shannon-W iener指数测定种的生态位宽度生态位宽度最小的5个种刺旋花、少齿小檗、虎榛子、细枝栒子和马鞍羊蹄甲,以Simpson指数的倒数来测定种的生态位宽度最大的5个种刺旋花、杭子梢、铁杆蒿、虎榛子和马鞍羊蹄甲,它们分布范围较小,这些物种的种群生态位宽度较小的原因可能是逐渐变化的环境中,适合它们分布的资源范围变窄,所适应的某一资源在生境中分布较窄,或者说在变化的生境中遇到对其生长或分布起限制作用的资源维较多。一个种的生态位越宽,该物种的特化程度就越小,也就是说它更倾向于一个泛化种;相反,一个种的生态位越窄,该种的特化程度就越强,即它更倾向于一个特化种[24]。
3.3 主要灌木种群生态位重叠分析
在群落中,复杂的生态关系使各种群的生态位通常不是表现为离散的,而总是倾向于分享其它种群的生态位部分,结果导致2个或更多的植物种群对某些资源的共同需求,使不同种群的生态位之间常处于不同程度的重叠状态。白龙江干旱河谷优势种群生态位重叠值见表3。在306对重叠值中,重叠值大于0.1的有33对占10.8%,小于0.1大于0.05的有28对占9.2%,小于0.05大于0.01的有113对占36.9%小于0.01的有132对占43.1%,生态位宽度大的种群间生态位重叠值Lhi与Lih较大,说明生态位宽度大的种群间生态位重叠值也大;生态位宽度小的种与生态位宽度大的种的Lhi较小但Lih较大,如小叶香茶菜与刺旋花和川甘亚菊与马鞍羊蹄甲,它们的Lhi值分别是0.167、0.095而Lih值分别是0.008、0.026,说明生态位宽度大的种与生态位宽度小的种的重叠值大,而生态位宽度小的种与生态位宽度大的种的重叠值较低,这与赵发珠[24]等人对陕北典型退耕还林区林下草本物种生态位研究相一致。
另外,本研究中生态位重叠最高的种对出现在小叶香茶菜与杭子梢之间,其Lhi值为0.845, Lih值为0.067。说明这两种植物的资源利用方式非常相似,两者存在一定的资源竞争;而生态位重叠最低的出现在胡枝子与细枝栒子之间,其Lhi值为 0.001,Lih值为0.001,反映了两物种生物生态学特性的异质性。在所有重叠值中,有54个种对的生态位重叠值为0, 占所有种对的17.6%,这一结果体现了这些种群在不同退耕模式及年限群落中, 由于种群间长期的竞争和适应, 生态位明显趋于特化。

表3 白龙江干旱河谷主要种群的生态位重叠值
注:接上表,种编号同表2。
3.4 主要灌木种群生态位相似性比例值分析
表4列出了白龙江主要灌木种群生态位相似性比例值,其中相似性比例大于0.1的有36对,约占群总对的23.5%,说明这些种对资源的利用相似程度较大,小于0.1的有117对,约占群总对的76.5%,说明这些种对资源的利用相似程度较小,生态位宽度大的物种其相似性比值也表较大,如小叶香茶菜与多花胡枝子、河朔荛花与多花胡枝子和小叶香茶菜与少齿小檗相似性比例值分别为0.391、0.336和0.309;而小叶香茶菜与酸枣和川甘亚菊与荆条生态宽度比较大,但相似性比例值比较小,说明物种生态位宽度与物种间的相似性比例值并不一致。
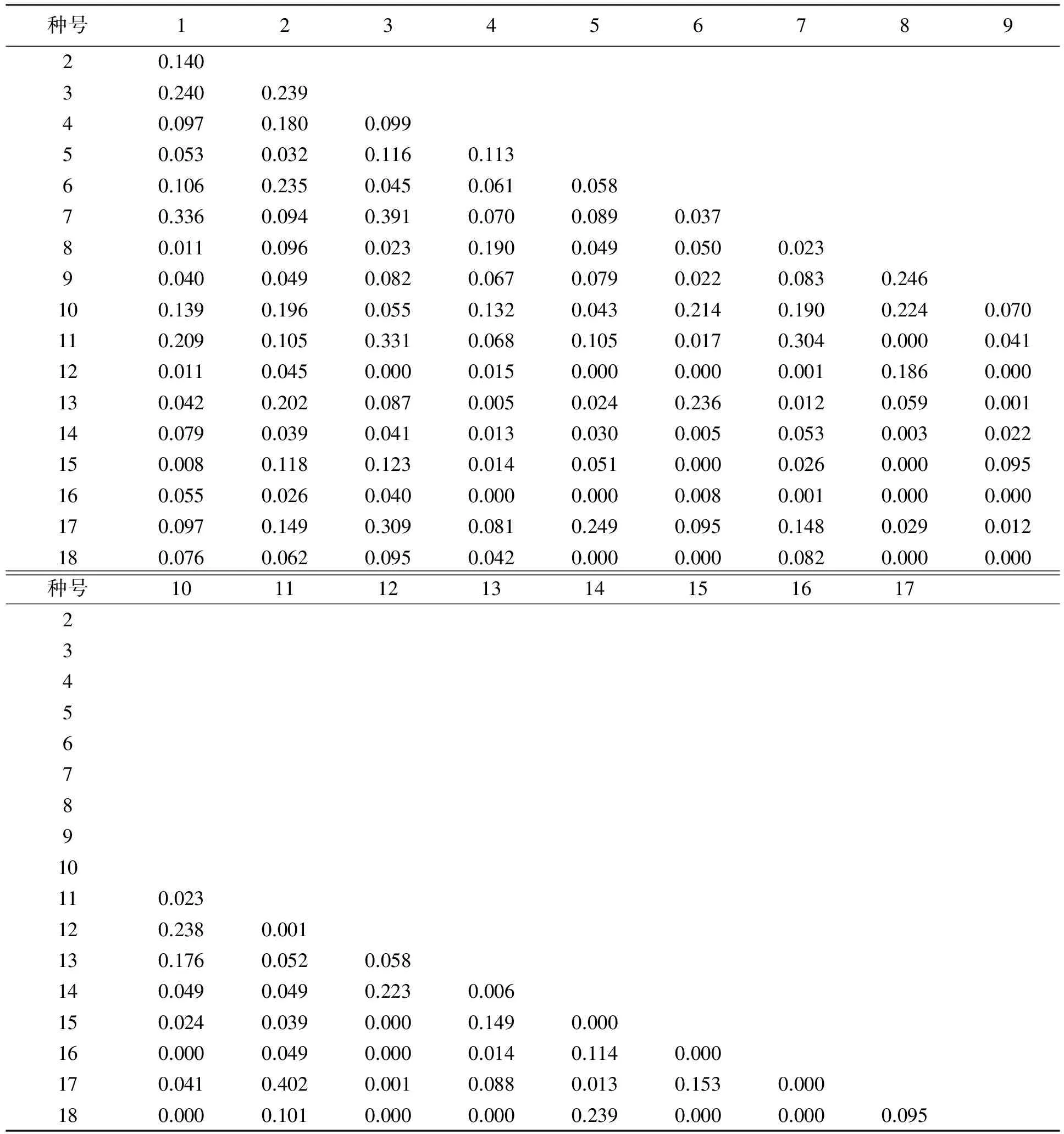
表4 白龙江干旱河谷主要种群的生态位比例相似值
4 结论与讨论
(1)生态位宽度值能很好地反映种群的生态适应性和利用资源的能力。白龙江干旱河谷主要灌木种群的生态位宽度的大小表明了它们在群落中的地位与作用,生态位宽度较大的6种为小叶香茶菜、河朔荛花、多花胡枝子、小叶石积木和川甘亚菊、荆条,这些具有较宽生态位的种群,其重要值也相应的较大,说明具有较强的生态适应性,对环境条件的要求不高,因而生存机会多,在白龙江干旱河谷地区有着较为广泛的分布,形成了该地区的优势种群。其所在的群落一般也是较为稳定的生态系统类型。大多数种群生态位宽度较小,反映白龙江干旱河谷群落结构的单调性;而重要值较小的种群其生态位宽度也比较小,由于对生存环境的要求比较高,只分布在一定的范围内,需加强对这些种物的保护。
(2)具有较大生态位宽度物种的种群往往具有较高的生态位重叠,但并非种群生态位宽度小的其重叠值也小,原因可能是具有较低生态位的物种本身的生态学特性不一定相同,对环境的适应也有较大的差别,对资源段位的要求并不完全一致,从而导致它们之间的生态位 重叠不一定较低。另一方面,环境条件也是影响此结果的重要原因 ,一定海拔范围内,影响此结果的关键因子是坡向、坡度存在的差异。它代表着光照、水分、养分等因素的变化,直接影响着水肥的再分配,使得物种所处的环境资源不同,但是, 生态位重叠值高也并不一定竞争激烈,若可利用的资源能满足物种的需求,一定的生态位重叠往往意味着对资源的有效利用[26]。
(3)本研究结果表明,生态位宽度值较大的两个种,其相似性比值可能高也可能低,生态位宽度值小的两个种其相似性比值也可能高或低。这一结论与赵发珠[24]对陕北典型退耕还林区林下草本物种生态位研究结果基本一致。究其原因可能是生态位宽度较大的物种之间,其资源的利用广泛,对资源的利用程度高。
灌木种群生态位的研究可用于指导白龙江干旱河谷植被的恢复。植被恢复的过程就是群落的建立和演替过程,其核心是物种的更替[27]。白龙江干旱河谷的植被恢复,应首先恢复适应性强的乡土物种,如小叶香茶菜、河朔荛花、多花胡枝子、荆条等作为首选物种,建立以这些物种为优势种,其他物种作为辅助物种的生态系统,达到生态系统回复的目的。
[1] 余世孝.鼎湖山厚壳桂群落优势种生态位宽度与重叠之研究[J].热带亚热带森林生态系统研究, 1985,(3):32-41.
[2] 柳江,洪伟,吴承祯,等.退化红壤区植被恢复过程中灌木层主要种群的生态位特征[J].植物资源与环境学报,2002,11(2):11-16.
[3] 孙儒泳,李庆芬,牛翠娟,等.基础生态学[M].北京:高等教育出版社,2002.
[4] 陈存及,陈新芳,刘金福,等.人工——天然杉阔混交林种群生态位及竞争研究[J].林业科学,2004,40(1):78-83.
[5] 魏文超,何友均,邹大林,等.澜沧江上游森林珍稀草本植物生态位研究[J].北京林业大学学报,2004,26(4):7-12.
[6] 颜廷芬,丛沛桐,刘兴华,等.环境因子对植物生态位宽度影响程度分析[J].东北林业大学学报,1999,27(1):35-38.
[7] ROSENTHAL G.Selecting target species to evaluate the success ofwet grassland restoration[J].Agriculture Ecosystem&Environment,2003, 98(3):227-246.
[8] 吴明作,刘玉萃,杨玉珍,等.河南省栓皮栎林主要种群的生态位研究[J].西北植物学报,1999,19(3):511-518.
[9] 李军玲,张金屯,郭逍宇,等.关帝山亚高山灌丛草甸群落优势种群的生态位研究[J].西北植物学报,2003,23(12):2 081-2 088.
[10] 晏兆莉,陈克明.岷江干旱河谷的生态特征与植被恢复研究[J].世界科技研究与发展, 2000,22(增刊):36-38.
[11] 吴承祯,洪伟,蓝斌,等.万木林中亚热带常绿阔叶林主要种群生态位研究[J].江西农业大学学报, 1996, 18(3):292-298.
[12] 冶明生,关文彬,吴斌,等.岷江干旱河谷主要灌木种群生态位研究[J].北京林业大学学报, 2008, 28(1):7-13.
[13] 邱祖青,杨永宏.白龙江干旱河谷木本植物多样性及其区系地理特征[J].甘肃农业大学学报2007,(10):119-125.
[14] 中国科学院青藏高原综合科学考察队.横断山区干旱河谷[M].北京:科学出版社,1992.
[15] 杨兆平,常 禹,布仁仓,等.岷江上游干旱河谷区域空间变化的定量判定[J].生态学报,2007,27(8):3250-3256.
[16] 云南大学生物系.植物生态学[M].北京:人民教育出版社, 1980:192-195.
[17] LEVINS R.Evolution in changing environments: some the-retical explorations [M].New Jersey: PrincetionUnivers-ity Press, 1968:116-121.
[18] PETRAITS P S.Likelihoodmeasures ofniche breadth andoverlap [J].Ecology, 1979, 60(4):703-710.
[19] 王伯荪,李鸣光,彭少麟.植物种群学[M].广州:广东高等教育出版社, 1995:132-143.
[20] 沈泽昊,方精云.基于种群分布格局的两种水青冈生态位比较研究.植物生态学报,2001,25(4):392- 398.
[21] HURLBERT S H.The measurement of niche breadth and overlap [J].Ecology, 1978, 59(1):67-77.
[22] 方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[M].生物多样性,2009,17(6):533-548.
[23] 王刚,赵松岭,张鹏云,等.关于生态位定义的探讨及生态位重叠计测公式改进的研究[J].生态学报,1984,4(2):119-127.
[24] 张金屯.数量生态学[M].北京:科学出版社,2004:110-114.
[25] 赵发珠,韩新辉,杨改河,等.陕北典型退耕还林区林下草本物种生态位研究[J].西北植物学报,2011,31(10):2090-2099.
[26] 林思祖,黄世国,洪伟,等.杉阔混交株主要种群多维生态位特征.生态学报,2002,22(6):962- 968.
[27] 张继义,赵哈林,张铜会,等.科尔沁沙地植物群落恢复演替系列种群生态位动态特征[J].生态学报,2003,23(12):2741-2746.
The Niche of Shrub Populations Located in the Dry River Valley of Bailong River in Gansu
WANG-Fei GUO-Xing CHEN-Guo peng HUANG-Xu dong
(BailingjiangForestInstitute,Lanzhou,Gansu730070)
Niche breadth index, niche overlap index and niche similarity index were applied to calculate the niche index of 18 shrub populations located in the dry rive valley of Bailong River.Two niche breadth indexes showed that apart from Isodon parvifolius, Wikstroemia chamaedaphne, Lespedeza floribunda, Osteomeles schwerinae, Ajania potaninii and Vervain Family which have wide niches, most of shrub populations have narrow niches, reflecting the monopoly of population structure.Species with wider niches normally would have higher niche overlaps, which, however, does not mean that populations with narrow niche would have lower overlaps of niches.There is no significant correlations between niche breadth and niche overlap.Niche similarity index showed that there are 117 species pairs with similarity index lower than 0.1, accounting for 76.5%, indicating the great difference of demand for environment by species populations.
Shrub populations; niche breadth; niche overlap; niche similarity; dry river valley of Bailong River
2015-02-18 基金项目:甘肃省科技支撑计划项目,项目编号:090NKCK119。
王飞(1986-),男,本科,助理工程师,主要从事林业科学研究。E-mail:173763825@qq.com。
S731.2
A
1001-2117(2015)04-0001-07
