外源NO对水分胁迫下平邑甜茶幼苗氧化损伤的缓解效应
2015-07-04王东方王汉海邹岩梅束怀瑞
曹 慧,王东方,王汉海,邹岩梅,束怀瑞*
(1 潍坊学院 山东高校生物化学与分子生物学重点实验室,山东潍坊261061;2 国家苹果工程技术研究中心,山东泰安271018)
一氧化氮(nitric oxide,NO)是生物体内通过酶促和非酶促途径产生的一种生物活性分子[1-2],广泛存在于植物组织中[3]。NO 也可作为重要的信号分子,它能使非生物胁迫条件下的植物生长发育免受活性氧(reactive oxygen species,ROS)的伤害,且其效应与植物细胞的生理条件及NO 处理浓度有关[4-5]。有研究表明,外源NO 通过增强盐胁迫下小麦[6]、黄瓜[7]、辣椒[8]幼苗,及 高 温、镉胁迫下姜[9]、水稻幼苗[10]超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性,提高非酶促抗氧化物质谷胱甘肽(GSH)和抗坏血酸(ASA)等的含量,从而缓解由于胁迫导致的叶片ROS的累积。全球每年因干旱导致作物减产约高达20%,如何提高作物的抗旱性是植物抗逆研究的重要课题,因此,研究水分胁迫下NO 与植物抗旱性的关系具有重要意义。平邑甜茶为北方苹果树的一种常用实生砧木,具有无融合生殖的特点。关于NO 在植物抗逆过程中的作用,目前研究主要集中在农作物、经济作物等植物上[11-13],而对木本植物的研究鲜有报道,尤其对苹果属植物的研究更为少见。因此,本研究选取苹果属植物平邑甜茶幼苗为试材,通过水培方法添加不同浓度的一氧化氮供体硝普纳(SNP)探讨渗透胁迫下(20%PEG-6000)外源NO对氧化损伤的缓解效应,为进一步了解NO提高植物抗逆性的作用提供理论依据。
1 材料和方法
1.1 材料培养与处理
选用苹果砧木平邑甜茶[Malushupehensis(Pamp.)Rehd.]为试验材料,试验于2005年2月~2008年12月在潍坊学院省级重点实验室进行。平邑甜茶种子经4℃层积处理,蛭石培养。待幼苗长出4~5片真叶以后,选取生长良好一致的幼苗转至1/2Hoagland营养液中培养。培养1周后转为全Hoagland营养液中培养,每周定期更换1 次营养液。当幼苗长到15~16片叶时,选择长势相对一致的幼苗进行试验处理。NO 供体硝普钠(sodium nitrop russide,SNP)购买自Sigma公司,先用蒸馏水配制为100mmol·L-1的母液,4℃保存,使用时按试验所需浓度进行稀释。
采用20%PEG-6000处理相当于-0.63 MPa,轻度水分胁迫模拟干旱处理[14-15]。试验设置6个处理分别为:Hoagland营养液(对照,CK);Hoagland营养液+20%PEG-6000溶液(Ⅰ);Hoagland营养液+20% PEG-6000 溶液+100μmol·L-1SNP(Ⅱ);Hoagland 营养液+20% PEG-6000 溶液+300μmol·L-1SNP(Ⅲ);Hoagland营养液+20%PEG-6000溶液+500μmol·L-1SNP(Ⅳ);Hoagland营养液+20% PEG-6000 溶液+700μmol·L-1SNP(Ⅴ)。每处理8株,3次重复,随机排列,为了保证处理浓度的稳定性,处理期间每天更换1次处理液。于胁迫处理0、3、6、9、12d分别选取中部9~12叶位成熟叶片进行各项生理指标测定,3 次重复。
1.2 测定指标及方法
超氧化物歧化酶(SOD)和过氧化物酶(POD)活性的测定参照陈贻竹等[16]的方法;过氧化氢酶(CAT)活性的测定参照Cakmak等[17]的方法;抗坏血酸过氧化物酶(APX)活性的测定参照Nakano等[18]的方法;抗坏血酸(ASA)含量参照Arakawa等[19]方法测定;产生速率参照王爱国等[20]的方法测定;H2O2含量参照刘俊等[21]的方法测定;丙二醛(MDA)含量参照李合生[22]的方法进行测定。
1.3 数据处理
试验数据采用SAS统计软件进行统计分析,用Duncan’s新复极差法进行多重比较。
2 结果与分析
2.1 外源NO 处理对水分胁迫下平邑甜茶幼苗SOD、POD和CAT活性的影响
由图1还可知,CAT 活性变化趋势与POD 相一致。经20%PEG 胁迫和不同浓度NO 处理,在处理0~3dPOD、CAT 活性变化幅度较为平缓,与对照之间差异未达到显著水平。但处理3d后处理Ⅱ和ⅣPOD、CAT 活性分别在处理后第6天和第9天均升至最高值,分别比对照提高了11.6%、15.4%和3.8%、20.1%,之后呈下降趋势;处理Ⅲ和处理Ⅴ在整个处理期内分别一直呈上升和下降趋势,到第12天与对照之间的差异均达显著水平。
以上结果说明不同浓度NO 处理可不同程度提高SOD、POD 和CAT 酶活性,起到酶促防御保护作用,防止水分胁迫对平邑甜茶所造成的伤害,其防御作用为:处理Ⅲ>处理Ⅳ>处理Ⅱ>处理Ⅴ。
2.2 外源NO 处理对水分胁迫下平邑甜茶幼苗APX活性和ASA含量的影响

图1 外源NO 处理对水分胁迫下平邑甜茶幼苗SOD、POD 和CAT 活性的影响Fig.1 Effects of NO concentrations on SOD,POD and CAT activities in M.hupehensis(Pamp.)Rehd.seedlings under water stress
2.3 外源NO 对水分胁迫下平邑甜茶幼苗活性氧和膜脂过氧化的影响
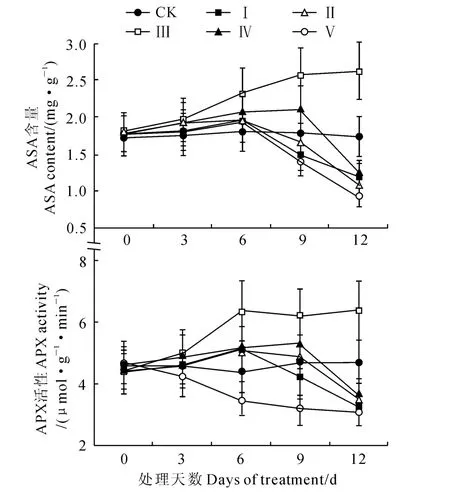
图2 外源NO 处理对水分胁迫下平邑甜茶幼苗ASA 含量和APX 活性的影响Fig.2 Effects of NO concentrations on ASA content and APX activity in M.hupehensis(Pamp.)Rehd.seedlings under water stress
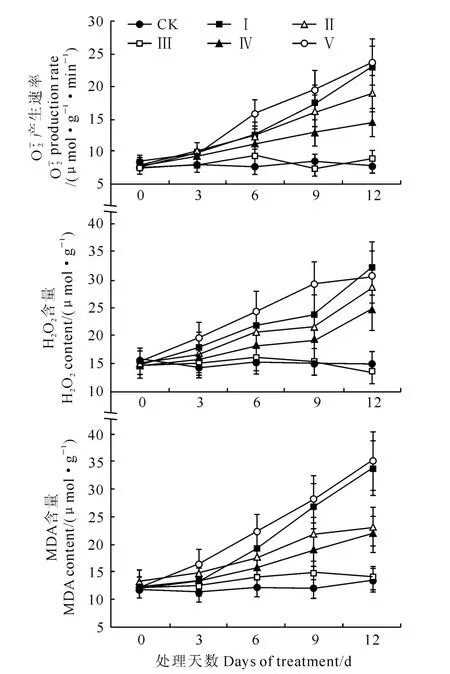
图3 外源NO 处理对水分胁迫下平邑甜茶幼苗产生速率、H2O2 和MDA 含量的影响Fig.3 Effects of NO concentrations on production rate of ,H2O2and MDA contents in M.hupehensis(Pamp.)Rehd.seedlings under water stress
3 讨 论
正常生长条件下,细胞内活性氧的产生量很小(240μmol·s-1)[23],当植物受到胁迫时,体内会产生大量的活性氧,而植物体内也存在活性氧自由基清除系统。SOD 是清除生物体内的唯一酶类,SOD 催化发生歧化反应生成的H2O2则由POD、CAT、APX 和ASA等来清除,APX 与H2O2的亲合力较强,经过抗坏血酸循环分解来完成[9]。本研究表明,在20%PEG 胁迫下,平邑甜茶幼苗叶片SOD、POD、CAT、APX 活性和ASA 含量在胁迫前期均有不同程度的提高,这些酶活性和ASA 含量的提高可能是20% PEG 胁迫前期一种自身适应,是平邑甜茶对逆境胁迫的应激反应,它们协同作用抵抗了胁迫前期诱导的氧化损伤。
随着胁迫时间的延长,至胁迫6d后平邑甜茶幼苗H2O2、和MDA 含量明显增加,均于胁迫后第9天与对照之间的差异达到显著水平。NO 是近年来备受关注的信号分子,它在植物信号传导和抗逆过程中发挥重要作用,且具有浓度效应,低浓度的NO 可作为抗氧化剂对等ROS具有清除作用,并能诱导抗氧化酶基因的表达而具有保护作用[24]。不同浓度的外源SNP 处理(处理Ⅱ~处理Ⅴ)对20% PEG 胁迫下平邑甜茶叶片氧化损伤有不同的效应,300μmol·L-1SNP(处理Ⅲ)处理显著提高了SOD、POD、CAT、APX 活性和ASA 含量,明显降低了产生速率、H2O2和MDA 的含量,与对照之间的差异达到显著水平,从而避免膜脂过氧化,表明300μmol·L-1SNP 能够显著缓解20% PEG 胁迫下膜脂过氧化损伤。外加100 μmol·L-1(处理Ⅱ)、500μmol·L-1(处理Ⅳ)的SNP处理虽然可以在一定程度上缓解干旱胁迫的伤害,但缓解程度与对照之间的差异不显著,且其缓解效果为处理Ⅲ>处理Ⅳ>处理Ⅱ,并随着处理时间的延长缓解作用越来越明显。外源NO 能够缓解氧化胁迫的原因可能是由于NO 作为信号分子对含铁的相关酶类有很高的亲和性,可诱导APX 和POD 等的活性上调或基因表达,也可能直接与反应或通过一系列复杂的信号转导通路提高SOD编码基因的表达而降低水平[4]。
高浓度的外源NO 处理(700μmol·L-1SNP)下,平邑甜茶幼苗SOD、POD、CAT、APX 活性和ASA 含量在胁迫9d 后大幅降低,膜脂过氧化加剧,表明700μmol·L-1SNP以上处理加速了渗透胁迫对平邑甜茶幼苗诱导的氧化损伤。这可能由于一方面过高浓度的SNP本身就是一种胁迫,另一方面高浓度 NO 与相互作用生成大量的ONOO-,而ONOO-经质子化形成具有强氧化性的HOONO,破坏生物大分子的结构和功能[25]。
综上所述,适宜浓度的外源NO 能够通过提高平邑甜茶体内SOD、POD、CAT、APX 活性和ASA含量,有效缓解20% PEG 渗透胁迫对其造成的过氧化伤害,并以300μmol·L-1SNP处理缓解效果最佳,而低于100 μmol·L-1SNP和高于700 μmol·L-1SNP均不利于氧化胁迫的缓解,甚至会造成毒害,这与作者前期的相关研究结果一致[26-27]。
[1]HELGA N,JOSEF M.Indications for the occurrence of nitric oxide synthases in fungi and plants and the involvement in photoconidiation ofNeurosporacrassa[J].PhotochemistryandPhotobiology,1996,64(2):393-398.
[2]BETHKE P C,BADGER M R,JONESA R L.Apoplastic synthesis of nitric oxide by plant tissues[J].ThePlantCell,2004,16(2):332-341.
[3]CHANDOK M R,YTTERBERG A J,VAN WIJK K J,etal.The pathogen-inducible nitric oxide synthase(iNOS)in plants is a variant of the P protein of the glycine decarboxylase complex[J].Cell,2003,113(4):469-482.
[4]BELIGNI M V,LAMATTINA L.Is nitric oxide toxic or protective?[J].TrendsinPlantScience,1999,4(8):299-300.
[5]BELIGNI M V,LAMATTINA L.Nitric oxide protects against cellular damage produced by methylviologen herbicides in potato plants[J].NitricOxide,1999,3(3):199-208.
[6]RUAN H H(阮海华),SHEN W B(沈文飚),LIU K L(刘开力),etal.Effects of exogenous NO donor on glutathione-dependent antioxidative system in wheat seedling leaf under salt stress[J].ActaAgronomicaSinica(作物学报),2005,31(9):1 144-1 149(in Chinese).
[7]FAN H F(樊怀福),GUO SH R(郭世荣),etal.Effects of nitric oxide on the growth and glutathione dependent anti oxidative system in cucumber(CucumissativusL.)seedlings under NaCl stress[J].ActaEcologicaSinica(生 态学报),2008,28(6):2 511-2 517(in Chinese).
[8]YU J H(郁继华),YONG SH Y(雍山玉),ZHANG J B(张洁宝),etal.Protective effects of exogenous nitric oxide on oxidative damage in pepper seedlings under NaCl stress[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2007,27(9):1 801-1 806(in Chinese).
[9]LI X(李 秀),GONG B(巩 彪),XU K(徐 坤).Effects of exogenous nitric oxide on reactive oxygen metabolism in Giger leavers under heat stress[J].ActaHorticulturaeSinica(园艺学报),2014,41(2):277-284(in Chinese).
[10]赵秀峰.一氧化氮对水稻幼苗镉毒害的缓解效应及生理机制[D].南京:南京农业大学,2012.
[11]WANG J(王 建),YU SH X(于世欣),WANG Y J(王逸筠),etal.Effect of exogenous NO on phytochelatins and arginine metabolism in tomato under copper stress[J].SoilandWaterConservation(水土保持学报),2014,28(4):317-323(in Chinese).
[12]YU X J(鱼小军),XU CH L(徐长林),JING Y Y(景媛媛),etal.Effects of exogenous NO germination and seedling growth ofMedicago ruthinicaseeds under NaCl stress[J].GrasslandandTurf(草原与草坪),2014,34(2):68-72(in Chinese).
[13]LIU SH L(刘柿良),PAN Y ZH(潘远智),YANG R J(杨容孑),etal.Effects of exogenous NO on mineral nutrition absorption,lipid peroxidation and ATPase of plasma membrane inCatharanthusroseustissues under cadmium stress[J].JournalofPlantNutritionand Fertilizer(植物营养与肥料学报),2014,20(2):445-458(in Chinese).
[14]CHEN X H(陈新红),WANG ZH Q(王志琴),YANG J CH(杨建昌).Effect of different nitrogen levels and water stress on qualities of rice seedling[J].AgriculturalResearchintheAridAreas(干旱地区农业研究),2007,25(1):78-93(in Chinese).
[15]CAO H(曹 慧),LI CH X(李春霞),WANG X W(王孝威),etal.Research of programmed cell death under water stress inMalusrobustaRehd.andMalushupehensis(Pamp.)Rehd[J].ActaHorticulturaeSinica(园艺学报),2009,36(4):469-474(in Chinese).
[16]CHEN Y ZH(陈贻竹),PATTERSON B D(帕特森).The effect of chilling temperature on the level of superoxide dismutase,catalase and hydrogen peroxide in some plant leaves[J].ActaPhytophysiologicaSinica(植物生理学报),1988,14(4):323-328(in Chinese).
[17]CAKMAK I,MARSCHNER H.Magnesium deficiency and high light intensity enhance activities of superoxide dismutase,ascorbate peroxidase,and glutathione reductase in bean leaves[J].PlantPhysiology,1992,98(4):1 222-1 227.
[18]NAKANO Y,ASADA K.Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplast[J].PlantandCell Physiology,1981,22:867-880.
[19]ARAKAWA N,TSUTSUM I,SANCEDA N G,etal.A rapid and sensitive method for the determination of ascorbic acid using 4,7-diphenyl-1,10-phenanthroline[J].AgriculturalandBiologicalChemistry,1981,45(5):1 289-1 290.
[20]WANG A G(王爱国),LUO G H(罗广华).Quantitative relation between the reaction of hydroxylamine and superoxide anion radicals in plants[J].PlantPhysiologyCommunications(植物生理学通讯),1990,(6):55-57(in Chinese).
[21]LIU J(刘 俊),LÜB(吕 波),XU L L(徐朗莱).An improved method for the determination of hydrogen peroxide in leaves[J].ProgressinBiochemistryandBiologicalPhysicsResearch(生物化学与生物物理进展),2000,27(5):548-550(in Chinese).
[22]李合生.植物生理生化实验原理与技术[M].北京:高等教育出版社,2000.
[23]MITTLER R,VANDERAUWERA S,etal.Reactive oxygen gene net work of plants[J].TrendsinPlantScience,2004,9(10):388-395.
[24]FRANK S,KAMPFER H,PODDA M,etal.Identification of copper/zinc superoxide dismutase as a nitric oxide-regulated gene in human(HaCaT)keratinocytes:implications for keratinocyte proliferation[J].BiochemicalJournal,2000,346(3):719-728.
[25]YAMASAKI H,SAKIHAMA Y,TAKAHASHI S.An alternative pathway for nitric oxide production in plant:new feather of an old enzyme[J].TrendsinPlantScience,1999,4(4):128-129.
[26]CAO H(曹 慧),WANG X W(王孝威),etal.Effects of exogenous nitric oxide on the several enzymes of nitrogen metabolism inMalus hupehensis(Pamp.)Rehd.seedlings under water stress[J].ActaHorticulturaeSinica(园艺学报),2009,36(6):781-786(in Chinese).
[27]CAO H(曹 慧),WANG X W(王孝威),etal.Effects of exogenous nitric oxide on chlorophyll fluorescence parameters and photosynthesis rate inMalushupehensisseedlings under water stress[J].ActaHorticulturaeSinica(园艺学报),2011,38(4):613-620(in Chinese).
