基于RSAP标记的大花三色堇遗传多样性分析
2015-07-04李小梅杜晓华穆金燕刘会超
李小梅,杜晓华,穆金燕,刘会超
(河南科技学院,河南新乡453003)
大花三色堇(Viola×wittrockiana.Gams.),又名蝴蝶花、猫脸花,为堇菜科(Violaceae)堇菜属(ViolaL.)园艺杂交种,由三色堇(V.tricolorL.)、黄堇(V.luteaHuds.)及阿尔泰堇菜(V.alticaKer Gawl.)杂交获得[1]。因其色彩艳丽、花色丰富、花期长、耐寒等优点而成为春秋季重要的花坛与盆栽花卉,在欧洲、美国和日本十分盛行,近年在中国城乡美化中开始大量应用,市场需求快速增长[2]。然而,目前中国大花三色堇生产用种子主要依赖进口,其品种主要为F1代,价高且适应中国生境的品种较少[2]。加快培育具有中国自有知识产权、适应中国生境的大花三色堇品种是当务之急。长期以来大花三色堇育种主要集中在欧美和日本等国的种子公司,因涉及商业秘密,外界对资源间的遗传关系不甚明了。对从各地引进的大花三色堇种质资源进行遗传多样性分析对于大花三色堇种质资源的收集、分类和育种具有重要的指导意义。
在大花三色堇种质资源研究方面,杜晓华等[3]基于表型对来自国内外的33份资源进行遗传差异分析。Ko等[4]将RAPD 标记和表型相结合研究了30种堇菜属植物的亲缘关系。Yockteng等[5]利用ITS与ISSR 标记研究了25种美丽堇菜亚属植物的系统分类。Culley等[6]用ISSR 标记分析了美国俄亥俄州6个堇菜种质亲缘关系,研究城市生境与种质基因的关系。王健等[7]用RAPD 标记研究了18个三色堇自交系的遗传多样性。王涛等[8]采用SRAP标记研究了43份三色堇与角堇资源的遗传多样性。限制性位点扩增多态性(restriction site amplification polymorphism,RSAP)是一种基于基因组上广泛分布的限制性酶切位点多态性的DNA标记系统,具有操作简便、稳定、中等产率等特点[9]。该标 记已在辣椒[10]、紫菜[11]、红花檵木[12]、大白菜[13]、龙须菜[14]、苎麻[15]、重楼属[16]、麦冬[17]等 植物的遗传多样性分析中被广泛应用[18]。但目前尚未见其在堇菜属植物上的应用报道。
本研究随机选取了从国外育种公司和国内育种单位引进后纯化以及本单位选育的41份大花三色堇种质资源,采用RSAP标记技术对其进行遗传多样性分析和亲缘关系分析,旨在为大花三色堇种质资源的收集、分类与种质创新提供依据,并评价RSAP标记系统在大花三色堇遗传多样分析中的适用性及其效力。
1 材料与方法
1.1 材料
供试的41份大花三色堇种质资源为多年自交纯化的自交系(表1),由河南科技学院新乡市草花育种重点实验室提供。
1.2 方 法
1.2.1 RSAP分析 以大花三色堇幼嫩叶片为材料,采用SDS法提取总DNA[19]。
RSAP分析方法参照杜晓华等[10]的方法进行,采用10条RSAP引物(表2),组成45对引物组合,对41份大花三色堇资源进行分析。PCR 扩增引物由生工生物工程(上海)股份有限公司合成。PCR反应体系共25μL,其中模板DNA 为20ng,Mg2+浓度2.5 mmol/L,dNTPs浓度0.2 mmol/L,TaqDNA 聚合酶为1.5 U,2条引物均为600nmol/L。PCR 扩增程序为:94 ℃预变 性5min;94 ℃变性1 min,35 ℃退火1min,72 ℃延伸1min,5个循环;94 ℃变 性1 min,45 ℃退 火1 min,72 ℃延 伸1 min,35个循环;72 ℃延伸10min,4 ℃保存。PCR扩增在Biometra Tgradient PCR 仪上进行。扩增产物用6%变性聚丙烯酰胺凝胶电泳检测,先在60 W 功率下预电泳30min,加样后在75 W 功率下电泳2.5~3h,银染检测,具体参照许绍斌等[20]的方法进行。
1.2.2 数据分析 RSAP分析产生的多态性条带,按二元数据进行统计,有带记为“1”,无帯记为“0”,模糊不清带不统计。每对引物组合的多态性比率(%)=引物组合扩增的多态性条带数/总条带数×100,每个引物的鉴别能力(%)=此引物组合可鉴别的材料数/总材料数×100。采用POPGENE version 1.32 软件计算每对RSAP引物的有效等位基因数、Nei’s遗传多样性指数和Shannon信息指数。采用MVSP 3.1 软件计算各材料间和组群间的Nei’s相似系数和遗传距离,并进行基于UPGMA法的聚类和主坐标作图。

表1 供试的41份大花三色堇种质资源信息Table1 Basic information of 41pansy germplasms used in this study
采用STRUCTURE 2.3.4[21]对所有RSAP标记数据进行Bayesian分析,推测合理组群数目及个体所在的组群。组群数目(K)设定为2~13,假定位点是独立的,采用非混合模型(no admixture),将Length of Burn Period 设为5000,MCMC(Markov Chain Monte Carlo)设定为50000,依据软件计算的后验概率LnP(D)值计算△K值,绘制△K曲线图。按照G.Evanno[22]提出的依据△LnP(D)最大值,推断最合理的组群数目K。
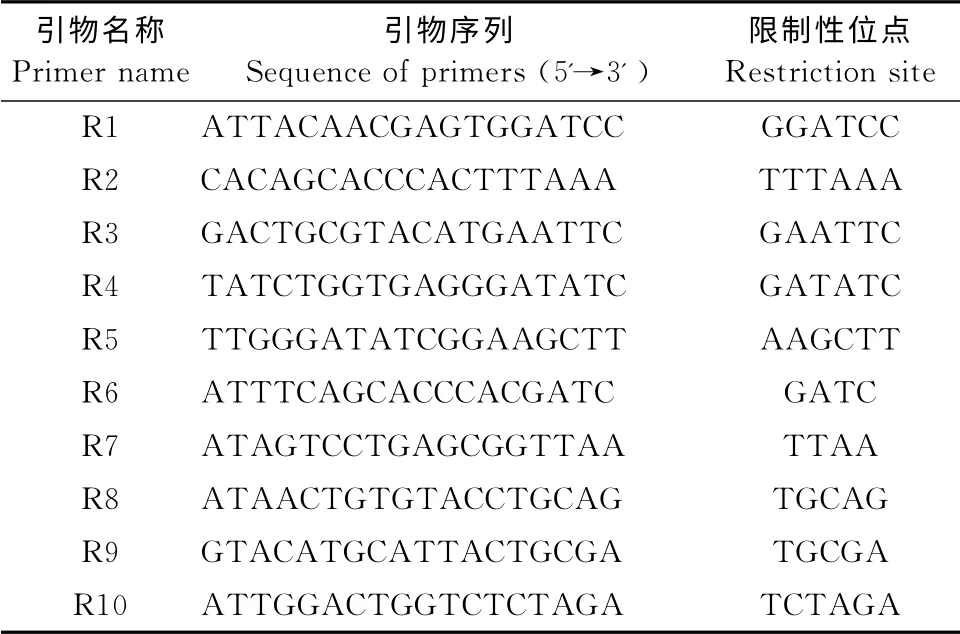
表2 实验用10条RSAP引物Table2 10RSAP primers used in the study
2 结果与分析
2.1 RSAP引物扩增多态性分析
从45 对引物组合中筛选出26 对引物组合对41份材料PCR 扩增结果表明(表3),每对引物组合可扩增出12~37条清晰条带,26对引物组合共扩增出588 条条带;每对引物组合产生多态性条带12~34条,平均单引物组合产生21.81条多态性条带,26对引物组合共产生多态性条带567条,多态性比率97.03%。18对引物组合的多态性条带百分率为100%,对41份大花三色堇资源的鉴别能力为100%,意味着使用其中任何一个单引物组合可区分供试的所有种质资源,占到引物组合总数的69%。其余引物组合的鉴别能力也在83%~98%之间,能区分大部分种质。RSAP 引物有效等位位点数为1.18~1.63,平均为1.40,以R1-R9引物组合所得值最大。
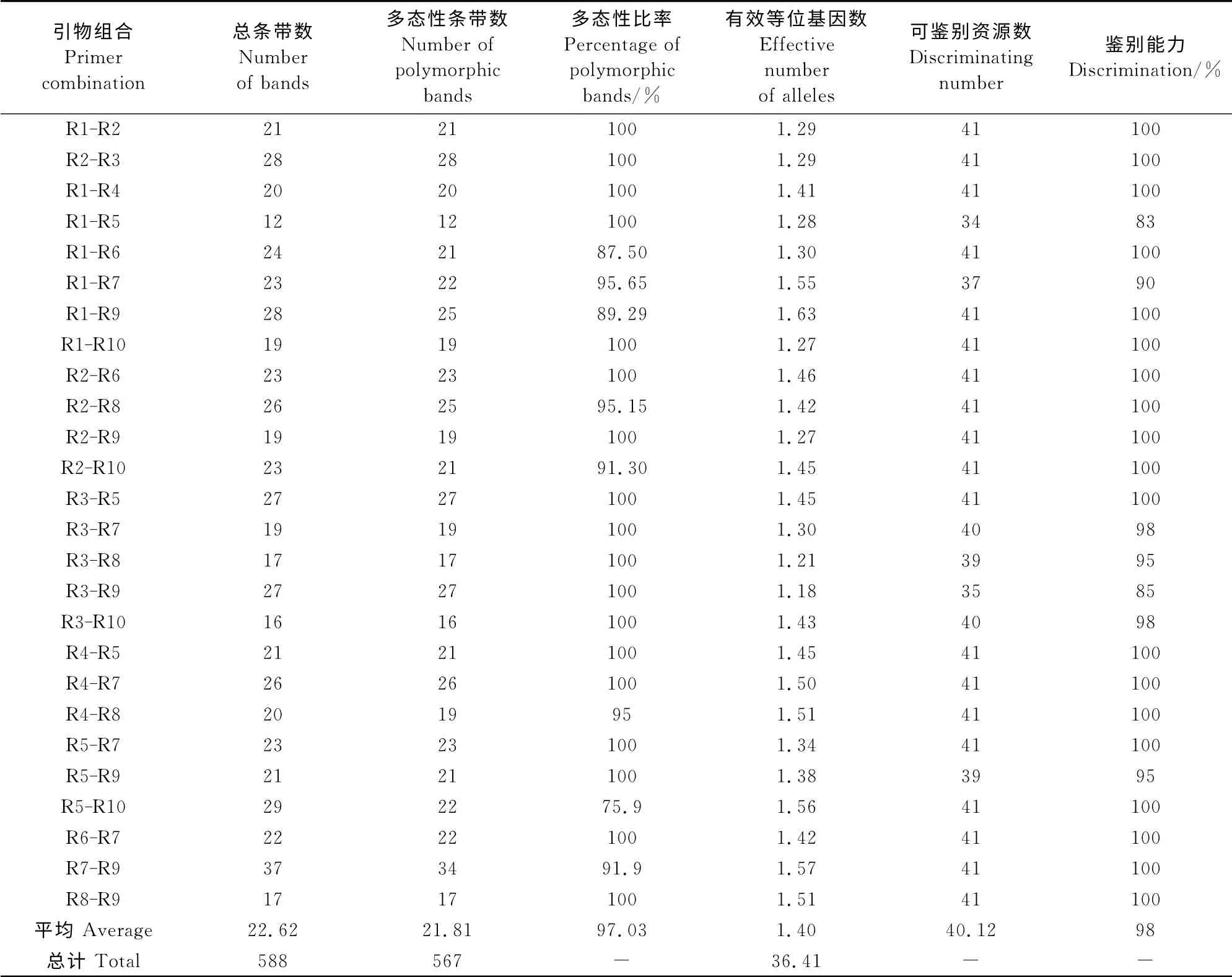
表3 26对引物组合对41份大花三色堇扩增结果Table3 Amplification results of 26RSAP primers on 41pansy accessions
2.2 大花三色堇种质资源遗传多样性与聚类分析
利用POPGENE version 1.32软件计算各位点Nei’s遗传多样性指数和Shannon 信息指数,结果显示,41份大花三色堇种质的平均Nei’s遗传多样性指 数 为0.248 9;平 均Shannon 信 息 指 数 为0.395 1。高于自花授粉植物普通烟草遗传多样性指数0.157[23],与异花授粉玫瑰的遗传多样性指数(0.266 5)和Shannon信息指数(0.403 3)[24]相近。
Bayesian分析结果表明,参试的41份大花三色堇种质最合理组群数为6(表4)。依据“Q 值>0.6视为谱系相对单一”的标准[25],37份种质(90%)可划分到相应的组群中,其中德国Sperli公司和美国泛美种子公司(除M-YC-1外)的所有种质,及荷兰花园种业3份种质(HCG-X-1、HAR2和HAR2-1)和河南科技学院1份种质(YB)归属组群Ⅰ;荷兰花园种业的其余5份种质归属组群Ⅱ;上海园林所所有种质与酒泉金秋园艺和河南科技学院的绝大多数种质归属组群Ⅲ;酒泉金秋园艺1 份种质(229.07)和河南科技学院2份种质(WH 和WO)归属组群Ⅳ;酒泉金秋园艺1份种质(G11-5-1)归属组群Ⅴ;美国泛美1份种质(M-YC-1)归属组群Ⅵ。酒泉金秋园艺3份种质(G11-6-1、229.04和229.05)和河南科技学院的1份种质(YL)在任何组群中的Q 值≤0.6,说明其谱系较为复杂。如果不考虑G11-5-1和M-YC-1,则绝大多数(35 份)种质可归属为4个组群,大多数种质的组群归属与其地理来源存在较高的相关性。

表4 41份大花三色堇种质的遗传结构Table4 Genetic structure of 41pansy germplasms
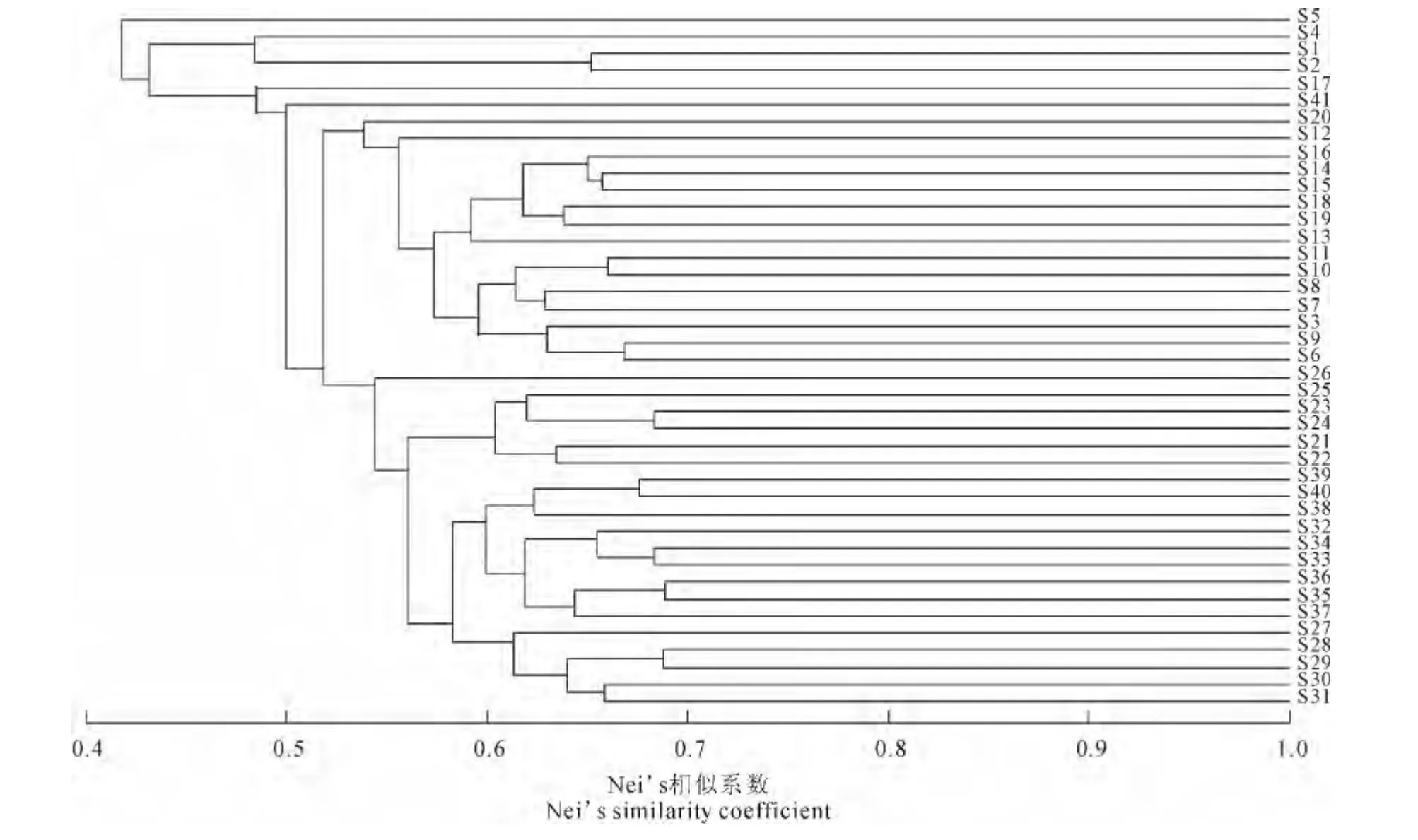
图1 41份大花三色堇的RSAP聚类图Fig.1 Dendrogram of 41pansy accessions based on RSAP
基于Nei’s相似系数的的亲缘关系分析表明,41份材料间的平均遗传相似系数为0.522,其中来自德国Sperli的‘SRFY’与上海园林所的ER01亲缘关系最远,遗传相似系数0.318;上海园林所的EWO 与河南科技学院自选的CW-1之间,酒泉金秋园艺的‘229.01’与‘229.14’之间的亲缘关系最近,遗传相似系数0.688。基于UPGMA 法的聚类结果显示(图1),在相似系数0.430处,41份材料可聚为三大类,其中美国泛美的‘M-YC-1’单独为Ⅰ类;德国Sperli种质和美国泛美的‘XXL-YB’聚为第Ⅱ类;其余37 份种质为第Ⅲ类。第Ⅲ类在相似系数0.518 处,又可分为4 个亚类:荷兰花园种业的‘HMB’单独为A 亚类;河南科技学院YP 为B 亚类;C亚类包括了美国泛美和荷兰花园种业的绝大多数种质资源;D 亚类包括了上海园林所、酒泉金秋园艺及河南科技学院几乎所有材料(除YP外)。聚类结果显示,来源地相近的种质绝大多数聚为一类,表明大花三色堇种质遗传多样性基础与来源地相关性较高。
2.3 不同地理来源大花三色堇种质资源遗传多样性与聚类分析
为进一步分析地理来源对大花三色堇种质遗传多样性影响,综合遗传结构分析结果和聚类结果,将41份材料按照地理来源划分为4个种质群:德国种质群、美国种质群、荷兰种质群和中国种质群。从Shannon信息指数看(表5),从高到低依次为:中国种质群>荷兰种质群>美国种质群>德国种质群。从各地理种质群Nei’s遗传多样性指数看,中国种质群最大(0.238 5),其次为荷兰种质群(0.209 5),再次为美国种质群(0.196 5),德国种质群最小(0.057 8)。
4个地理种群遗传相似系数范围为0.854 4~0.963 6(表5),其中荷兰与中国种质群之间的遗传相似系数最高,为0.963 6,遗传距离最近;而德国和中国种质群的遗传相似系数最低,为0.854 4,遗传距离最远。基于群体间的Nei’s遗传相似系数,采用MVSP 软件作出了地理群体间的三维坐标图(图2),从图2可见,荷兰种质群与中国种质群相距最近,其次为美国种质群,德国种质群相距其他种质群较远。
2.4 不同花色大花三色堇种质资源的遗传多样性
为了解大花三色堇花色的种质资源遗传多样性,将41份材料按花色划分为6个花色群:黄色群、蓝色群、白色群、红色群(包括粉色)、紫色群和黑色群。经Popgene version 1.32软件计算得到大花三色堇6 个花色群Shannon 信息指数变化范围为0.199 4~0.364 9(表6),各花色群多样性指数由高到低依次为:白色群、紫色群、红色群、紫色群、蓝色群、黑色群。总体来看,白色群Shannon信息指数要高于其他花色群,说明白色群的遗传多样性较为丰富;而黑色群和蓝色群的Shannon 信息指数相对偏低,遗传多样性较低。
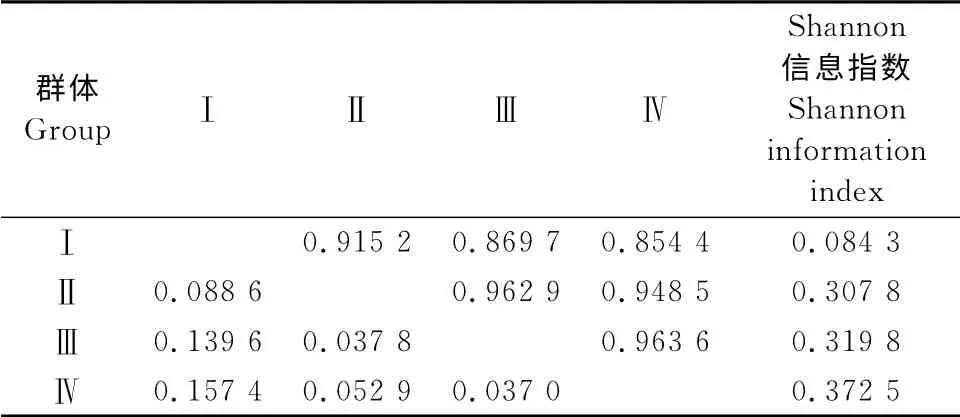
表5 大花三色堇4个地理种质群间的Nei’s遗传相似系数(对角线上方)、遗传距离(对角线下方)和Shannon 信息指数Table5 Nei’s genetic similarity coefficient(above diagonal),genetic distance(below diagonal)and Shannon information index among the four groups of pansy populations separated by geographical distributions
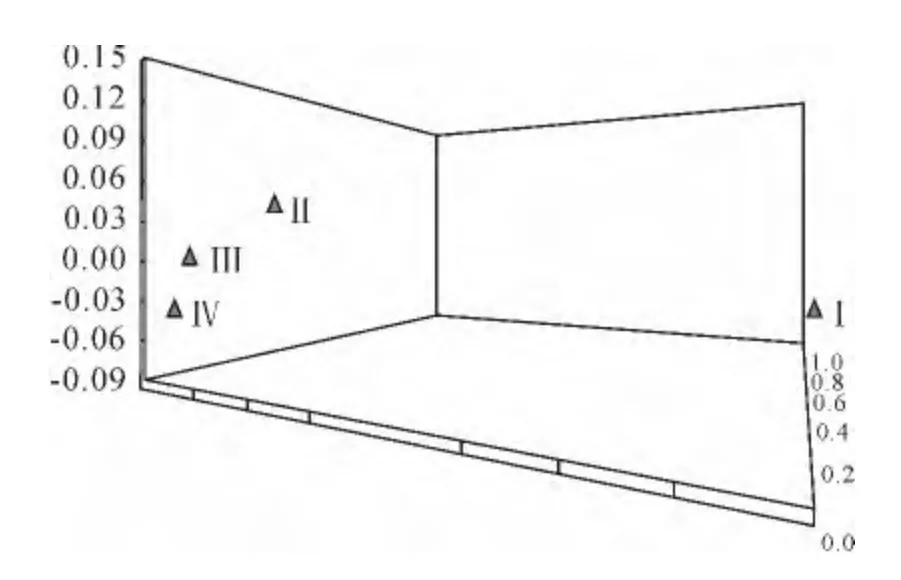
图2 4个大花三色堇地理种质群三维分布图Fig.2 3-dimension distributions of four pansy geographical groups
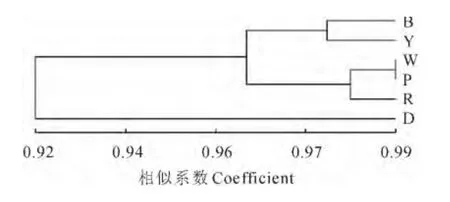
图3 大花三色堇6个花色群的RSAP聚类图Fig.3 RSAP dendrogram of 6pansy groups divided by flower color
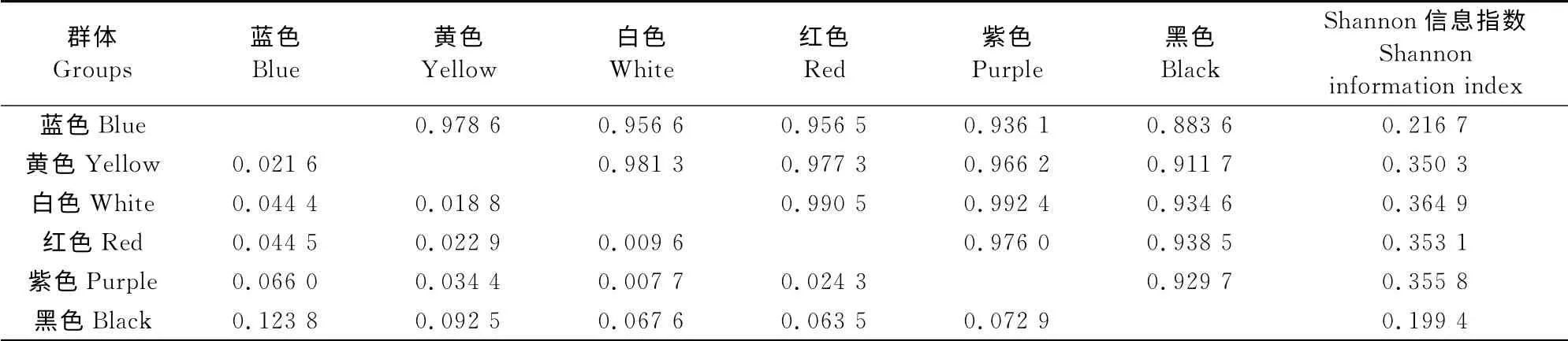
表6 大花三色堇6个花色群体间Nei’s遗传相似系数(对角线上方)、遗传距离(对角线下方)和Shannon信息指数Table6 Nei’s genetic similarity coefficient(above diagonal),genetic distance(below diagonal)and Shannon information index among six pansy groups separated by flower color
6个花色群的遗传相似系数 为0.883 6~0.992 4(表6),其中白色群体与黄色群体、白色群体与紫色群体间的遗传相似系数在99%以上;而蓝色群和黑色群的遗传相似系数最低,为0.883 6。基于各花色群体间Nei’s遗传相似系数聚类结果显示(图3),在遗传相似系数0.964处,6个花色群聚成3类,其中,黑色群体单独为一类,黄色和蓝色群体聚为一类,红色、白色和紫色群体聚为一类。
3 讨 论
3.1 RSAP在种质资源多样性分析中的适用性
操作简便、重复性高、在基因组分布均匀、成本低是理想DNA 标记的重要内容[26]。以揭示基因组限制性位点多态性为目的的RSAP 标记系统与RFLP和AFLP 相比,操作更简便,只需一步PCR就能完成,但具有与AFLP和SRAP(相关序列扩增多态性)相媲美的产率[27]。在三色堇上,本研究单次RSAP-PCR可扩增12~37条带,明显优于RAPD标记[28],与SRAP-PCR 扩增的13~30 条带[8]相近。由于SRAP 仅检测基因开放阅读框区(ORF)多态性[29],而RSAP检测的限制性位点在基因组分布更加广泛,因此对基因组变异的反映会更全面,分析效力可能更高。这一推断在辣椒[27]和三色堇遗传多样性分析中均得到印证。例如,王涛等[8]在对43份三色堇和角堇的SRAP分析中,筛选的21对引物组合中6对鉴别能力达到100%;而本研究采用RSAP 对41 份大花三色堇的分析显示,26对引物组合中就有18对引物组合(69.2%)的鉴别能力达到了100%。从分析成本来看,SRAP 获得了500条多态性条带,合成了引物19条引物;而RSAP合成10 条引物,即获得了567 条多态性条带,应用成本上更低。综上所述,在种质资源多样性分析上RSAP具有一定优势。当然,SRAP 标记的优势在于,发现与目标性状关联的DNA 标记,从而实现基因的定位、克隆或标记辅助选择[26]。
3.2 大花三色堇种质资源多样性与种质交流
从遗传多样性来看,大花三色堇种质的多样性指数高于自花授粉植物普通烟草[23],接近异花授粉植物玫瑰[24],这与大花三色堇的异花授粉习性[28]相一致。同为异花授粉植物,玫瑰栽培历史久,遗传改良和种质创新时间长;三色堇栽培时间相对短[1];但由于三色堇常用做一二年栽培,有性杂交造成的基因重组频繁,可能是其在遗传多样性上与玫瑰接近的主要原因,这也反映出近百年来大花三色堇的育种成就比较突出。
基于Bayesian 的遗传结构分析和基于Nei’s相似系数的聚类结果均表明,来源地相同或相近的大花三色堇种质资源亲缘关系较近,这与杜晓华等[3]基于表型的聚类结果和王健等[7]采用RAPD的研究结论一致,说明大花三色堇的种质交流存在一定的地域限制。原因可能在于:当前大花三色堇育种主要集中在德国、美国和日本等几家种子公司,为了自身利益,延长品种使用寿命,各公司无形中限制了种质交流。来源于国内几个单位的种质亲缘关系较近,反映了国内资源在遗传多样性上仍存在一定的局限性,需要继续加强种质资源的引进和创新。中国种质群与德国种质群的亲缘关系较远,提示应要加强德国种质的引进。
3.3 花色遗传多样性
有关花色研究表明,花色素的形成是一个复杂的生化代谢过程,涉及许多不同基因编码的酶。此外,花色的呈现还受花瓣细胞的结构、细胞液pH 等多种因素的影响[30]。因此不同花色类型的遗传基础不同。本研究通过DNA 标记对三色堇花色遗传多样性分析表明,大花三色堇白色群体的遗传多样性指数最高,说明白色在三色堇中的遗传基础广泛,这可能与三色堇为自然早春开花植物,因授粉媒介的关系白色类型花占主导地位[31]。大花三色堇黑色群体的遗传多样性指数较低,说明黑色类型品种培育在大花三色堇品种选育中受到一定制约。
[1] ZHANG Q SH(张其生),BAO M ZH(包满珠),LU X X(卢兴霞),etal.Research advances in breeding of pansy(Viola×wittrockiana)[J].ChineseBulletinofBotany(植物学报),2010,45(1):128-133(in Chinese).
[2] WANG X Y(王新悦).Breed passionately-visiting ZHU Chaowei,Asian technology consultant of Banner Seed Company in Germany[J].ChinaFlowers& Horticulture(中国花卉园艺),2012,(13):20-22(in Chinese).
[3] DU X H(杜晓华),LIU H CH(刘会超),LIU M G(刘孟刚),etal.Preliminary analysis on genetic difference of 33pansy cultivars[J].JournalofNorthwestForestryUniversity(西北林学院学报),2010,25(4):78-82(in Chinese).
[4] KO M K,YANG J,JIN Y H,etal.Genetic relationship ofViolaspecies evaluated by random amplified polymorphic DNA analysis[J].JournalofHorticulturalScienceandBiotechnology,1998,73(5):601-605.
[5] YOCKTENG R,BALLARD H E,MANSION G,etal.Relationships among pansies(ViolasectionMelanium)investigated using ITS and ISSR markers[J].PlantSystematicsandEvolution,2003,241(3-4):153-170.
[6] CULLEY T M,SBITA S J,WICK A.Population genetic effects of urban habitat fragmentation in the perennial herbViolapubescens(Violaceae)using ISSR makers[J].AnnalsofBotany,2007,100(1):91-100.
[7] WANG J(王 健),BAO M ZH(包满珠).Application of RAPD on analysis of genetic relationship between inbred lines and prediction of heterosis in pansy(Violawrittorckiana)[J].JournalofWuhanBotanicalResearch(武汉植物学研究),2007,25(1):19-23(in Chinese).
[8] WANG T(王 涛),XU J(徐 进),ZHANG X X(张西西),etal.Genetic relationship of 43cultivars ofViolatricolorandViolacornutausing SRAP marker[J].ScientiaAgriculturaSinica(中国农业科学),2012,45(3):496-502(in Chinese).
[9] DU X H(杜晓华),WANG D Y(王得元),GONG ZH H(巩振辉).Development and refinement of a new marker technique-restriction site amplified polymorphism(RSAP)[J].JournalofNorthwestA&FUniversity(Natural Science Edition)(西北农林科技大学学报·自然科学版),2006,34(9):45-54(in Chinese).
[10] DU X H(杜晓华),GONG ZH H(巩振辉),WANG D Y(王得元).Estimation and comparison of genetic distances among elite inbred lines in hot pepper(CapsicumannuumL.)by RSAP and SSR[J].JournalofNorthwestA&FUniversity(Natural Science Edition)(西北农林科技大学学报·自然科学版),2007,35(7):97-102(in Chinese).
[11] QIAO L X(乔利仙),WENG M L(翁曼丽),etal.The application of RSAP marker technique in diversity detection and germplasms identification ofPorphyra[J].PeriodicalofOceanUniversityofChina(中国海洋大学学报),2007,37(6):951-956(in Chinese).
[12] LI X L(李炎林),XIONG X Y(熊兴耀),etal.Establishment and optimization of RSAP-PCR reaction forLoropetalumchinesevar.[J].JournalofHunanAgriculturalUniversity(Natural Sciences Edition)(湖南农业大学学报·自然科学版),2009,35(1):65-68,75(in Chinese).
[13] ZHANG M K(张明科),ZHANG L G(张鲁刚),GONG ZH H(巩振辉),etal.New primers design and reaction system optimization of restriction site amplified polymorphism(RSAP)marker technique[J].JournalofNorthwestA&FUniversity(Natural Science Edition)(西北农林科技大学学报·自然科学版),2009,37(2):148-154,147(in Chinese).
[14] WANG J G(王津果),SUI ZH H(隋正红),ZHOU W(周 伟),etal.RSAP analysis onGracilariopsislemaneiformisand conversion of SCAR marker[J].PeriodicalofOceanUniversityofChina(中国海洋大学学报),2014,44(4):47-53(in Chinese).
[15] ZOU Z ZH(邹自征),CHEN J H(陈建华),LUAN M B(栾明宝),etal.Genetic relationship of ramie germplasm revealed by RSAP,SRAP,and SSR markers[J].ActaAgronomicaSinica(作物学报),2012,38(5):840-847(in Chinese).
[16] XIN B H(辛本华),TIAN M L(田孟良),WU B L(吴镔锣),etal.Genetic diversity analysis of Paris by RSAP markers[J].ChinaJournalofChineseMaterialMedica(中国中药杂志),2011,36(24):3 425-3 427(in Chinese).
[17] XU H CH(徐护朝),ZHANG J Y(张君毅),XU C(徐 灿).Genetic diversity of different populations of lilyturf revealed by RSAP analysis[J].ChinaJournalofChineseMaterialMedica(中国中药杂志),2014,39(20):3 922-3 927(in Chinese).
[18] 中国农业百科全书编辑部.中国农业百科全书(观赏园艺卷)[M].北京:农业出版社,1996:366-367.
[19] 王关林,方宏筠.植物基因工程[M].北京:科学出版社,2009.
[20] XU SH B(许绍斌),TAO Y F(陶玉芬),YANG ZH Q(杨昭庆),etal.A simple and rapid methods used for silver staining and gel preservation[J].Hereditas(遗传),2002,24(3):335-336(in Chinese).
[21] PRITCHARD J K,STEPHENS M,DONNELLY P.Inference of population structure using multilocus genotype data[J].Genetics,2000,155(2):945-959.
[22] EVANNO G,REGNAUT S,GOUDET J.Detecting the number of clusters of individuals using the software STRUCTURE:a simulation study[J].MolecularEcology,2005,14(8):2 611-2 620.
[23] QI J M(祁建民),LIANG J X(梁景霞),etal.Genetic diversity and evolutionary analysis of tobacco(NicotianatabacumL.)germplasm resources based on ISSR and SRAP markers[J].ActaAgronomicaSinica(作物学报),2012,38(8):1 425-1 434(in Chinese).
[24] XU Z D(徐宗大),ZHAO L Y(赵兰勇),ZHANG L(张 玲),etal.Analysis of genetic diversity and construction of fingerprint ofRosa rugosaby SRAP[J].ScientiaAgriculturaSinica(中国农业科学),2011,44(8):1 662-1 669(in Chinese).
[25] XU G B(徐刚标),WU X Q(吴雪琴),JIANG G X(蒋桂雄),etal.Genetic diversity and population structure of an endangered species:TsoongiodendronodorumChun[J].JournalofPlantGeneticResources(植物遗传资源学报),2014,15(2):255-261(in Chinese).
[26] GEORGE A.Principles of Plant Genetics and Breeding(Second Edition)[M].New York:John Wiley &Sons,Ltd.2012:389-423.
[27] DU X H,WANG D Y,GONG ZH H.Comparison of RSAP,SRAP and SSR for genetic analysis in hot pepper[J].IndianJournalof Horticulture,2010,64(4):66-71.
[28] 王健.三色堇杂交育种、RAPD标记辅助育种及组织培养的研究[D].武汉:华中农业大学,2005.
[29] LI G,QUIROS C F.Sequence-related amplified polymorphism(SRAP),a new marker system based on a simple PCR reaction:its application to mapping and gene tagging inBrassica[J].TheoreticalandAppliedGenetics,2001,103:455-461.
[30] 戴思兰.园林植物遗传学[M].北京:中国林业出版社,2010.
[31] FREY F M.Opposing natural selection from herbivores and pathogens may maintain floral-color variation inClaytoniavirginica(Portulacaceae)[J].Evolution,2004,58(11):2 426-2 437.
