基于树干液流技术对杨树冠层吸收臭氧特征的分析1)
2015-06-28陈波鲁绍伟李少宁
陈波 鲁绍伟 李少宁
(北京市农林科学院林业果树研究所,北京,100093)
全球城市化的加速造成了严重的空气污染[1],由于大范围的建设和机动汽车数量的增加,北京市臭氧污染问题非常严重[2]。对流层空气中的臭氧是一种全球范围内的二次污染物,影响植被和人类健康,降低大气可见度[3]。林木是城市生态系统的重要组成部分,在吸收臭氧、改善微气候、净化空气、美化环境等方面发挥着重要作用[4]25。不同林木吸收臭氧速率差别较大,Nowak等[4]26-28对美国14个城市森林冠层去除臭氧速率进行研究,得出其变化范围为 3.82~2.10 g·m-2,北京市森林冠层去除臭氧速率为 5.50 g·m-2[5];Manes 等[6]对不同树种搭配(常绿树种vs落叶树种、阔叶树种vs针叶树种)的树种吸收臭氧速率进行了研究,结果表明差异较大;此外,还有学者对落叶树种与常绿树种的单位冠层投影面积吸收臭氧速率差异的显著程度进行了研究[7-9]。这些研究结果对于改善空气质量具有极大的促进作用。值得注意的是,林木吸收臭氧的研究目前主要集中在林木吸收臭氧速率方面。林木吸收臭氧格局对群落吸收臭氧功能影响显著,因此被认为是林木吸收臭氧的一个复杂、重要方面,同时围绕日变化、季节变化、年际变化格局等方面开展了大量研究[10-11]。
在一定的外界臭氧质量浓度下,气孔导度是整树吸收臭氧的关键限制因子,进而受微气候、冠层位置、树龄及海拔等因素的影响[12-14]。目前,从林木吸收臭氧效率的角度探讨林木吸收臭氧功能和城市林木吸收臭氧的环境影响机制研究较少。现有的大部分研究主要集中在挪威云杉(Picea abies)、瑞士五叶松(Pinus cembra)、欧洲山毛榉(Fagus sylvatica)、欧洲落叶松(Larix decidua)等树种天然林吸收臭氧的影响机制[15-16]。在冠层尺度上城市树木吸收臭氧特征也少有研究,仅见王华等[17]基于树干液流技术,结合天气变化和大气臭氧质量浓度分析,研究夏秋季刺槐(Robinia pseudoacacia)整树冠层吸收臭氧特征及环境影响因素。本研究以北京市杨树人工林为研究对象,采用树干液流技术及环境监测相结合的方法,从速率、效率及环境因子三个方面揭示杨树整树吸收臭氧功能变化的关系,研究结果可为树木吸收臭氧功能、空气质量提高及城市林木管理提供科学依据和理论基础,也对杨树人工林的抚育、生态服务功能的发挥以及生态环境的改善具有重要的理论和实践意义。
1 研究地概况
研究地位于北京市大兴区榆伐镇大兴林场,海拔30 m左右。属暖温带半湿润季风气候。年均温11.5℃,一年中最热月是7月,平均气温26℃,最高气温达40.6℃,最冷月是1月份,平均气温-5.1℃,最低气温可达-27.4℃。≥10的积温4 143℃,年均无霜期204 d,年均降水量568.9 mm,年内的雨量分布极不均匀,7—9月份降水占全年降水的60%~70%,土壤为沙土。光照时间较长,全年光照时间达2 769.3 h,光照率达63%,全年太阳辐射量 561 kJ·cm-2。该区的风向以北风、西北风为主,冬、春两季风多、风速大。人工林内树种主要是107欧美杨(Populus×euramericana“74/76”),其中,75%为 2002年种植,其余为1998年、2001年和2003年种植。混交树种有刺槐(Robinia pseudoacacia)。林分均匀整齐,林下主要为当年生草本植物,主要有紫花苜蓿(Medicago sativa)、黄香草木樨(Melilotus officinalis)、猪毛菜(Salsola collina)、尖头叶藜(Chenopodium acuminatum)、灰菜(Chenopodium album)、蒺藜(Tribulus terrestris)、附地菜(Trigonotis peduncularis)等[18-19]。
2 材料与方法
2.1 杨树标准木特征
在样地选择标准木4株,其特征见表1。
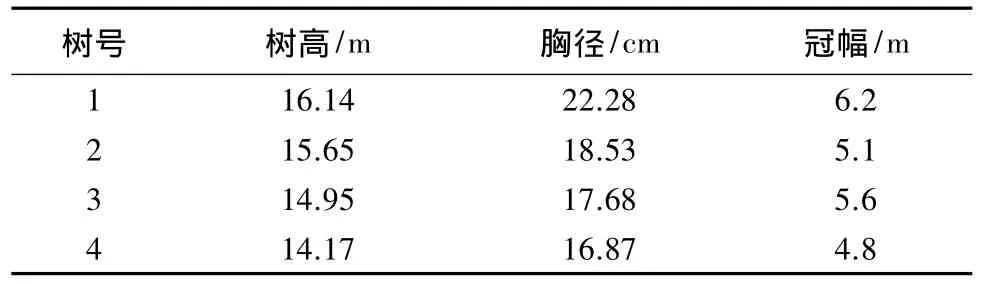
表1 研究区杨树标准木特征
2.2 液流密度的测定
树干边材液流用热扩散式边材液流探针TDP测定。选取待测样木,在树干的东、西两个方位胸高处(1.3 m)钻孔。两孔尽量保持平行,并与树干保持垂直。将TDP探针小心插入钻孔中(含有红色导线的探针在上,2线探针为环境探针在下)。探针与树皮接触处用喷剂式泡沫封住以免雨水接触探针,造成测量不准。为了防止太阳辐射对探头的影响,在安装好探头的外层包上3层铝箔。通过测量这两个固定距离的探针的温度之差(每30 s测读1次,每10 min 进行平均并储存数据)[20],用 Granier[21]经验公式计算树干液流[22]:
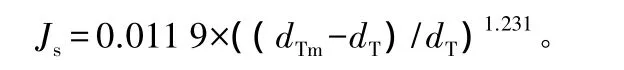
式中:Js为液流密度;dTm为分析日24 h内上下探针的最大温差值;dT为某时刻瞬时温差值,即当时测定的温差值。
2.3 臭氧质量浓度测定
在大兴林场液流监测场附近设有大气臭氧观测站,该站点每小时记录一次臭氧质量浓度数据。
2.4 环境因子测定
实验林内设有美国HOBO自动气象站,用于实时监测空气温度、湿度、风速和风向、光合有效辐射、总辐射、不同深度土壤温度和土壤含水量、降雨量等气象要素参数,所有传感器均与数据采集器相连。设定数据采集间隔为10 min。
2.5 边材面积确定
测定5棵试验树,为避免对研究对象的伤害,选取样地中与研究对象大小最接近的杨树(胸径差别在0.5 cm以内),以生长锥取木芯,直尺测定边材厚度,同时测定其胸径和边材面积,分析二者关系(图1),得到的边材面积作为试验树的边材面积。结果表明,胸径和边材面积之间存在着高度相关的关系,可以用三次曲线较好地表示,其决定系数达到显著水平(R2=0.957 8,p<0.0001)。
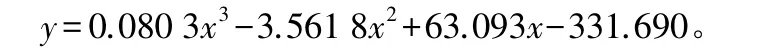
式中:y为边材面积;x为胸径。

图1 胸径与边材面积的关系
2.6 空气水汽压亏缺计算
为了综合反映温度和空气相对湿度对树木蒸腾的协同效应,采用水汽压亏缺这一指标,空气水汽压亏缺(D)是衡量植物蒸腾高低的重要因子,VPD在较低的范围内,气孔导度会随D上升而增大,引起单位叶面积蒸腾(E)逐渐升高至最大值;如果VPD继续上升,气孔导度会呈指数降低,蒸腾出现下降;如果VPD明显增高,气孔会部分关闭[23]。通过以下公式计算求出:
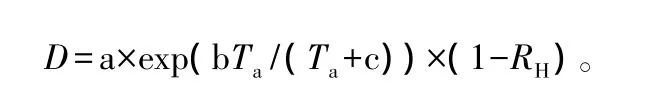
式中:常数 a、b、c 分别为 0.611 kPa、17.502、240.97℃[24];Ta为大气温度;RH为空气湿度。
2.7 冠层蒸腾速率计算
植物冠层在一定时间内单位叶面积蒸腾的水量Ec=Js×(As/AG)[25]。式中:Ec为单棵样树的冠层蒸腾速率;Js为样树液流密度值;As为样树边材面积;AG为样树冠层的地面投影面积。
2.8 冠层导度计算
冠层导度是植物冠层与大气之间的二氧化碳、水蒸气或热量的传导度,计算公式为:
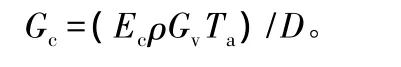
式中:Ec是冠层蒸腾速率;ρ是水的密度;Gv是水蒸气的通用气体常数;Ta是大气温度;Gc整树冠层导度;D是大气水汽压亏缺[26]。
2.9 臭氧吸收速率计算

式中:FO3为冠层臭氧吸收速率;[O3]是外界大气中的臭氧质量浓度;Gc是冠层导度;0.613是转换系数,代表了空气中臭氧和水蒸气扩散系数比[27]。
2.10 数据处理
采用Excel 2003和SPSS 16.0软件,对杨树液流密度、大气水汽压亏缺、冠层导度和臭氧吸收速率进行计算并绘图分析,用曲线拟合杨树边材面积。
3 结果与分析
3.1 液流密度
图2为杨树不同样株5、6、7月份的液流密度月变化(图2A-D)、日变化(图2E-H)以及不同方位的液流密度变化值(图2I-L)。
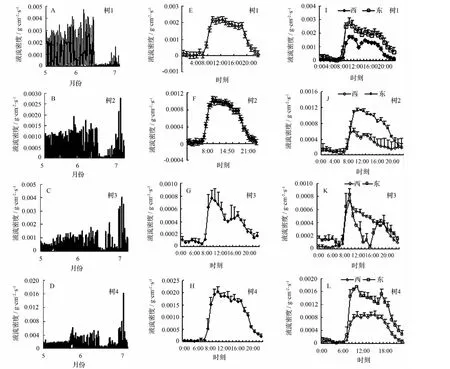
图2 杨树不同样株液流密度月变化、日变化和方位变化
图2中日变化为2013年5月15日的测定值,不同方位变化为2013年5月11日的测定值。从月变化来看,5月最大,6月相对较低,树1变化比较平稳,树3最小,树4的液流密度值最大。从不同方位来看,树3是在10:00以后为西侧液流密度值大于东侧,其它样树均是东侧大于西侧,但总体来看,东侧的液流密度值大于西侧,如图2L,在西侧液流密度最大值仅为 0.000 78 g·cm-2·s-1,而在东侧为0.001 72 g·cm-2·s-1,两者相差了 2.2 倍。说明东侧蒸腾大于西侧,这与东侧接收太阳直射较多有关。
3.2 臭氧质量浓度
大气中的臭氧质量浓度日均值表现出明显的时间变化,从5、6、7三个月份来看,最高质量浓度达到了 342.65 μg·m-3,5 月份臭氧质量浓度最大,7 月份最低(图3)。5月份时最高质量浓度月均值达到了(256.33±55.96)μg·m-3,7 月份为(202.67±36.94)μg·m-3。臭氧质量浓度与温度的高低成正比,温度越高时其对应的臭氧质量浓度值也越高。如图4所示,当臭氧质量浓度为最低的 25.50 μg·m-3时,其对应的温度也为较低值,为21℃,当温度为最低的20.67 ℃时,其对应的臭氧质量浓度仅为 26.00 μg·m-3;当臭氧质量浓度为最大的 175.00 μg·m-3时,其对应的温度为27℃,为一天中的最高温度。由此可知,温度是影响臭氧质量浓度高低的主要因素。因此,在温度过高时为防止臭氧质量浓度过高对树木造成危害,高温时段树木要关闭气孔,减少蒸腾,这或许也是树木对环境的一种响应或适应。
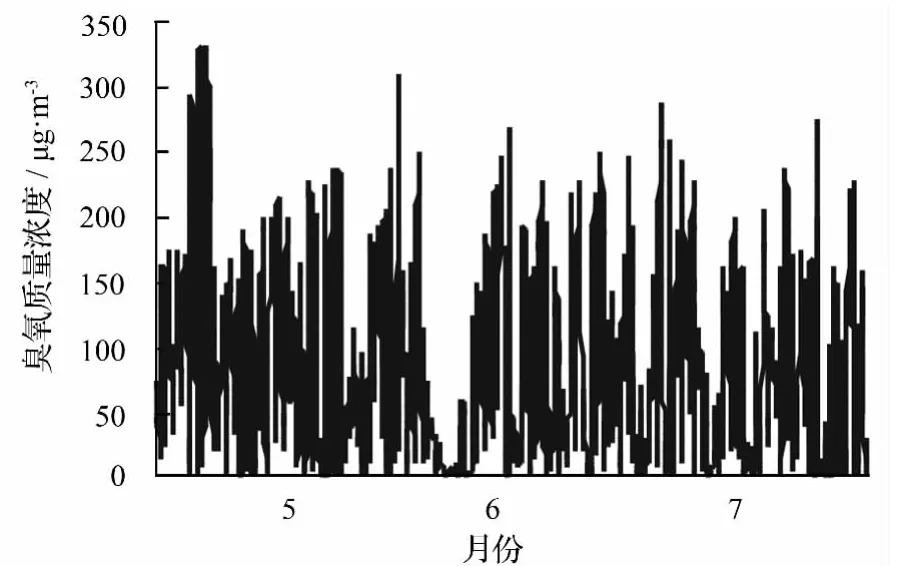
图3 臭氧质量浓度月变化

图4 臭氧质量浓度日变化
3.3 臭氧吸收
图5显示了4株杨树月均冠层对臭氧的导度(GO3)和整树臭氧吸收速率(FO3)。GO3和FO3呈现相同的变化趋势,最大值均出现在5月份,分别为996.06 和 610.59 nmol·m-2·s-1;从时间变化来看均为5月份>6月份>7月份;FO3由高到低依次为树1(126.45 nmol·m-2·s-1)、树 4(103.34 nmol·m-2·s-1)、树 2(58.52 nmol·m-2·s-1)、树 3(24.57 nmol·m-2·s-1);GO3由高到低依次为树 1(206.28 mmol·m-2·s-1)、树 4(168.59 mmol·m-2·s-1)、树 2(95.46 mmol·m-2·s-1)、树 3(40.08 mmol·m-2·s-1)。4株样树 GO3均值为(127.60±64.35)mmol·m-2·s-1,FO3均值为(78.22±39.45)nmol·m-2·s-1。
3.4 臭氧吸收对环境因子的响应
臭氧吸收是由冠层导度和外界臭氧质量浓度的相互作用决定的[7]。这导致了FO3的变化格局与外界臭氧质量浓度的变化格局接近,以2013年6月的月均值分析臭氧吸收对环境因子的响应(图6)。在一定的外界臭氧质量浓度下,冠层对GO3有效的调控了FO3,进而受水汽压亏缺(D)和太阳总辐射(Rs)的影响。对于研究的杨树GO3对D和Rs的响应方式相似,但是不同的杨树样株的GO3大小差别较大(图6B,C)。
空气水汽压亏缺(D)和风速与臭氧质量浓度呈正比关系,当D达最大值2.52 kPa时,臭氧质量浓度也达到最大值,为 225.46 μg·m-3,当风速达到最大的3.30 m·s-1时,臭氧质量浓度最大,为 342.45 μg·m-3,而太阳总辐射(Rs)为 28 W·m-2时,臭氧质量浓度开始下降。
GO3和FO3均随着 D、Rs和风速增大而增大(图6B、C、E、F、H、I),由图6可知,树1 表现的较为弱一些,树2至树4均较强烈,树4变化最明显。水汽压亏缺在1.52 kPa时GO3和FO3均出现了不同程度的降低,因此,在不考虑物种的情况下,D对更高的GO3和FO3更敏感,随着D增加臭氧吸收将减少。同时Rs在达到50 W·m-2时GO3和FO3也有所降低。然而,随着Rs增加臭氧吸收基本保持不变。
3.5 臭氧质量浓度与树干液流的关系
从24 h变化来看(图7),臭氧质量浓度日变化趋势呈单峰曲线,最高值出现在17:00,其值为333.00 μg·m-3,最小值出现在 07:00,仅为 6 μg·m-3。从07:00开始,臭氧质量浓度不断增大,从17:00开始缓慢下降,在夜间也有一个微弱的峰值,在0:00,其值为180 μg·m-3。
使用TDP测定树干液流,由每个时刻得到的树干液流温度差数据计算液流密度值。如图7所示,不同杨树液流密度呈单峰型变化趋势,不同样树日变化均值树 1 最大,为 0.002 6 g·cm-2·s-1,树 3 最小,仅为0.000 64 g·cm-2·s-1;从日变化时间来看,4株样树均在06:00液流密度开始上升,10:00—11:00达到一天中的最大值,树1在10:00达最大值,树2、树3和树4均在11:00达最大值;16:00液流密度开始缓慢下降,在22:00—06:00均处于较低值,在夜晚液流密度较低,树木蒸腾耗水较少。
图5 不同样树冠层的臭氧导度和臭氧吸收速率
如图7所示,臭氧质量浓度与液流密度的关系并不是一一对应的。当树1、树2、树3和树4分别在 06:00、06:00、02:00、04:00 处于一天的最小值,而臭氧质量浓度在07:00最小;当树1在10:00,树2、树3和树4均在11:00达最大值,而臭氧在17:00最大。当臭氧质量浓度为最低值时,液流密度值开始上升;当臭氧质量浓度值达最大值时,液流密度值开始缓慢下降,说明臭氧质量浓度大小影响液流速率的变化。
4 讨论
4.1 杨树整树臭氧吸收
研究发现基于树干液流技术确定树木年臭氧吸收量明显小于UFORE模型方法确定的值,这种区别表明在臭氧风险评估中应考虑特定树种的通量。本研究结果显示,杨树臭氧吸收量为3.75 g·m-2(4个研究样树累计气孔臭氧吸收通量均值78.22 mmol·m-2×48 g·mol-1/1 000=3.75 g·m-2),这比模型得到的结果低很多[28,5]。Nowak 等[4]29采用城市影响模型,并结合野外观测数据,发现美国几个城市树冠污染物年去除量均值为9.3 g·m-2,变化范围为6.6~12.0 g·m-2。臭氧在去除污染物中的比例为31.8%[4]29-33。因此,树木对臭氧的去除速率为 2.96 g·m-2(9.3 g·m-2×31.8%=2.96 g·m-2)。采用相同的模型,按照Yang等[5]指出的北京城市森林污染物去除量(27.5 g·m-2),臭氧在去除污染物中的比例为20%,可得出杨树对臭氧的吸收速率为5.50 g·m-2。由此可知,本研究野外观测的结果与模型方法的结果有巨大的差异,分析原因主要为:第一,树干液流测定的数据仅确定了进入叶片的臭氧通量,而模型的方法计算了总的干沉降(包括了气孔的吸收和树木外部非蒸腾表面的吸附);第二,两种方法的应用尺度不同,树干液流技术的臭氧吸收是基于冠层尺度,模型的方法是树木臭氧吸收或者去除通常是区域或者城市尺度,而且假定外界臭氧质量浓度均匀[5];第三,叶面积指数不同,在 UFORE模型中用于计算冠层气孔阻力的叶面积指数是4.30[3],而本研究的指数值是 3.26,显然该值比本研究的值高许多;第四,液流测量仅仅估计臭氧通量的叶片水平,建模方法计算总的干沉积,包括气孔臭氧的吸收和吸附的非蒸发外树的表面,这可能会导致更高的臭氧去除;第五,计算的空气动力学UFORE模型包含了沉积速度,准层流边界层和冠层阻力[29],但在本研究中,只计算冠层阻力。总之,这种差异表明在臭氧风险评估中考虑物种特异通量是必要的。

图6 臭氧质量浓度、冠层臭氧导度、臭氧吸收速率与水汽压亏缺、太阳总辐射和风速间的关系
4.2 杨树臭氧吸收的环境调控
由于林木吸收臭氧和蒸腾通过气孔行为耦合,林木水分利用分配格局的改变会导致其吸收臭氧格局改变。研究表明,林木夜间液流占全天总液流的比例通常为7%~10%[30],干旱沙漠植被两者的比例则高达30%~60%[31]。由此推断,城市林木水分利用分配格局亦不同。因此,城市环境下林木吸收臭氧格局可能改变:城市林木夜间吸收臭氧量占全天吸收臭氧总量的比例可能较高,从而导致林木受臭氧伤害风险增加。林木吸收臭氧特征主要取决于大气臭氧质量浓度和气孔导度[27]。
杨树臭氧吸收的变化是由于外界臭氧质量浓度和冠层导度的变化而引起的,在本研究地点,不同月份(5、6、7月份)臭氧质量浓度差异显著。这导致了杨树臭氧吸收速率和冠层对臭氧的导度与外界大气臭氧质量浓度有相近的变化格局。
调控杨树整树臭氧吸收的气孔开放程度主要受水汽压亏缺和辐射的影响,而气孔是限制整树臭氧吸收的关键因子[12,14]。杨树臭氧吸收量高是由于以下原因造成的。首先,在较低的水汽压亏缺条件下,气孔开放允许较高的臭氧进入叶片,即使在很低的外界臭氧质量浓度条件下。其次,低辐射条件下高的气孔导度加快了臭氧吸收,意味着城市中夜间、凌晨或者阴雨天气孔保持部分张开。大量的相关研究表明,一些物种的气孔夜间并不完全关闭,允许夜间蒸腾的进行。夏季较高的空气温度和较低的空气湿度都可能促使气孔在夜间开放。最后,研究阶段较高的土壤水分可利用性促进了臭氧吸收,对幼苗和枝条的试验证实了干旱胁迫下植物通过调控气孔的开放以避免臭氧的危害[32]。然而,在研究时段的5、6、7三个月份北京有大量的降水事件发生,杨树得到了及时的灌溉,土壤水分含量没有限制气孔的开放。
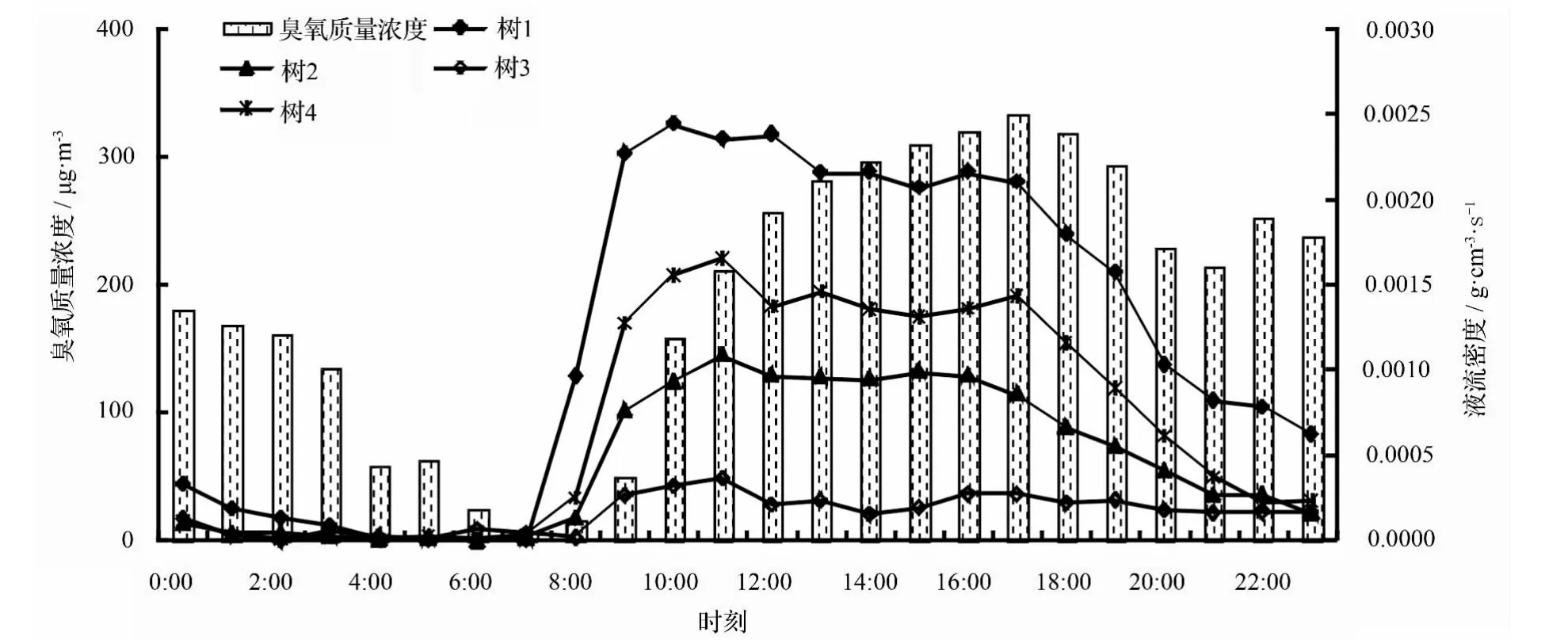
图7 臭氧质量浓度与液流密度的关系
4.3 杨树冠层臭氧与树干液流关系
Matyssek等[10]通过研究夜间林木通过气孔途径吸收臭氧,指出臭氧对树木造成较大伤害,因而建议评价臭氧引起林木伤害的指标应考虑夜间林木吸收臭氧量。林木吸收臭氧季节变化格局差异性较大,如:北京市典型绿化树种吸收臭氧速率夏季较高[7];而 Wieser等[11]对加那利松(P.canariensis)吸收臭氧速率的研究,指出季节变化不明显。Matyssek等[10]对林木吸收臭氧的年际格局变化进行了研究,指出年际变化格局较大。不同功能型的树种吸收臭氧格局互补,在不同的气候条件下维持稳定的群落吸收臭氧功能。
臭氧质量浓度与不同杨树树干液流并不一一对应,而是滞后于液流密度的峰值。树干液流是树木蒸腾耗水的表征,它与光合有效辐射、温度和湿度等环境因子有关。气温与臭氧质量浓度呈正相关,温度越高,臭氧质量浓度也越大,但过高的臭氧质量浓度会抑制光合,从而抑制树木蒸腾。因此,当臭氧质量浓度最大的时候,液流密度开始减小、树木蒸腾变小;相反,当臭氧质量浓度最小时,液流密度开始上升,说明臭氧质量浓度的高低影响液流密度的变化。
由TDP得到液流密度,进而推导计算臭氧吸收速率和臭氧冠层导度。结果显示,不同月份的树干液流密度表现为树1>树4>树2>树3,而臭氧吸收速率也表现为树1>树4>树2>树3,二者大小排序完全一致。当液流密度处于一天的最大值时,臭氧吸收速率也为一天中的最大值;当液流密度最小时,臭氧吸收速率也处于最小值。杨树不同样株树1、树2、树3和树4均有相同的变化规律:液流密度增大时,臭氧吸收速率也随之增大,液流密度减小时,臭氧吸收速率也随之减小,一天中各个时刻的值大小均一一对应,4株杨树中树3表现的最为明显,树1表现稍显弱一些,这与树1胸径较小(19.74 cm)、蒸腾较小有关。说明基于树干液流技术得到的液流密度、计算出冠层气孔导度,结合外界的臭氧质量浓度,能够估算不同树形级别杨树的臭氧吸收量,该方法可以适用于衡量不同环境下植物的臭氧吸收量,液流密度的变化决定了臭氧吸收速率的变化。
4.4 臭氧质量浓度超过临界水平引起树木的响应
相关研究采用基于暴露浓度和通量的临界水平(AOT40和AFst1.6)以衡量研究地点臭氧对树木的潜在影响。本研究采用基于暴露浓度和通量的临界水平时,研究地点杨树受臭氧危险的潜在风险较高。生长季杨树 AOT40 为 72.02 mg·m-3·h-1,远远超过对树木危害的臭氧质量浓度的临界水平AOT40 10.71 mg·m-3·h-1[7]。采用基于通量的临界水平,落叶树种AFst1.6值高于对树木产生危害的臭氧通量的临界水平 4 mmol·m-2[7]。但是,常绿树种AFst1.6值低于对树木产生危害的臭氧通量的临界水平。这些研究结果表明,与城市中的常绿树种相比,落叶树种更容易受到臭氧危害的影响。与本研究结果一致的是,地中海地区常绿灌木比落叶灌木对臭氧胁迫的耐受能力更强[33]。此外,本研究结果表明,基于测量更适合于臭氧对城市树木的影响比风险暴露法评价通量。这些结果与以往的研究相比,臭氧通量比AOT40曝光的预测臭氧可见叶面损伤的测量[34]更准确一致。
5 结论
臭氧吸收量受到水汽压亏缺和太阳总辐射等环境因子的影响,气孔的开放促进杨树臭氧吸收,臭氧质量浓度影响液流密度变化。本研究从速率、效率及环境因子三个方面揭示城市环境下林木整树吸收臭氧功能变化的信息,D对更高的GO3和FO3更敏感,随着D增加臭氧吸收减少;臭氧质量浓度超过临界水平引起树木相应的响应,杨树AOT40为72.02 mg·m-3·h,远超过对树木危害的臭氧质量浓度的临界水平10.71 mg·m-3·h,说明了杨树的高臭氧风险,树干液流技术适用于计算植物的臭氧吸收量。结合林木冠层气孔导度响应环境变化的敏感性,阐明林木吸收臭氧速率、效率及格局对城市环境的响应机制,既是对城市生态学有关环境改变影响林木吸收臭氧特征的前沿性探索,又可对城市林木吸收臭氧速率和效率提高的问题提出新的解释。
[1]Wu Jianguo.Making the case for landscape ecology:an effective approach to urban sustainability[J].Landscape Journal,2008,27(1):41-50.
[2]Chan C K,Yao Xiaohong.Air pollution in mega cities in China[J].Atmospheric Environment,2008,42(1):1-42.
[3]Nowak D J,Crane D E.Carbon storage and sequestration by urban trees in the USA[J].Environmental Pollution,2002,116(3):381-389.
[4]Nowak D J,Dwyer J F.Understanding the benefits and costs of urban forest ecosystems[M].New York:Springer,2007:25-46.
[5]Yang J,McBride J,Zhou J,et al.The urban forest in Beijing and its role in air pollution reduction[J].Urban Forestry & Urban Greening,2005,3(2):65-78.
[6]Manes F,Incerti G,Salvatori E,et al.Urban ecosystem services:tree diversity and stability of tropospheric ozone removal[J].Ecological Applications,2012,22(1):349-360.
[7]Wang Hua,Zhou Weiqi,Wang Xiaoke,et al.Ozone uptake by adult urban trees based on sap flow measurement[J].Environmental Pollution,2012,162(1):275-286.
[8]Wieser G,Matyssek R,Köstner B,et al.Quantifying ozone uptake at the canopy level of spruce,pine and larch trees at the alpine timberline:an approach based on sap flow measurement[J].Environmental Pollution,2003,126(1):5-8.
[9]Musselman R C,Minnick T J.Nocturnal stomatal conductance and ambient air quality standards for ozone[J].Atmospheric Environment,2000,34(5):719-733.
[10]Matyssek R,Günthardt-Goerg M S,Maurer S,et al.Nighttime exposure to ozone reduces whole-plant production in Betula pendula[J].Tree Physiology,1995,15(3):159-165.
[11]Wieser G,Luis,VC,Cuevas E.Quantification of ozone uptake at the stand level in a Pinus canariensis forest in Tenerife,Canary Islands:An approach based on sap flow measurements[J].Environmental Pollution,2006,140(3):383-386.
[12]Emberson L D,Wieser G,Ashmore M R.Modelling of stomatal conductance and ozone flux of Norway spruce:comparison with field data[J].Environmental Pollution,2000,109(3):393-402.
[13]Wieser G,Häsler R,Götz B,et al.Role of climate,crown position,tree age and altitude in calculated ozone flux into needles of Picea abies and Pinus cembra:a synthesis[J].Environmental Pollution,2000,109(3):415-422.
[14]Nunn A J,Wieser G,Metzger U,et al.Exemplifying whole-plant ozone uptake in adult forest trees of contrasting species and site conditions[J].Environmental Pollution,2007,146(3):629-639.
[15]Köstner B,Matyssek R,Heilmeier H,et al.Sap flow measurements as a basis for assessing trace-gas exchange of trees[J].Flora-Morphology,Distribution,Functional Ecology of Plants,2008,203(1):14-33.
[16]Braun S,Schindler C,Leuzinger S.Use of sap flow measurements to validate stomatal functions for mature beech(Fagus sylvatica)in view of ozone uptake calculations[J].Environmental Pollution,2010,158(9):2954-2963.
[17]王华,欧阳志云,任玉芬,等.基于树干液流技术的北京市刺槐冠层吸收臭氧特征研究[J].生态学报,2013,33(23):7323-7331.
[18]金莹,张志强,方显瑞,等.杨树人工林生态系统通量贡献区分析[J].生态学报,2012,32(12):3966-3974.
[19]张津林,张志强,查同刚,等.沙地杨树人工林生理生态特性[J].生态学报,2006,26(5):1523-1532.
[20]申李华.杨树人工林树干液流动态变化与林分蒸腾[D].北京:北京林业大学,2007.
[21]Granier A.Use nouvelle method pour lamesure du flux desève brute dansla tronc des arbres[J].Annales des Sciences forestières,1985,42(2):193-200.
[22]赵平,饶兴权,马玲,等.马占相思(Acacia mangium)树干液流密度和整树蒸腾的个体差异[J].生态学报,2006,26(12):4050-4058.
[23]Monteith J L.A reinterpretation of stomatal response to humidity[J].Plant,Cell and Environment,1995,18(4):357-364.
[24]Campbell G S,Norman J M.An introduction to environmental biophysics[M].New York:Springer-Verlag,1998:36-51.
[25]Granier A.Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements[J].Tree Physiology,1987,3(4):309-320.
[26]Köstner B,Schulze E D,Kelliher F M,et al.Transpiration and canopy conductance in a pristine broad-leaved forest of Nothofagus:an analysis of xylem sap flow and eddy correlation measurements[J].Oecologia,1992,91(3):350-359.
[27]Wieser G,Havranek W M.Environmental control of ozone uptake in Larix decidua Mill.:a comparison between different altitudes[J].Tree Physiology,1995,15(4):253-258.
[28]Nowak D J,Grane D E.The urban forest effects(UFORE)model:quantifying urban forest structure and functions[C].Boise:North Central Research Station,St.Paul,MN,2000:714-720.
[29]Nowak D J,Civerolo K L,Trivikrama Rao S,et al.A modeling study of the impact of urban trees on ozone[J].Atmospheric Environment,2000,34(10):1601-1613.
[30]Zeppel M,Tissue D,Taylor D,et al.Rates of nocturnal transpiration in two evergreen temperate woodland species with differing water-use strategies[J].Tree Physiology,2010,30(8):988-1000.
[31]Snyder K A,Richards J H,Donovan L A.Night-time conductance in C3 and C4 species:do plants lose water at night?[J].Journal of Experimental Botany,2003,54:861-865.
[32]Wieser G,Havranek W M.Ozone uptake in the sun and shade crown of spruce:quantifying the physiological effects of ozone exposure[J].Trees,1993,7(4):227-232.
[33]Calatayud V,Marco F,Cerveróa J,et al.Contrasting ozone sensitivity in related evergreen and deciduous shrubs[J].Environmental Pollution,2010,158(12):3580-3587.
[34]Gerosa G,Marzuoli R,Desotgiu R,et al.Visible leaf injury in young trees of Fagus sylvatica L.and Quercus robur L.in relation to ozone uptake and ozone exposure.an open-top Chambers experiment in south Alpine environmental conditions[J].Environmental Pollution,2008,152(2):274-284.
