小麦品种太空6号和中育6号对禾谷孢囊线虫Ha43致病型的抗性遗传分析
2015-06-27石颖斐胡艳峰代君丽李洪连
石颖斐,胡艳峰, 代君丽,李洪连
(河南农业大学植物保护学院,河南 郑州 450002)
小麦品种太空6号和中育6号对禾谷孢囊线虫Ha43致病型的抗性遗传分析
石颖斐,胡艳峰, 代君丽,李洪连
(河南农业大学植物保护学院,河南 郑州 450002)
分别构建了感病亲本温麦19同太空6号和中育6号的杂交组合F2代分离群体。禾谷孢囊线虫(Cereal cyst nematode,CCN )须水群体的致病型为一种新致病型Ha43,为了明确太空6号和中育6号对Ha43致病型的抗性遗传特点,采用主基因+多基因混合遗传模型(mixed major gene plus poly-gene inheritance model)分析法,分析了温麦19×太空6号和温麦19×中育6号杂交组合F2代群体的抗性分离情况。结果表明,太空6号和中育6号对Ha43的抗性遗传效应均表现为数量性状遗传。太空6号对Ha43的抗性遗传效应符合B-1模型,推测太空6号对CCN的抗性由2对主基因+多基因控制,主基因表现为加性-显性-上位性效应,抗性主基因遗传率为95.40%。中育6号对Ha43的抗性遗传效应符合B-2模型,推测中育6号对CCN的抗性由2对主基因+多基因控制,主基因表现为加性-显性效应,抗性主基因遗传率为90.31%。
太空6号;中育6号;禾谷孢囊线虫; Ha43须水群体; 抗性遗传
禾谷孢囊线虫(Cereal cyst nematode,CCN) 1874年在德国首次发现,现已经传播至世界上50多个国家[1]。其在西亚、北非、欧洲、澳大利亚和美国都造成严重的经济损失[1,2]。中国1989年在湖北省天门县的小麦上首次发现,已在湖北、江苏、安徽、山东、河南、山西、北京、陕西、天津、新疆和西藏[3]等16个省(自治区、直辖市)发生,给当地小麦生产造成了严重危害。河南省作为中国重要的小麦产区,最早于1990年在安阳市发现该病,近年为害日益严重,据统计发病面积约130万hm2,重病地块达10多万hm2,小麦产量损失为28.8%~35.6%[4]。禾谷孢囊线虫是一个复合种群,已报道有12个已定名的种类和一些尚未定名的种[5]。这些种类中,燕麦孢囊线虫Heteroderaavenae是全球分布范围最广的寄生线虫[6,7]。其主要危害小麦、大麦、黑麦、燕麦等禾谷类作物及多种禾本科牧草,危害植物根部,使根系变成球状瘤节和须根团,不能产生根毛,影响作物对水分和营养物质的吸收,抑制作物的生长从而影响产量。目前,防治禾谷孢囊线虫病最为经济有效的措施是种植抗病品种[8],许多国家应用该措施防治CCN已经取得了很好的效果[9]。同时,该措施不需要额外的设备和成本,易于大面积推广[2,10]。CCN存在多种致病型,新致病型的出现往往使得抗病品种的应用受到限制,因为不同的CCN致病型对不同的寄主和栽培品种的毒力不同[11-13]。因此,选育对当地CCN致病型抗病的小麦品种显得尤为重要。YUAN等[14]发现禾谷孢囊线虫郑州须水群体,能够侵染A组的Ha1和Ha2使其表现感病, 不能使Ha3感病,不同于与其最相近的Ha13群体。该群体也能侵染B组的一些大麦品种,2个燕麦品种和小麦品种Capa使其表现感病,毒性谱较广。因此确定该群体为H.avenae一个新致病型,并定名为Ha43。付博等[15]在用RFLP分子标记法和rDNA-ITS序列分析法分析黄淮麦区主要禾谷孢囊线虫致病型时,发现郑州须水群体与荥阳群体有差别。吴绪金[4]和孙君伟[16]对太空6号和中育6号进行了多年抗性鉴定,发现太空6号和中育6号对H.avenae须水群体具有较好的抗性且抗性稳定,了解它们对Ha43的抗性遗传规律对于进一步利用其抗性具有重要的价值和意义。盖钧镒等[17]提出的主基因+多基因遗传模型,既能够检测和鉴定数量性状主基因和多基因的存在,又能够对方差和基因效应等遗传参数进行估计。国内已经有很多报道采用该模型对作物的数量性状遗传规律进行了研究[18,19]。宗莹莹等[20]和代君丽等[21]的研究发现,中育6号和太空6号对燕麦孢囊线虫荥阳群体和菲利普孢囊线虫焦作群体的抗性呈数量性状遗传。本研究拟采用主基因+多基因混合遗传模型分析太空6号和中育6号对燕麦孢囊线虫新致病型Ha43的抗性遗传特点,以明确它们对Ha43的抗性遗传规律,为小麦抗CCN基因的标记定位及其辅助育种提供理论依据。
1 材料与方法
1.1 供试材料
本试验构建的F2代分离群体是分别以抗CCN品种中育6号和太空6号为父本,以高感CCN品种温麦19为母本,杂交产生F1代,再由F1代自交产生F2代。其中,中育6号系中国农科院棉花研究所小麦育种室育成的小麦新品种,品质优良,具有良好的抗病性和抗逆性。太空6号系河南省农业科学院小麦研究所育种研究室和中国航天科技集团合作,经太空诱变后连续多年选育而成。
1.2 田间病圃抗性鉴定与遗传分析
本试验于2012年10月至2013年5月在郑州市须水崔寨(经度113.49°,纬度34.81°,郑州市中原区)进行,该试验田土质为沙壤土,肥力较好,常年种植小麦,小麦孢囊线虫病发生严重,上茬作物为玉米。感病亲本温麦19,抗病亲本太空6号和中育6号,每个品种种植一个小区,小区每行长2 m,宽1 m,行距20 cm,每行播种60粒,温麦19×太空6号杂交组合F2代群体和温麦19×中育6号杂交组合F2代群体各组合播种260粒左右。小麦生长期间采用常规栽培管理措施,于2013年5月调查取样,对温麦19、太空6号和中育6号在每个小区内前、中、后随机选取3点,每点至少10株,调查单株根部以及根系周围土壤中的孢囊量,根据品种平均单株孢囊量,进行方差分析,并判断其抗病性[22]。对2个组合的F2代分离群体,直接根据单株根部及根系周围土壤中的孢囊数确定抗性情况,如果F2代单株抗感分离比率符合孟德尔遗传定律,则说明为质量性状抗性,存在主效基因;若F2代抗感分离比率呈现正态分布,则说明为数量性状抗性,存在QTL。
1.3 数据统计分析

2 结果与分析
2.1 温麦19×太空6号组合和温麦19×中育6号组合亲本和F2代抗性分析
在温麦19×太空6号组合中,温麦19的平均单株孢囊量为23.059个,太空6号的平均单株孢囊量为2.251个,独立样本t检测值表明,两亲本的CCN单株孢囊量表现出显著性差异(P<0.05) (表1),说明温麦19和太空6号存在显著的遗传差异。在温麦19×中育6号杂交组合中,温麦19的平均单株孢囊量为23.059个,中育6号的平均单株孢囊量为2.858个,独立样本t检测表明,两亲本的CCN单株孢囊量也表现出显著性差异(P<0.05) (表1),说明温麦19和中育6号也存在显著的遗传差异。对2个杂交组合F2代不同单株的孢囊量进行统计,制作频数分布图(图1)。从图1看出,2个杂交组合F2代单株孢囊量均呈单峰连续分布。偏度和峰度检验(表1)表明,这2个杂交组合的F2代单株孢囊量均呈偏态分布,这说明太空6号和中育6号对Ha43的抗性均属数量性状遗传,但存在明显的主基因。

表1 亲本及F2代单株孢囊量的分析Table 1 Analysis of the cyst number per plant in the parents and F2 generation

注: A: 温麦19×太空6号; B: 温麦19×中育6号。Notes: A: Cross Wenmai 19 and Taikong No.6; B: Cross between Wenmai 19 and Zhongyu No.6.
2.2 遗传模型的筛选

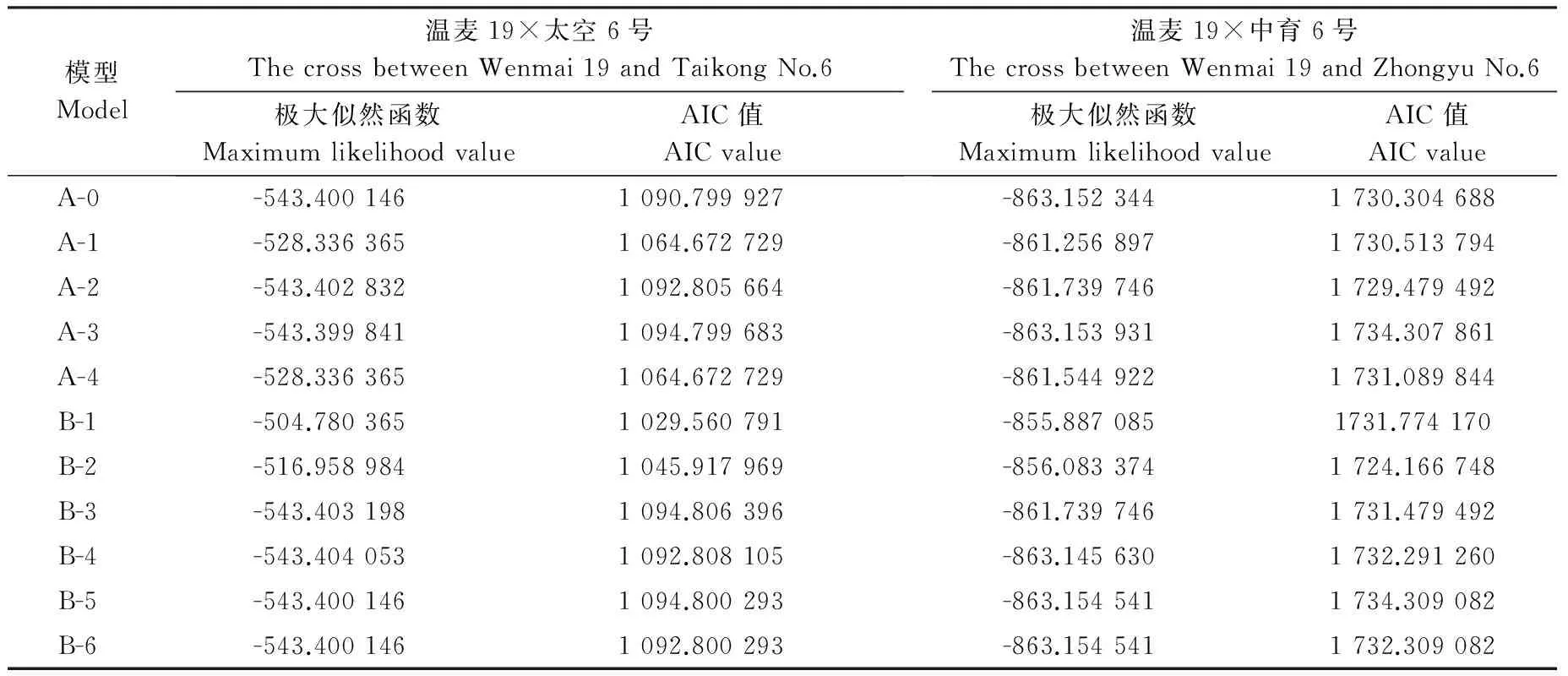
表2 温麦19×太空6号组合和温麦19×中育6号组合F2群体对Ha43抗性在不同遗传模型下的极大似然函数和 AIC值Table 2 Maximum likelihood values and AIC values in different genetic models for CCN resistance in the F2 population derived from the cross between Wenmai 19 and Taikong No.6 and the cross between Wenmai 19 and Zhongyu No.6
注:11个遗传模型可以分为3类,第1类为无主基因模型,包含1个遗传模型A-0,第2类为1对主基因模型,包含4个遗传模型A-1、A-2、A-3和A-4,第3类为2对主基因遗传模型,包含6个遗传模型B-1、B-2、B-3、B-4、B-5和B-6。
Notes: The genetic models could be divided into three types. The first type is none major gene model, including one model A-0. The second type is one major gene model, including four models A-1,A-2,A-3 and A-4. The third type is two major genes model, including six models B-1,B-2,B-3,B-4,B-5 and B-6.

表3 温麦19×太空6号组合和温麦19×中育6号组合F2代群体CCN 抗性遗传模型的适合性检验Table 3 Fitness of genetic models for CCN resistance in the F2 population derived from the cross between Wenmai 19 and Taikong 6 and the cross between Wenmai 19 and Zhongyu 6


2.3 最适遗传模型相关参数估计
根据最适遗传模型的极大似然估计值, 估算最适遗传模型的一阶、二阶遗传参数(表 4)。结果表明, 温麦19×太空 6号杂交组合在大田病圃试验的环境条件下抗Ha43的2 对主基因(A和B)加性效应均表现为正向效应(da>0,db>0),主基因显性效表现为负向效应(ha<0,hb<0),主基因之间有正向的加性×加性效应和显性×显性效应(i>0,l>0),主基因之间有负向的加性×显性效应和显性×加性效应(jab<0,jba<0)。温麦19×中育6号杂交组合在大田病圃实验的环境条件下抗Ha43的2对主基因(A和B)加性效应均表现为正向效应(da>0,db>0),主基因显性效均表现为负向效应(ha<0,hb<0),主基因之间不存在加性×加性、加性×显性、显性×加性、显性×显性效应。温麦19×太空6号杂交组合对其抗性的主基因遗传率为95.40%,温麦19×中育6号杂交组合对其抗性的主基因遗传率为为90.31%,说明太空6号和中育6号在大田病圃试验中对Ha43的抗性均有很高的遗传力,该抗性性状能够稳定地从亲本遗传给子代。

表4 温麦19×太空6号和温麦19×中育6号F2群体CCN 抗性遗传参数估计值Table 4 Estimates of genetic parameters for CCN resistance in the F2 population derived from the cross between Wenmai 19 and Taikong No.6 and the cross between Wenmai 19 and Zhongyu No.6

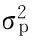
3 讨论
主基因+多基因混合遗传模型综合了分离世代的整体信息,避免了人为划分抗感界限,可以减少个体分级的主观性。目前,国外在抗CCN遗传分析方面的报道较少,已有的研究多是针对小麦近缘种属大麦和燕麦进行了抗禾谷孢囊线虫遗传分析,结果显示大麦和燕麦对禾谷孢囊线虫的抗性多呈质量性状特征[23~25]。高秀[26]利用Madsen与烟农21、晋麦47、徐麦856和龙辐麦10号等感病小麦品种配制杂交组合,F2代对菲利普孢囊线虫抗病和感病植株的分离比例均符合3∶1的比例,表明Madsen对菲利普孢囊线虫的抗病基因可能受一对显性单基因控制。代君丽等[21]分析了感病品种豫麦 47和抗病品种太空 6 号的杂交 F2代群体在室内接种二龄幼虫和田间病圃中对H.avenae郑州群体的抗性遗传规律,结果表明抗性遗传效应均符合 B-2 模型,既2对主基因+多基因模型,且主基因表现为加性-显性效应。推测太空6号对H.avenae的抗性由 2 对主基因+多基因控制, 主基因表现为加性-显性效应。这和本研究中太空6号对Ha43的抗性遗传效应由2对主基因+多基因控制,但主基因表现为加性-显性-上位性的结果不一致。宗莹莹等[20]采用主基因+多基因混合遗传模型分析方法,分析了温麦19×中育6号杂交组合F2代群体对菲利普孢囊线虫(H.filipjevi)焦作博爱群体和燕麦孢囊线虫(H.avenae)荥阳群体抗性分离状况,结果表明中育6号对两种禾谷孢囊线虫的抗性表现均为数量性状遗传,且由一对主基因控制。这和本研究中中育6号对Ha43的抗性遗传效应由2对主基因+多基因的结果不一致。这可能是因为在大田病圃试验中,线虫的分布不均匀,受环境的影响较大所致;也可能因为须水群体 Ha43与荥阳群体和焦作博爱群体的致病型不同[19,20],导致中育6号对其抗性遗传特点有差别。下一步需要进行室内盆栽试验,并构建这2个品种和温麦19组合的遗传图谱,定位出这2个品种中抗Ha43致病型的抗病基因或QTLs,为加速抗禾谷孢囊线虫的小麦新品种的选育提供更加有效的信息。
[1] EVANS K, TRUDGILL D L, WEBSTER J M. Plant Parasitic Nematodes in Temperate Agriculture [M]. Wallingford :CAB International Press, 1993: 259-303.
[2] RIVOAL R, NICOL J M. Past research on the cereal cyst nematode complex and future needs [C]//REY T, NICOL J M, DABABAT A A. Cereal Cyst Nematodes: Status, Research and Outlook. Ankara: CIMMYT Press, 2009:100-105.
[3] 李惠霞,柳永娥,魏 庄,等.新疆和西藏发现禾谷孢囊线虫[C]//中国线虫学研究(第四卷), 北京:中国农业科学技术出版社,2012.
[4] 吴绪金.小麦禾谷孢囊线虫病产量损失估计及抗病性研究[D].郑州:河南农业大学,2007.
[5] HANDOO Z A. A key and compendium to species of theHeteroderaavenaegroup(Nematoda:Heteroderidae) [J].Journal of Nematology, 2002, 34:250-202.
[6] NICOL J M, RIVOAL R. Global knowledge and its application for the integrated control and management of nematodes on wheat [C]//CIANCIO A, MUKERJI K G. Integrated Management and Biocontrol of Vegetable and Grain Crops Nematodes. Dordrecht: Springer, 2007:251-294.
[7] SMILEY R W, NICOL J M. Nematodes which challenge global wheat production[C]//CARVER B F. Wheat: science and trade. Hong Kong: Printed and Bound in Singapore, 2009:171-187.
[8] HOLGADO R, ANDERSSON S, MAGNUSSON C. Management of cereal cyst nematodesHeteroderaspp. in Norway [J].Communications in Agricultural and Applied Biological Sciences, 2006,71: 639-645.
[9] FISHER J M, RATHJENA J, DUBE A J. Tolerance of Commercial Cultivars and Breeders’ Line of Wheat toHeteroderaavenaeWoll[J].Journal of Nematology,1981,32(4):545-551.
[10]NICOL J M, OGBONNAYA F, SINGH A K, et al. Current global knowledge of the usability of cereal cyst nematode resistant bread wheat germplasm through international germplasm exchange and evaluation[C]//RILEY I T, NICOL J M, DABABAT A A. Cereal cyst Nematodes: Status, Research and Outlook, Ankara: CIMMYT Press, 2009:149-153.
[11]ADDABBO T, SASANELLI N. Race and resistance studies on two Italian populations ofHeteroderaavenae[J]. Nematologica Mediterranea, 1994, 22:79-82.
[12]NICOL J M, ELEKCIOGLU I H, BOLAT N, et al. The global importance of the cereal cyst nematode (Heteroderaspp.) on wheat and international approaches to its control [J]. Communications in Agricultural and Applied Biological Sciences, 2007, 72: 677-686.
[13]MOKABLI A, VALETTE S, GAUTHIER J P, et al. Variation in virulence of cereal cyst nematode populations from North Africa and Asia [J].Nematology, 2002, 4:521-525.
[14]YUAN H X, SUN J W, YANG W X, et al. New pathotypes ofHeteroderaavenae(cereal cyst nematode) from winter wheat in Zhengzhou, Henan, China [J].Australasian Plant Pathology, 2010, 39: 107-111.
[15]FU B, YUAN H X, ZHANG Y, et al. Molecular characterization of cereal cyst nematodes in winter wheat on the Huang-Huai floodplain of China using RFLP and rDNA-ITS sequence analyses[J]. Australasian Plant Pathology, 2011, 40: 277-285.
[16]孙君伟.黄淮麦区四个禾谷孢囊线虫群体种类和致病型鉴定及主推品种的抗性评价[D].郑州:河南农业大学,2010.
[17]盖钧镒,章元明,王建康. 植物数量性状遗传体系[M].北京:科学出版社,2003:96-102.
[18]郑 义,陆维忠,陈建民,等. 大麦赤霉病新抗源盐96157抗性的遗传分析[J]. 麦类作物学报,2008,28(2):329-333.
[19]李 华,刘联正,杨兴圣,等.小麦抗源材料0911-3抗白粉病的主基因+多基因遗传分析[J].华北农学报,2013,28(1):37-43.
[20]宗莹莹,代君丽,袁虹霞,等.普通小麦品种中育6号对两种禾谷孢囊线虫的抗性遗传分析[J].麦类作物学报,2013,33(2):249-254.
[21]代君丽,崔 磊,刘 珂,等.小麦品种太空6号对Heteroderaavenae郑州群体的抗性遗传分析[J].作物学报,2013,39(4):642-648.
[22]袁虹霞,张福霞,张佳佳,等.CIMMYT小麦种质资源对菲利普孢囊线虫(Heteroderafilipjevi)河南许昌群体的抗性[J].作物学报,2011,37(11):1956-1966.
[23]O'BRIEN P C, SPARROW D H B, FISHER J M. Inheritance of Resistance toHeteroderaavenaein Barley [J].Nematologica, 1979, 25(3):348 -352.
[24]COTTEN J, HAYES J D. Genetic studies of resistance to the cereal cyst nematode (Heteroderaavenae) in oats (Avena spp.)[J].Euphytica, 1972, 21:538-542.
[25]CLAMOT G, RIVOAL R. Genetic resistance to cereal cyst nematodeHeteroderaAvenae[J].Euphytica, 1984, 33:27-32.
[26]高 秀.小麦禾谷孢囊线虫(Heteroderafilipjevi和H.avenae)新抗源抗性研究[D].秦皇岛:河北科技师范学院,2012.
(责任编辑:蒋国良)
Genetic analysis of resistance to Ha43 ofHeteroderaavenaein wheat cultivars Taikong No. 6 and Zhongyu No. 6
SHI Yingfei, HU Yanfeng, DAI Junli, LI Honglian
(College of Plant Protection, Henan Agricultural University, Zhengzhou 450002,China)
In this research, Taikong No.6 and Zhongyu No.6 were crossed with the susceptible cultivar Wenmai 19 to produce F2 segregating population, respectively. The Xushui population ofHeteroderaavenaehad been identified as a new pathotype Ha43. In order to confirm the genetic characteristics of resistance to CCN in wheat cultivars Taikong No.6 and Zhongyu No.6, segregation characters of resistance among F2 progeny of crosses of Wenmai 19×Taikong No.6 and Wenmai 19×Zhongyu No.6 were analyzed. The method of mixed major gene plus poly-gene inheritance model was used to analyze the inheritance of resistance to Ha43. The results showed that the genetic model B-1 was most fitted for the resistance for the cross of Wenmai 19×Taikong No.6, which indicated that the resistance to cereal cyst nematode in Taikong No.6 was controlled by two major genes with additive-dominant-epistasis effects plus polygene effect. The major gene heritability of resistance for the cross of Wenmai 19×Taikong No.6 to CCN in the field was 95.40%. The genetic model B-2 was most fitted for the resistance for the cross of Wenmai 19×Zhongyu No.6, which indicated that the resistance to cereal cyst nematode in Zhongyu No.6 was controlled by two major genes with additive-dominant effects plus polygene effect. The major gene heritability of resistance for the cross of Wenmai 19×Zhongyu No.6 to CCN in the field was 90.31%.
Taikong No.6; Zhongyu No.6; cereal cyst nematode; Ha43; resistance inheritance
1000-2340(2015)01-0068-06
2014-07-19
国家自然科学基金项目(30971899);国家公益性(农业)科研专项(200903040-4);国家高技术研究发展计划(863计划)(2012AA101105)
石颖斐(1991-),女,河南平顶山人,硕士研究生,主要从事小麦禾谷孢囊线虫病研究。
代君丽(1977-),女,陕西西安人,副教授。
S435
A
