从传粉模式、花序与壳斗特征来看壳斗科的演化
2015-06-24陈曦
陈 曦
(云南大学 生态学与环境学院,云南 昆明 650091)
从传粉模式、花序与壳斗特征来看壳斗科的演化
陈 曦
(云南大学 生态学与环境学院,云南 昆明 650091)
壳斗科目前包括10个属,1 000 多个物种,是重要的经济类群.结合化石证据、分子系统发育学和形态学,壳斗科的进化研究主要集中于3个关键的生殖特征:传粉模式、花序结构和壳斗类型.从复杂到简单花序,虫媒到风媒传粉,以及二歧聚伞到花朵壳斗的演化是壳斗科进化的3个代表特征,与壳斗科的迁移与扩散过程息息相关.其中,具有先进风媒传粉和特化花序的栎属,与保持传统虫媒传粉和复杂花序的石栎属同时具有花朵壳斗这一现象是值得探讨的进化学问题.
壳斗科;进化;传粉综合征;花序;壳斗
壳斗科是北半球各种类型森林系统中的重要组成成分.在北美和中美的温带和半干旱地区栎属(Quercus)植物丰富度尤为高(世界415种,北美中美215种);而温暖湿润的东南亚地区有着丰富的石栎属(Lithocarpus)和栲属(Castanopsis)[1].壳斗科丰富的化石记录不仅可以与其现代分布相比较,更可以用于构建和评估该科起源,迁移和地理分隔模式的各种假说.这些特点使壳斗科成为一个无法优越的生物地理学研究模型.虽然栎属的研究相对系统详尽,但壳斗科的系统发育和宏观进化问题复杂,仍是生物进化学上的研究热点和难点之一.
1 壳斗科的分类现状
壳斗科目前包括10个属和超过1000个物种,是壳斗目(Fagales)下最大并且最具有经济重要性的一个类群[2].传统分类系统将壳斗科可分为3个亚科,栎亚科(Quercoideae)、水青冈亚科(Fagoideae)和栗亚科(Castaneoideae)[2](见表1).栎亚科仅包括栎属(QuercusL.)一属.水青冈亚科包括水青冈属(FagusL.)、中国三棱栎属(Formanodendron(Camus)Nixon & Crepet)、南美三棱栎属(Colombobalanus(Lozano,Hdz-C.& Henao)Nixon & Crepet)和三棱栎属(TrigonobalanusForman).栗亚科包括栗属(CastaneaL.)、栲属(CastanopsisSpach.)、黄叶柯属(ChrysolepisHjelmquist)和石栎属(LithocarpusBl.).但是,分子系统分类学的研究指出水青冈亚科在风媒传粉这一进化特征上的多源性.最近的系统发生学研究也否定了栗亚科是单系群这一传统说法.因此,对于壳斗科的亚科系统的使用需要慎重,或者进行进一步的调整.另外,2008年,研究者发现长期以来被鉴别是石栎属在北美唯一物种的Lithocarpusdensiflorus,实际与栎属,栗属和栲属的关系更加接近[3].因此,L.densiflorus现在被单立为壳斗科中的第10个属,Northolithocarpus(见表1).
2 传粉综合征、花序和壳斗的演化是壳斗科分化和进化的关键
壳斗科各属的分化与其多样化的形成是由动物散播种子,子叶留土幼苗,风媒传粉花序的进化以及单果实结合不分瓣壳斗结构等一系列特殊形态演化过程组合下推动促进产生的[2].壳斗科各属的无性形态因植物生长形态如生物量、生产力、死亡率等生态特征的相似而类似.而壳斗科多样的生殖特征的差异,被广泛应用于进化动态和分类的研究中.其中壳斗科的花序和壳斗显著的多样性和其传粉综合征的多态性(风媒和虫媒传粉)塑造了该科的独特性.首先,壳斗科的花序主要是葇夷与穗状花序,众多无柄或有柄的花朵聚集在一起形成聚伞或二歧聚伞花序.壳斗科展示了从复杂到简单,两性到单性一系列多样的花序和花朵结构,它们是研究壳斗科分化的和进化的重要材料.其次,传粉综合征是与吸引特定类型传粉者相关的花部特征的组合,其进化与花序的进化密切相关.壳斗科的传粉综合征的多态性在以风媒为传粉模式的壳斗目下显得尤为特殊.因此,了解壳斗科中虫媒传粉的进化模式是阐释壳斗科迁移和分化的关键.再者,壳斗科果实形态迥异,而包被果实的壳斗结构与雌花数目的关系成为果实特征的最佳体现[2].壳斗是无性枝融合后形成的包被果实的扁平状或碗装器官,壳斗类型的发生学不仅被广泛应用于壳斗科的分类,更帮助明晰各属之间的系统发育学关系.综上所述,传粉综合征、花序与壳斗的变异为壳斗科的起源,迁移和分化的各种假说提供证据,是分析壳斗科的系统发育和进化历程的关键.
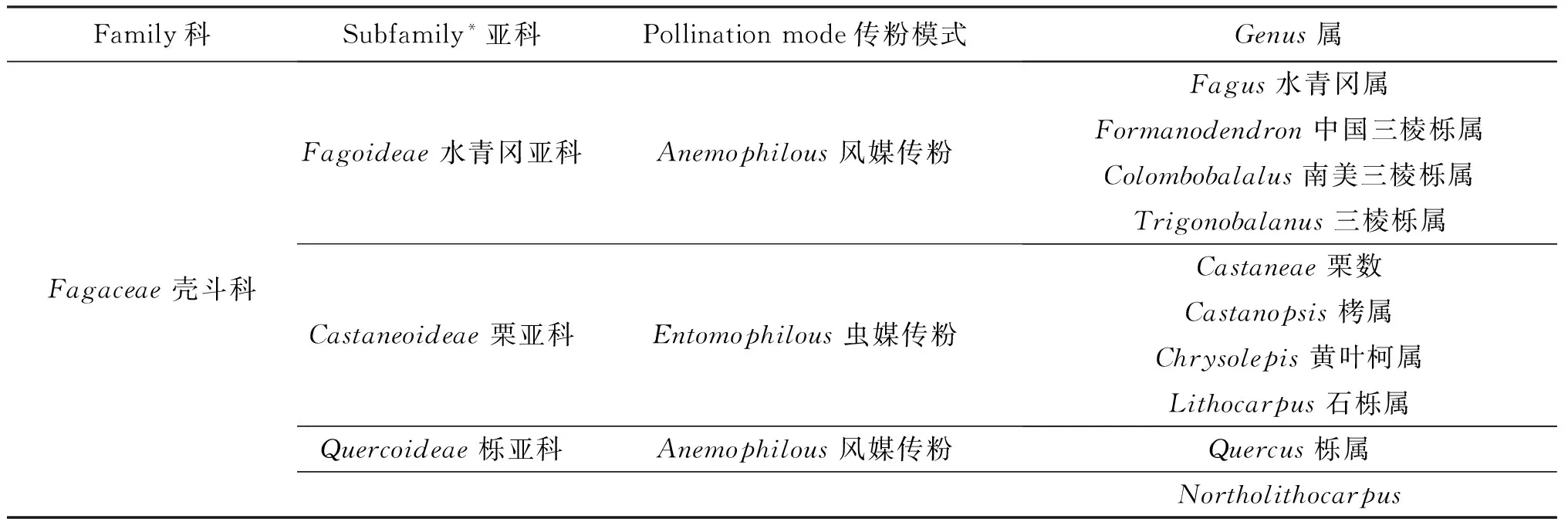
表1 壳斗科的分类现状
注:*现代系统发育学证据对于传统的亚科系统持有怀疑和保留.
3 传粉模式的演化
在以风媒传粉为主的庞大的壳斗目下,壳斗科的传粉综合征多态性,包括虫媒(entomophilous或insect pollination)和风媒(anemophilous或wind pollination)授粉铸就了该科的独特性.地质学的进化分析显示,被子植物各科从生物到非生物传粉模式的转变具有极度不对称性,且这种转化是以物种分化率的降低为代价的[3].这一理论在壳斗目的在进化枝上得到了很好的体现,除壳斗科外的其他风媒传粉科系的物种丰富度低下且雌雄同体.与这些科系相比,壳斗科的独特之处在于它同时具有2种传粉综合征和属内显著的物种分化程度.壳斗科的栗亚科的各属均为虫媒传粉,而水青冈亚科各属均为风媒传粉[4](见表 1).栎亚科的唯一一属栎属也为风媒传粉.令人吃惊的是,各种不同类型研究均显示壳斗科的原始传粉类型并非风媒授粉,而是通常被认为是更高级的虫媒传粉模式.例如,进化枝的分析显示,栗亚科虫媒传粉的花结构与壳斗科的原始花型类似[4].同样,白垩纪晚期的壳斗与雄花化石证据也显示当时的传粉方式为虫媒授粉[5-6].另外,被认为是水青冈亚科较原始的并系群的中国三棱栎属,南美三棱栎属,三棱栎属,也被证明是从虫媒到风媒传粉转变的过渡孑遗类群[4].系统发育学研究同样显示,风媒传粉作为新型传粉模式在壳斗科中至少进化了3次[2].
有研究指出,风媒传粉在壳斗科中的出现与进化是为了对应不同开花时期和不同昆虫活动的活跃程度而产生的.例如,在相同纬度下,壳斗科古热带区的各属,例如栲属、栗属和石栎属的开花期从早春一直持续到初夏,这种长时间的开花期与昆虫活动的高峰期相对应,因此维持虫媒传粉模式是适应频繁昆虫传粉活动的一种体现[7-8].而温带的栎属和水青冈属为了适应昆虫活动贫乏的春季开花期,进化出风媒传粉模式,成为对温带季节性气候适应性进化的成功案例[7].这种看似无法实现的从虫媒到风媒传粉的转变与壳斗科雌雄花序的分离分化密切相关[9].因此,为诠释从生物到非生物传粉转变的意义,必须在生态学背景下,对比化石记录与现代各属传粉模式,同时结合花序的功能进化,分析和对比2种传粉模式在壳斗科分化与进化中的作用.
4 花序的演化
壳斗科的花常单性,雌雄同株.雄花序花序主要是众多无柄或有柄的花朵聚集在一起形成葇夷或穗状花序.雌花单生或者2~3朵簇生于总苞内,总苞单生或者聚集在一起形成穗状花序.壳斗科花序充分展现了从虫媒授粉的完全花到风媒授粉的不完全花,以及两性到单性花的进化转变的各个阶段,是了解壳斗科进化与分化的重要研究材料.
壳斗科花序的演化从复杂到简单,包括失去无性与生殖的部分枝序,以及雄花与雌花的分离[10].无性与有性枝序的丧失与二歧聚伞花序的花朵数量减少和节间延伸的减少的2个进化特征相吻合.壳斗科祖先的花序可能类似于圆锥花序和聚伞圆锥花序.石栎属的患穗状花序聚集在一起形成更高级和更复杂的花序结构,被认为是壳斗科现存最复杂最原始的花序.栗属和栲属也有类似于石栎属的较原始的复杂花序结构[11].相对于这些复杂花序结构,栎属的花序最简单也最特化.栎属不仅实现了雌性葇荑花序与雄性穗状花序的完全分离,同时还具有确定数量的生殖枝.这种趋于简单化的花序进化被认为是复杂穗状花序由于生殖枝减少导致花序承载力下降的产物.花序的进化与传粉模式紧密对应:具有特化花序的栎属与先进的风媒传粉对应,而石栎属、栗属和栲属较原始的花序与他们传统的虫媒传粉模式相呼应[11].研究者认为具有风媒传粉模式的葇荑花序是从具有虫媒穿粉模式的穗状花序进化而来,并且随着海拔的上升花序复杂程度降低.这一假说与风媒与虫媒传粉类群的分布情况相吻合:虫媒传粉和复杂花序的类群如石栎属显著分布于低海拔的热带与亚热带地区;而风媒穿粉和简单化花序的类群如栎属与水青冈属广泛分布于更高海拔的温带常绿阔叶林中.然而,关于花序功能的其他证据十分缺乏,因验证花序与传粉模式的共同进化还需要从生态学的角度分析和对比化石证据和现存类群的联系.
5 壳斗的起源与演化
壳斗科最重要的形态学特征之一就是包裹果实之外的壳斗结构,壳斗结构丰富的多样性为壳斗科的分类提供了有力依据[2].虽然在其他科,如南水青冈科(Northofagaceae)[1],胡桃科(Juglandaceae)[3],金缕梅科(Hamamelidaceae)[3]中也具有壳斗或类似壳斗的结构,但是无论从起源或是系统发育来看,它们都与壳斗科差异较大,有着不同的起源[12].因此,壳斗的存在使壳斗科成为一个较自然的类群.作为壳斗科的共源性状,壳斗与花朵的特征决定了壳斗科各属之间的进化关联.
关于壳斗在壳斗科的起源问题,主要存在2种假说[4,13-15].一种理论认为壳斗是伴随壳斗科起源的产物[16];另一种理论认为壳斗是已有的植性结构或叶性结构,如苞片、托叶、无性小枝等的衍生物[13-18].众多研究结果支持第2种理论,尤其支持壳斗是从植性结构衍生而来的观点,认为壳斗是雌性花序的无性小枝融合后所形成的扁平状或碗装的结构[15-17].
目前主要有3种模式来阐释壳斗的演化途径:单一起源单向演化、单一起源双向演化和单一起源多向演化.单一起源单向演化提出时间最早,该理论认为水青冈属(Fagus)[1]和南水青冈属(Northofagus)[2]的裂瓣壳斗是最原始类型,在进化过程中裂瓣融合简化成其他类别的壳斗.该理论提出之时,三棱栎属(Trigonobalanus)[3]和黄叶柯属(Chrysolepis)[4]还没有被归纳到壳斗科之下,因此该理论未能相对完整地解释现代壳斗类型的关系.其次,后来人们发现南水青冈属与壳斗科并非单系,而是将其独立成南水青冈科(Northofagaceae).这两方面成为单一起源单向演化模式的重大缺陷.单一起源双向演化模式在加入了三棱栎属和黄叶柯属后由Forman提出[12,15].该理论假设的原始壳斗类型分别向两个方向演化,一个方向演化出石栎属(Lithocarpus)[5]与栗属(Castanea)[6]的壳斗;另一个方向演化出关键壳斗类型黄叶柯属壳斗后,进一步演化形成三棱栎属、栲属、水青冈属和南水青冈属的壳斗.该模式很好地解释了现代壳斗科至少5个属的壳斗形态特征演化关系,但因为把与壳斗科系统关系较远的南水青冈属包括在内而使其系统背景混乱,且该系统并未获得广泛的分子系统支持.单一起源多向演化假说是基于壳斗是二歧聚伞花序外部不育枝的衍生物这一理论,根据二歧聚伞花序分枝规律,即最外部分枝比内部分枝数多1的规律,提出壳斗分瓣数比雌花数多1的观点.该理论假设的原始壳斗类型一个方向朝着黄叶柯属演化,并且黄叶柯属壳斗保持了最多的原始特征,与假设原始壳斗类型有最直接的亲缘关系;另一个方向最初演化出模型中的关键类型,三角形果实的三棱栎属,然后分2个方向演化:一支形成水青冈属和栎属,另一支形成栲属和石栎属.3个壳斗演化模式的演化路径和原始壳斗类型各不相同,但最近的分子系统关系研究很大程度上支持了壳斗单一起源多向演化的模式,该模式也为其他研究者所认同[12].
在提出壳斗单一起源双向演化模式的同时,Forman提出了按2个方向演化的2种壳斗形态,二歧聚伞壳斗(dichasium cupule)和花朵壳斗(flower cupule)(见图1).二歧聚伞壳斗的雌花和其后来形成的果实被分瓣的壳斗结构所包被.这些分瓣呈三角形,在早期或开裂,或保持结合的形态对果实产生不同程度的包被,直至果实成熟时才开裂.而花朵壳斗的每朵雌花均为一个不分瓣的壳斗包被[1].传统认为花朵壳斗是壳斗科中的原始壳斗类型,在进化过程中通过与相邻花朵的壳斗结合,演变形成了二歧聚伞壳斗.而最近的进化枝分析显示二歧聚伞壳斗才是较原始的形态[4,16].这一结论不仅与壳斗科的壳斗化石证据相吻合,更得到发育生物学研究的支持,发现水青冈属和三棱栎属的二歧聚伞壳斗是壳斗科早期的分枝代表.
二歧聚伞壳斗是壳斗科中的主要壳斗类型,其壳斗的分瓣数与雌花数量成(n+1)/n的关系(见图1).例如,由三朵雌花组成的二歧聚伞花序的坚果被四个分瓣所包裹[4].二歧聚伞壳斗的衍征类型是水青冈属两花四瓣的壳斗[22,24].需要说明的是,一度与壳斗科并非姐妹类群的南水青冈科(Northofagaceae)被认为也具有也有二歧聚伞壳斗结构,后来发现其壳斗虽然形态与二歧聚伞壳斗类似,但其并非起源于二歧聚伞花序,而是起源于苞片外部的托叶[23-24].因此,二歧聚伞壳斗只在壳斗科中存在.

花朵壳斗仅在石栎属和栎属中出现.栎属与石栎属的壳斗都包裹着从单一雌花发育出的果实这一相似性造成了2个属在分类学上的混淆.同时,具有先进风媒传粉和特化花序的栎属,与保持传统虫媒传粉和复杂花序的石栎属同时具有花朵壳斗这一现象让人费解.对于这一矛盾,有学者提出栎属与石栎属的壳斗在起源上有本质的不同,栎属壳斗并非源于花朵壳斗,而是源于二歧聚伞壳斗,在进化过程失去了两侧的花朵,单一的中心花朵最终被融合了的壳斗结构包被.因此,栎属壳斗仅在形态上与石栎属的壳斗类似.另外,有证据显示2个属的壳斗在发生学上有着本质的不同.栎属的壳斗结构始于2个原基发展后的融合体,而石栎属的花朵壳斗始于一个单一的环状原基.对石栎属和栎属花朵壳斗的发生学进一步的研究可以明晰以及2个属之间的关系,验证2个属壳斗发生的异源性.
后来,Oh等提出第3种壳斗类型的概念[1],黄叶柯属的壳斗作为过渡类型介于花朵壳斗和二歧聚伞壳斗之间,所以其壳斗为二歧聚伞/花朵壳斗类型(Dichasium/flower cupule)(见图1).壳斗科壳斗的演化问题因黄叶柯属果实间的内生瓣(internal valves)结构而变得更加复杂.黄叶柯属壳斗的内生瓣结构被认为是复杂分枝体系的残留结构,与原始壳斗类型较为接近[19].但是各种分子系统研究不能统一3种壳斗类型的演化位置,而且单一起源多向演化假说不能很好地解释3种壳斗类型的发生和演化.因此除了系统发育证据,需要结合其他研究,如化石和形态分析来理解3种壳斗类型的进化顺序及关联.
6 结论
生殖特征如花序类型、传粉模式和壳斗类型的变异是探究壳斗科系统发育和进化的关键.分析各属的花序类型、穿粉综合征以及壳斗类型,发现壳斗科的进化是虫媒到风媒传粉,复杂到简单花序,以及二歧聚伞到花朵壳斗等一系列演化过程的结合和体现.其中,栎属具有最特化花序和新型风媒传粉模式,而石栎属具有非特化花序和原生的虫媒传粉模式.另外,虽然栎属和石栎属均为花朵壳斗,但是其发生学和起源可能存在本质的区别.
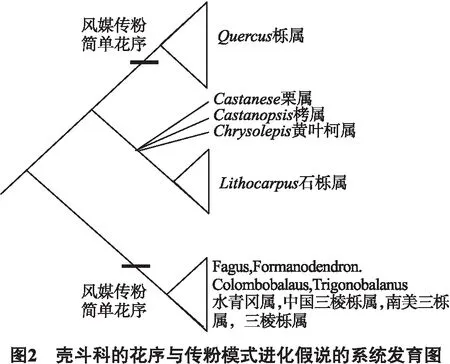
传粉综合征和花序的进化关联可以利用DNA的枝序分析结果[2]建立一个假说来解释.壳斗科的祖先生存于热带和亚热带地区的森林中.这些地区的充沛雨水可能冲洗掉大量的花粉,因此需要复杂花序承载大量花粉.同时,由于树冠层物种丰富度大,风媒传粉困难,因此壳斗科祖先以虫媒传粉和复杂穗状花序为特征.从渐新世早期到中新世,由于季节性和寒冷气候的转变,可能促进了部分壳斗科植物向温带的扩张和迁移.温带空旷的树冠层中空气流动性良好,风媒传粉在温带更有效并更具优势,因此迁移到温带的壳斗植物从虫媒到风媒传粉的演化从这里展开[26].同时,良好的空气流动保证了花粉的传播,花序承受力受到削减而演化成简单花序.其中,迁移到温带的水青冈亚科中的全部4个属都演变成风媒传粉和简单花序;迁移到温带的另一枝同样获得风媒传粉和简单花序性状,成为栎属(见图2).而留在热带亚热带地区的其余壳斗科植物,保持原有的虫媒传粉和复杂花序模式.它们在进化过程中分化成2枝,一枝成为栗亚科除石栎属外的三属,另一枝成为具有花朵壳斗结构[2](见图2).但是,拥有截然不同传粉模式和花序的栎属与石栎属,是在什么状况下进化出相同的花朵壳斗的仍不能得到很好的解释,需要进一步用发生学和系统发育学来了解.除此之外,3种壳斗类型的演化过程也需要进一步地探究.综上所述,壳斗科植物生殖特征,如花序,壳斗和传粉综合征等相互作用,共同影响了壳斗科的在迁移扩散和进化.因此综合地解析壳斗科各个特征的化石证据,结合生态学和生物地理学的证据是全面解析壳斗科的系统发育问题的关键.

[1] OH S-H,MANOS P S.Molecular phylogenetics and cupule evolution in Fagaceae as inferred from nuclear CRABS CLAW sequences[J].Taxon,2008,57(2):434-451.
[2] KAUL R B.Evolution and reproductive biology of inflorescences in Lithocarpus,Castanopsis,Catanea,and Quercus(fagaceae)[J].Ann Missouri Bot Gard,1986,73:284-296.
[3] DODD M E,SILVERTOWN J,CHASE M W.Phylogenetic analysis of trait evolution and species diversity variation among Angiosperm families[J].Evolution(N Y),1999,53(3):732-744.
[4] CREPET W L,NIXON K C.Earliest megafossil evidence of Fagaceae:phylogenetic and biogeographic implications[J].Am J Bot,1989,76(6):842-855.
[5] HERENDEEN P S,CRANE P R,DRINNAN A N.Fagaceous flowers,fruits,and cupules from the Campanian(Late Cretaceous)of central Georgia,USA[J].Int J Plant Sci,1995,156(1):93-116.
[6] SIMS H J,HERENDEEN P S,CRANE P R.New genus of fossil Fagaceae from the Santonian(late Cretaceous)of central Georgia,U.S.A.[J].Int J Plant Sci,1998,159(2):391-404.
[7] KAUL R B,ABBE E C,ABBE L B.Reproductive phenology of the oak family(Fagaceae)in the lowland rain forests of Borneo[J].Biotropica,1986,18(1):51-55.
[8] FOGDEN M P L.The seasonality and population dynamics of equatorial forest birds eat Sarawak[J].Ibis(Lond 1859),1972,114(3):307-343.
[9] BULL J,CHARNOV E.On irreversible evolution[J].Evolution(N Y),1985,39(5):1149-1155.
[10] KAUL R B,ABBE E C.Inflorescence architecture and evolution in the Fagaceae[J].J Arnold Arbor,1984,65:375-401.
[11] KAUL R B.Reproductive morphology of Quercus(Fagaceae)[J].Am J Bot,1985,72(12):1962-1977.
[12] CHEN Y,DENG M,ZHOU Z.A hypothesis on cupule evolution and the evidence from molecular phylogenies and fossils(in Chinese)[J].J Syst Evol,2008,46(1):41-52.
[13] BRETT D W.The inflorescence of Fagus and Castanea,and the evolution of the cupules of the Fagaceae[J].New Phytol,1964,63(1):96-118.
[14] ABBE E C.Flowers and inflorescences of the “Amentiferae”[J].Bot Rev,New York Botanical Garden,1974,40(2):160-256.
[15] FORMAN L L.On the evolution of the cupules in the Fagaceae[J].Kew Bull,1966,18(3):385-419.
[16] ENDRESS P K.Evolutionary trends in the Hamamelidales-Fagaceae-group[G]//KUBITZKI K.Flowering Plants.Vienna:Springer Vienna,1977:321-347.
[17] FEY B S,ENDRESS P K.Development and Morphological interpretation of the cupule in Fagaceae[J].Flora,1983,173:451-468.
[18] MACDONALD A D.Inception of the cupule of Quercus macrocarpa and Fagus grandifolia[J].Can J f Bot,1979,57:1777-1782.
[19] NIXON K C,CREPET W L.Trigonobalanus(Fagaceae):taxonomic status and phylogenetic relationships[J].Am J Bot,1989,76(6):828-841.
[20] LI J,SHU Q,ZHOU S,et al.Review and prospects of Quanternary Glaciation research in China(in Chinese)[J].J Glaciol Geocryol,2004,26(3):235-243.
[21] MANOS P S,ZHOU Z,CANNON C H.Systematics of Fagaceae?:phylogenetic tests of reproductive trait evolution[J].Int J Plant Sci,2001,162(6):1361-1379.
[22] OKAMOTO M.A comparative study of the ontogenetic development of the cupules in Castanea and Lithocarpus(Fagaceae)[J].Plant Syst Evol,1989,168(1-2):7-18.
[23] ROZEFELDS A c.,DRINNAN A N.Pistillate inflorescence architecture and morphology of the southern beeches[J].Am J Bot,1997,84(6):54.
[24] AXELORD D I.Biogeography of oaks in the Arcto-Tertiary province[J].Ann Missouri Bot Gard,1983,70(4):629-657.
(责任编辑 王 琳)
Evolution of pollination modes, inflorescence and cupule features of Fagaceae
CHEN Xi
(School of Ecology and Environment, Yunnan University,Kunming 650091,China)
Ten genera and over 1 000 species of Fagaceae are widely distributed in the northern hemisphere. The studies on the evolution of Fagaceae have focused on three main research aspects: the evolution of pollination modes, inflorescence and cupule. These three aspects are closely related to the migration and distribution of Fagaceae. So far, simplified inflorescence, anemophilous pollination and flower cupule are identified as apomorphies in the evolution of Fagaceae. However,whyQuercusandLithocarpus,which share the advanced flower cupule with extremely different inflorescences and pollination modes, remain unexplained. Therefore, combining fossil evidences, morhoplogical and phylogenetic is crucial to clarify the evolution connections among pollination modes, inflorescence and cupule of Fagaceae.
Fagaceae;evolution;pollination syndromes;inflorescence;cupule
2014-06-23.
国家自然科学基金(31460095).
陈曦(1983-),女,博士,讲师.主要研究方向:石栎属植物的生态与进化.
Q941
A
1672-8513(2015)01-0019-06
