Na2CO3胁迫对藜渗透调节物质和保护酶活性的影响
2015-06-15丁楚楚黄鑫金彗莉李雪华胡玉龙赵苹艺孙存华
丁楚楚+黄鑫+金彗莉+李雪华+胡玉龙+赵苹艺+孙存华


摘要:探讨不同浓度Na2CO3胁迫对藜叶片中渗透调节物质和抗氧化酶活性的影响,用0.15%、0.30%、0.45% Na2CO3溶液对藜幼苗进行胁迫处理,测定相关生理参数的变化。结果表明:随着胁迫程度的增加和胁迫时间的延长,藜幼苗生理特性发生明显变化:渗透调节物质(可溶性糖、脯氨酸、可溶性蛋白)与丙二醛含量在一定范围内显著提高;保护酶(超氧化物歧化酶、过氧化物酶、过氧化氢酶)活性随胁迫时间的延长呈先升高后降低的趋势。综合比较可知,高浓度、长时间的碱性盐胁迫会对藜造成一定的损伤。
关键词:藜;Na2CO3碱性盐胁迫;渗透调节物质;保护酶
中图分类号: Q945.78 文献标志码: A
文章编号:1002-1302(2015)04-0338-03
收稿日期:2014-05-29
基金项目:国家科技重大专项(编号:2014ZX08012-002、2011ZX08012-002);江苏省优势学科建设工程;江苏省徐州市自然科学基金(编号:XF13C056)。
作者简介:丁楚楚( 1993—),女,江苏徐州人,本科生,从事植物生理及分子生物学研究。E-mail:dingchuchu9393@163.com。
通信作者:孙存华,教授,硕士生导师,从事植物生理生态及分子生物学研究。E-mail:chsun@jsnu.edu.cn。
随着全球环境的不断恶化,土壤盐碱化日益加重,威胁着人类赖以生存的土壤资源,是制约农业发展的主要环境问题。近年来,有关植物抗盐碱机制方面的研究不断深入,但多以NaCl胁迫为主要研究对象[1-2]。实际上,在我国大面积的盐碱地中,有不少为复合型盐碱地,即存在着盐化和碱化相伴发生的情况[3]。颜宏等曾报道,中性盐胁迫、碱性盐胁迫是2种不同性质的胁迫,既有相关性又有明显的区别[4]。有研究发现,碳酸盐的累积使土壤的pH值升高,所造成的土壤碱化问题比NaCl等中性盐更加严重[5]。因此,碱胁迫也是生产实践中存在的主要问题,而目前关于碱性盐胁迫对植物生理特性影响的研究相对较少[4-5]。
藜(Chenopodium album L.)为藜科藜属的1年生草本植物,其叶和嫩茎可食用,种子可榨油,能止泻痢、止痒、止疼,具有较高的营养价值和药用价值。藜耐旱性较强,适应于干旱地区,在盐碱地中也能正常生长[6]。目前有关藜的抗盐碱研究报道不多[7-8],有关藜在碱性盐胁迫下的渗透调节物质和保护酶活性的研究未见报道。因此,本研究以藜为材料,通过人工控制Na2CO3浓度来研究藜对碱性盐胁迫的生理生化反应,以探讨其耐碱性盐的能力,从而为盐碱地区藜的人工栽培提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为藜,选饱满大小一致的种子,用0.1% HgCl2消毒后播种在装有石英砂的塑料盒中,用Hoagland溶液在阳光充足的条件下培养。待藜苗长到6叶期时,用含0.15%、0.30%、0.45% Na2CO3的Hoagland溶液进行根部胁迫处理,同时以不含 Na2CO3的溶液作为对照。分别在处理后0、1、3、5、7 d,对植株上部第3~5对功能叶片进行各项生理指标的测定,每项指标的测定设3个重复。
1.2 测定方法
可溶性糖的鲜质量含量的测定采用蒽酮比色法[9];脯氨酸的鲜质量含量的测定用采用酸性茚三酮显色法[9];丙二醛(MDA)的鲜质量含量的测定采用硫代巴比妥酸显色法[9];可溶性蛋白的鲜质量含量的测定用采用紫外吸收法[10];超氧化物歧化酶(SOD)活性的测定采用南京建成生物工程研究所生产的SOD试剂盒;过氧化物酶(POD)活性的测定采用愈创木酚法[10];过氧化氢酶(CAT)活性的测定参照Jablonski等的方法[11]。
1.3 数据处理
应用Excel作图并进行数据处理。
2 结果与分析
2.1 Na2CO3碱性盐胁迫对藜幼苗渗透调节物质的影响
2.1.1 Na2CO3碱性盐胁迫对藜幼苗可溶性糖含量的影响 从图1可以看出,各处理浓度植物体内的可溶性糖含量随着处理时间的延长和处理溶液浓度的增加大体呈不断升高的趋势。在前3 d,各处理浓度植物体内可溶性糖的含量与对照差异不明显;5 d后,各处理浓度可溶性糖的含量急剧升高,部分处理间差异显著(P<0.05);在处理7 d后,0.15%、0.30%、0.45% Na2CO3浓度处理下藜幼苗可溶性糖的含量分别是对照的3.09、507、7.18倍。结果表明,可溶性糖是藜在碱胁迫下的重要渗透调节物质。
2.1.2 Na2CO3碱性盐胁迫对藜幼苗脯氨酸含量的影响 渗透胁迫能诱导植物体内脯氨酸大量积累,它是植物体内重要的渗透调节物质,对于提高植物盐碱胁迫耐受性具有重要作用[12]。从图2可以看出,在Na2CO3胁迫下,藜幼苗叶片内脯氨酸含量大量积累,处理后1 d,各浓度Na2CO3碱胁迫下的藜幼苗叶片内脯氨酸含量与对照差异不明显;随着Na2CO3胁迫浓度的增加和处理时间的延长, 藜幼苗叶片内脯氨酸的
积累量急剧增加,尤其是0.45% Na2CO3胁迫处理3 d,藜幼苗叶片内脯氨酸含量与对照相比显著增加(P<0.05),在处理5 d达到最大值,是对照的6.97倍。这些结果表明,藜幼苗叶片内的游离脯氨酸是藜的重要渗透调节物质。
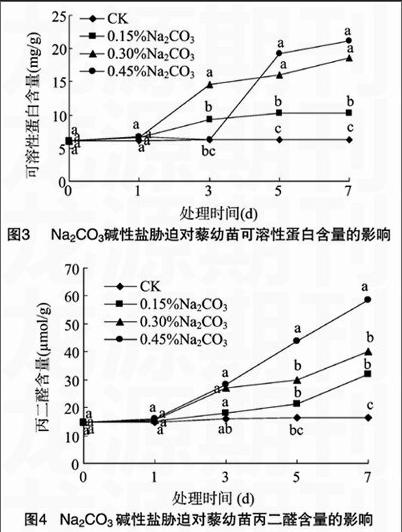
2.1.3 Na2CO3碱性盐胁迫对藜幼苗可溶性蛋白含量的影响 余叔文等的研究表明,可溶性蛋白与调节植物细胞的渗透势有关,高含量的可溶性蛋白可帮助维持植物细胞较低的渗透势,抵抗逆境产生的胁迫[13]。从图3可以看出,处理开始时,各浓度Na2CO3碱性盐胁迫下藜幼苗叶片内可溶性蛋白含量与对照差异不大;处理1 d后,各处理组藜幼苗叶片内可溶性蛋白含量呈上升趋势;处理5 d和7 d,可溶性蛋白的含量明显增加,0.45%Na2CO3处理浓度下,可溶性蛋白含量从处理后3 d起急剧增加,到处理7 d,各处理组可溶性蛋白含量高于对照,0.15%、0.30%、0.45% Na2CO3处理浓度下可溶性蛋白含量分别是对照的1.63、2.96、3.37倍。结果表明,可溶性蛋白是参与藜在碱胁迫下抗逆反应的重要渗透调节物质。endprint
2.2 Na2CO3碱性盐胁迫对藜幼苗膜脂过氧化作用和保护酶的影响
2.2.1 Na2CO3碱性盐胁迫对藜幼苗丙二醛含量的影响 丙二醛是膜脂过氧化作用的产物,也是反映膜脂过氧化导致膜损伤的主要标志[14]。从图4可以看出,在处理1 d后,各处理组丙二醛含量增幅不明显;从处理3 d开始,丙二醛含量增幅明显,特别是0.45%Na2CO3胁迫下,丙二醛含量急剧上升;处理7 d后,各处理浓度的植物体内的丙二醛含量分别是对照的1.94、2.47、3.57倍,部分处理间差异显著(P<005)。总体看出,各处理浓度下藜幼苗的丙二醛含量均有所增加且比对照高,这说明高浓度及长时间的Na2CO3胁迫会使藜幼苗的细胞膜受到严重伤害。
2.2.2 Na2CO3碱性盐胁迫对藜幼苗SOD活性的影响 SOD活性的高低是植物抗逆性强弱的重要标志之一,SOD可以将超氧阴离子自由基歧化成H2O2,是抗氧化伤害的关键酶之一。从图5可以看出,在处理1 d后,0.45%Na2CO3胁迫下藜幼苗SOD活性达到最大值,是对照的1.33倍,随着处理时间的延续,活性下降;0.15%、0.30% Na2CO3胁迫下藜幼苗的SOD活性与对照相比差异不大。
2.2.3 Na2CO3碱性盐胁迫对藜幼苗POD活性的影响 从图6可以看出,各浓度Na2CO3胁迫下,藜幼苗POD活性是先上升后下降,但都比对照高。在处理3 d后,0.30%、0.45% Na2CO3胁迫下,藜幼苗POD活性达到最大值,分别是对照的4.37、4.59倍(P<0.05);0.15% Na2CO3胁迫下,藜幼苗POD活性在处理5 d达到最大值,是对照的2.15倍,且差异显著(P<005)。
2.2.4 Na2CO3碱性盐胁迫对藜幼苗CAT活性的影响 CAT是抗氧化酶系统的重要组成成分,与SOD、POD协同作用防御活性氧及其他过氧化物自由基对细胞膜系统所造成的伤害,从而提高植物的抗逆性[15]。从图7可以看出,随胁迫时间的延长,不同浓度Na2CO3胁迫下的CAT活性变化很大。处理
1 d后,CAT的活性和对照相比增加不显著;处理3 d后,各处理组的藜幼苗CAT的活性急剧增长;处理5 d后,各处理浓度达到最大值,0.15%、0.30%、0.45% Na2CO3胁迫下藜幼苗CAT的活性分别是对照的2.89、3.51、4.19倍(P<0.05);处理7 d后,各处理浓度下CAT的活性又急剧下降,但总体比对照高。
3 结论与讨论
渗透调节是植物适应盐碱胁迫的一种重要的生理机制,在渗透胁迫下,植物会通过主动积累有机溶质,如可溶性糖、游离氨基酸(特别是脯氨酸)、可溶性蛋白等来降低渗透势和水势,适应环境带来的变化,维持正常的代谢活动[16]。本研究中,藜幼苗中可溶性糖、脯氨酸、可溶性蛋白的含量都随碱性盐浓度增加而增加,这与前人以芨芨草[17]、青山杨[18]等植物为试验材料,在Na2CO3胁迫下的研究结果相一致。说明可溶性糖、脯氨酸和可溶性蛋白是植物体内重要的渗透调节物质,而且藜具有一定的渗透调节能力,对盐碱环境具有一定的适应性。
SOD、POD、CAT 都是细胞膜系统的保护酶,当植物处于逆境时,SOD能将超氧阴离子自由基歧化成H2O2,而POD、CAT可将H2O2变为H2O,3种酶协调作用可使活性氧维持于一个较低水平,减弱膜脂过氧化作用对细胞膜的伤害,对于保持植株体内的代谢平衡起着重要作用[5]。在本研究中,SOD、POD、CAT这3种酶的活性基本上呈先上升后下降的趋势,说明在碱胁迫下,保护酶可以缓解膜脂过氧化伤害;但随着胁迫时间的延长,这3种保护酶的活性有所下降,表明胁迫时间太长会超过藜自身的耐受程度,使膜脂过氧化严重,植株受到损害,导致酶活性下降。
处理结束时,0.45%浓度碱胁迫下的藜幼苗因胁迫强度过高导致全部死亡;几天后,0.30%浓度碱胁迫下的藜幼苗叶片变红也逐渐死亡;0.15%浓度的碱胁迫苗虽然能存活一段时间,但是生长缓慢不旺盛,说明高浓度、长时间的碱性盐胁迫会对藜造成严重损伤。
综上所述,在Na2CO3碱性盐胁迫下,藜幼苗叶片通过增加渗透调节物质的含量来降低渗透势和水势,通过增强保护酶的活性来减弱膜脂过氧化作用。说明藜在渗透调节能力以及膜保护体系方面对碱胁迫具有一定的适应性,但是高浓度、长时间的碱胁迫会对藜造成一定的损伤。
参考文献:
[1]高武军,于美玲,邓传良,等. NaCl胁迫对6种红花幼苗渗透调节物质及抗氧化系统的影响[J]. 武汉植物学研究,2010,28(5):612-617.
[2]李 聪,郭梦阳,韩烈保. NaCl胁迫对烟草种子萌发及幼苗生理特性的影响[J]. 中国农学通报,2013,29(3):103-107.
[3]李 彬,王志春,孙志高,等. 中国盐碱地资源与可持续利用研究[J]. 干旱地区农业研究,2005,23(2):154-158.
[4]颜 宏,赵 伟,盛艳敏,等. 碱胁迫对羊草和向日葵的影响[J]. 应用生态学报,2005,16(8):1497-1501.
[5]武 德,曹帮华,郑黎明,等. 碱式盐胁迫对绒毛白蜡保护酶系统的影响[J]. 安徽农学通报,2007,13(9):61-62.
[6]Sun C H,Li Y,He H Y,et al.Physiological and biochemical responses of Chenopodium album to drought stresses[J]. Acta Ecologica Sinica,2005,25(10):2556-2561.
[7]油天钰. 藜科植物藜在盐胁迫下的形态结构及其对盐的生理响应[D]. 乌鲁木齐:新疆大学,2010.
[8]施剑杰,金慧丽,孙存华,等. NaCl胁迫对藜幼苗叶片渗透调节物质和保护酶的影响[J]. 安徽农业科学,2011,39(12):6943-6946.
[9]李合生,孙 群,赵世杰,等. 植物生理生化实验原理与技术[M]. 北京:高等教育出版社,2000:184-195.
[10]张志良,瞿伟菁. 植物生理学实验指导[M]. 3版. 北京:高等教育出版社,2003:123-124.
[11]Jablonski P P,Anderson J W.Light-dependent reduction of dehydroascorbate by ruptured pea chloroplasts[J].Plant Physiol,1981,67(6):1239-1244.
[12]赵福庚,刘友良. 胁迫条件下高等植物体内脯氨酸代谢及调节的研究进展[J]. 植物学通报,1999,16(5):540-546.
[13]余叔文,汤章城. 植物生理和分子生物学[M]. 北京:科学出版社,1999:739-745.
[14]汪月霞,孙国荣,王建波,等. NaCl胁迫下星星草幼苗MDA含量与膜透性及叶绿素荧光参数之间的关系[J]. 生态学报,2006,26(1):122-129.
[15]胡云云,杨 洁,张富春. NaCl胁迫对灰绿藜中保护酶活性及MDA含量的影响[J]. 生物技术,2010,20(4):52-54.
[16]梁新华,刘凤敏. NaCl和Na2CO3胁迫对甘草幼苗渗透调节物质含量的影响[J]. 农业科学研究,2006,27(2):96-98.
[17]董秋丽,夏方山,董宽虎. 碱性盐胁迫对芨芨草苗期脯氨酸和可溶性蛋白含量的影响[J]. 畜牧与饲料科学,2010,31(4):11-12,28.
[18]闫永庆,王文杰,朱 虹,等. 混合盐碱胁迫对青山杨渗透调节物质及活性氧代谢的影响[J]. 应用生态学报,2009,20(9):2085-2091.endprint
