中国部分鸽mtDNA D—loop区遗传多态性与系统进化分析
2015-06-15付胜勇刘宏祥谢鹏童海兵徐善金陈
付胜勇+刘宏祥+谢鹏+童海兵+徐善金+陈卫彬+卜柱


摘要:利用PCR测序及生物信息学分析技术,测定中国境内8个品种(系)鸽108份样本mtDNA D-loop区部分序列,研究中国部分肉鸽品种(系)的遗传多态性与系统进化关系。结果表明,在扩增的761 bp mtDNA D-loop区序列间发现3个变异位点,约占分析位点总数的0.39,具4个单倍型;8个群体内单倍型多样度为0.333~0.867,总体单倍型变异度为0.559,总体核苷酸多样度为0.000 85,表现出较为贫瘠的遗传多态性,未表现出显著的遗传分化;石岐鸽与欧洲肉鸽Ⅰ、银羽王鸽与欧洲肉鸽Ⅰ、泰深鸽与银羽王鸽间呈现出较大的遗传距离,在后期配套系培育及杂交优势利用方面具较大遗传选育空间。
关键词:鸽;mtDNA D-loop;遗传多态性;系统进化
中图分类号: S836.2 文献标志码: A
文章编号:1002-1302(2015)04-0041-03
收稿日期:2014-05-16
基金项目:江苏省农业科技自主创新资金[编号:CX(12)3073];江苏省扬州市农业科技攻关(编号:yz2011068);江苏省苏北科技发展计划(编号:BN2013043)。
作者简介:付胜勇(1985—),男,四川内江人,硕士,助理研究员,从事家禽营养与品种选育研究。E-mail:xnfsy@163.com。
通信作者:卜 柱,硕士,副研究员,从事肉鸽生产与装备创新研究。E-mail:jsbuzhu@163.com。
高等动物的线粒体DNA(mtDNA)为共价闭合的环状双链DNA分子,大小为16 355~16 359 bp,其结构简单稳定,由37个基因和1段D-loop区(控制区)组成,符合母系遗传,不受引进品种杂交改良的影响,可以真实地反映当地物种起源[1]。哺乳动物及鸟类在mtDNA中进化速度最快、多态性最丰富,在动物起源演化、遗传距离测算生物地理学、系统发育等方面得到广泛的应用[2]。从已公布的序列来看,鸽线粒体DNA由17 229对碱基构成[3],分布有13个基因位点,其中,第15 573~17 229 bp序列为D-loop区,全长1 656 bp。本研究对中国鸽mtDNA D-loop区多态性及系统进化进行取样分析研究,以推进我国肉鸽的系统选育工作。
1 材料与方法
1.1 材料
石岐鸽和银羽王鸽血样各12份,采自广东省中山市石岐鸽场;2002年引自法国的欧洲肉鸽Ⅰ、Ⅱ、Ⅲ系血样各12份、源自美国的白羽王鸽和银羽王鸽血样各12份、源自深圳天翔达祖代种鸽场的卡奴鸽和泰深鸽血样各12份,均采自江苏省江阴威特凯鸽业有限公司。每群体取成年公鸽和母鸽各6羽,共计8个品种(系)108份样本,采用柠檬酸+柠檬酸钠+葡萄糖(ACD)抗凝静脉采血1 mL,加入裂解液,4 ℃保存;采用Guo等的方法[4]提取基因组DNA。
1.2 引物设计
从NCBI(http://www.ncbi.nlm.nih.gov/nuccore/NC_013978)下载鸽子线粒体全序列(登录号:NC_013978.1)为参考序列,用Primer 5.0软件设计2对引物,扩增片段覆盖D-loop区。引物信息为:F1:5′-TTCCGCTTGGCCTTTCTC-3′,R1:5′-GATGACCGCAATGCCGAA-3′;F2:5′-CTCGACCTCAGGTACTACTGGCGT-3′,R2:5′-ACTTCTGCGGTTCTGCCGGGATGT-3′。
1.3 线粒体DNA D-loop区扩增与序列测定
PCR扩增体系的反应程序:95 ℃预变性5 min;94 ℃变性30 s,55 ℃ 退火60 s,72 ℃延伸30 s,38个循环;72 ℃充分延伸 7 min,4 ℃保存。PCR产物用2.0%琼脂糖(美国FMC)凝胶电泳检测其片段大小、纯度、亮度,小量胶回收试剂盒回收目的片段;分别取2.5~5.0 μL回收产物,用PCR引物进行测序反应,测序试剂采用Thermo Sequenase Cycle Squencing Kit(美国USB公司),按照操作规程进行;委托上海英俊生物测序公司使用LI-COR 4200 DNA自动分析仪(LI-COR Biotechnology Division,Lincoln,NE)进行序列分析。
1.4 数据处理
扩增所得的mtDNA D-loop区序列用DNA MAN进行序列编辑;用Mega软件进行同源序列比对,DNA SP5.10软件统计不同鸽品种(系)mtDNA D-loop区的核苷酸多态性位点数(number of polymorphie sites)、总突变位点数(total number of mutations)、单倍型数(number of Haplotypes)、单倍型多样度(haplotype diversity)、核苷酸多样性(nueleotide diversity)、平均核苷酸差异(average number of nucleotide differences)、遗传距离等;采用邻接法(Neighbor-Jointing,NJ)最大似然法、最大简约法构建分子系统发育树,其中节点自举置信水平采用Bootstrap估计和1 000次循环;用Network软件分析单倍型演变及分布情况。
2 结果与分析
2.1 不同鸽品种(系)mtDNA D-loop区单倍型与核苷酸变异
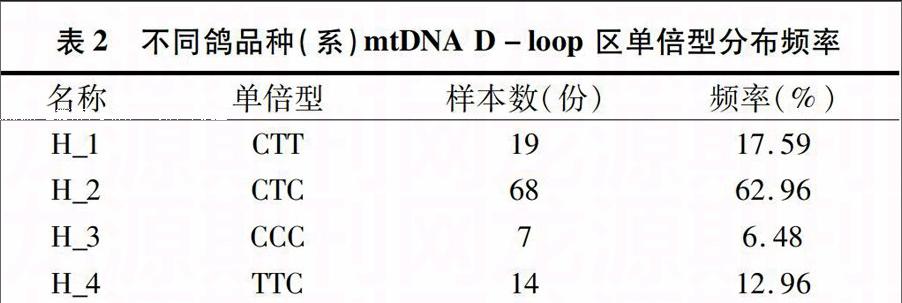
本研究扩增并测定肉鸽部分的D-loop 序列,靠近tRNAPro基因一侧扩增片段总长为761 bp。由表1可见,8个群体单倍型多样度为0.333~0.867,总体单倍型变异度为0559,总体核苷酸多样度为0.000 850;利用Mega分析软件对108条mtRNA D-loop区进行同源序列比对,在8个鸽品种(系)中共发现3个多态位点,分别位于第50、第51、第148碱基处;mtRNA D-loop区序列段由4种单倍体型组成。由图1和表2可见,单倍型分布频率以CTC型所占比例最大,为 62.96%,单倍型总数和多态位点总数以中山石歧鸽和欧洲肉鸽Ⅲ最丰富,涵盖有全部多态性位点和单倍型。
表1 不同鸽品种(系)mtDNA D-loop 区单倍型与核苷酸变异
品种(系) 样本数(份) 多态位点数(个) 单倍型数 单倍型多样度 核酸多样度
石岐鸽 12 3(50 bp,51 bp,148 bp) 4 0.525 0.000 770
银羽王鸽 24 2(50 bp,51 bp) 3 0.386 0.000 530
欧洲肉鸽Ⅰ 12 1(148 bp) 2 0.667 0.000 880
欧洲肉鸽Ⅱ 12 1(51 bp) 2 0.500 0.000 660
欧洲肉鸽Ⅲ 12 3(50 bp,51 bp,148 bp) 4 0.867 0.000 158
卡奴鸽 12 2(50 bp,148 bp) 3 0.733 0.000 114
白羽王鸽 12 1(148 bp) 2 0.333 0.000 440
泰深鸽 12 1(148 bp) 2 0.533 0.000 700
总和 108 3 4 0.559 0.000 850
2.2 不同鸽品(系)种mtDNA D-loop 区序列系统发育进化树构建
由图2可见,本研究的鸽品种(系)mtDNA D-loop序列
表2 不同鸽品种(系)mtDNA D-loop 区单倍型分布频率
名称 单倍型 样本数(份) 频率(%)
H_1 CTT 19 17.59
H_2 CTC 68 62.96
H_3 CCC 7 6.48
H_4 TTC 14 12.96
大致聚为2个聚类簇,欧洲肉鸽Ⅰ、Ⅲ和泰深鸽可分为一大类,卡奴鸽、石岐鸽、银羽王鸽、白羽王鸽等品种(系)可分为另一大类,在系统进化上显现出品种(系)间的血缘关系远近和亲疏讯息。
2.3 不同鸽品种(系)间遗传距离
由表3可见,我国现存不同鸽品种(系)间遗传距离变异范围为0.000 7~0.005 1;石岐鸽与欧洲肉鸽Ⅰ、银羽王鸽与欧洲肉鸽Ⅰ、泰深鸽与银羽王鸽间距离值较大,分别为 0.004 7、0.005 1、0.004 4;其他品种(系)间距离值相对较小,变异范围为0.000 7~0.004 1。
表3 不同鸽品种(系)间kimura双参数遗传距离
品种(系)
遗传距离
SQ YY OⅠ OⅡ OⅢ KN TS BY
SQ — 0.001 6 0.004 7 0.002 0 0.003 4 0.002 3 0.004 1 0.002 3
YY 0.001 6 — 0.005 1 0.001 8 0.003 4 0.002 2 0.004 4 0.002 2
OⅠ 0.004 7 0.005 1 — 0.003 3 0.002 9 0.002 9 0.000 7 0.002 9
OⅡ 0.002 0 0.001 8 0.003 3 — 0.002 9 0.001 8 0.002 9 0.001 8
OⅢ 0.003 4 0.003 4 0.002 9 0.002 9 — 0.002 9 0.002 9 0.002 9
KN 0.002 3 0.002 2 0.002 9 0.001 8 0.002 9 — 0.002 7 0.001 9
TS 0.004 1 0.004 4 0.000 7 0.002 9 0.002 9 0.002 7 — 0.002 7
BY 0.002 3 0.002 2 0.002 9 0.001 8 0.002 9 0.0019 0.002 7 —
注:SQ、YY、OⅠ、OⅡ、OⅢ、KN、TS、BY分别为石岐鸽、银羽王鸽、欧洲肉鸽Ⅰ、欧洲肉鸽Ⅱ、欧洲肉鸽Ⅲ、卡奴鸽、泰深鸽、白羽王鸽。
3 结论与讨论
本试验测定了8个鸽品种(系)间共108个mtDNA D-loop序列,发现有3个变异位点,分别位于第50、第51、第148碱基,占分析总数的0.39%,所有多态位点均为两核苷酸变异;8个鸽品种(系)间检测到4种单倍型,单倍型多样度为0.559,核苷酸多样度为0.000 850,多样性相对较为匮乏,这与胡永胜等研究结论[5]不一致,可能是因为选择的序列区域与比对范围不同。相对于驴、牦牛、奶山羊、鸡等家畜禽[2,6-7]动辄成十上百的变异位点而言,我国现有鸽品种(系)显现出较为不丰富的遗传多态性,这可能与鸽专门化品系选育工作开展较晚及鸟类特有的飞翔迁移流动性较大有关。品种(系)选育导致品种间有差异变化的基因突变不能有效留存下来;鸟类特有的飞翔迁移流动性导致群体间不能形成有效的环境、气候、生态样生殖隔离,进而闭锁繁衍出新的类群或亚型,引起自然地域隔离造成生态多样性减少[8]。系统进化分析结果表明,本研究中的鸽种群均可追溯于同一母系起源,我国现存鸽品种(系)大致分为2类,从欧洲引进的欧洲肉鸽和泰深鸽为1类,从美国引进的白羽王鸽、银羽王鸽及地方品种石岐鸽等为另一大类。
石岐鸽是由广州中山海外侨胞带回的优良种鸽与中山本地优良鸽品种进行杂交培育而成[8],距今已有100年历史。本试验结果表明,石岐鸽与银羽王鸽间遗传距离较小,为0001 6,在系统发育进化树中也与银羽王鸽同支相连,这说明在石岐鸽品种形成过程中,可能引入了银羽王鸽血统。卡奴鸽在美国棕榈鸽场1915年开始选育,1932年正式命名,先后导入卡奴鸽与白色贺姆鸽、白羽王鸽和白色仑替鸽等血脉[9],卡奴鸽与其他各品种(系)间遗传距离差异较小,主要与其育成过程有关。
泰深鸽、欧洲肉鸽属于欧洲地域品种,而白羽王鸽、卡奴鸽、银羽王鸽属于北美地域品种,国内的石岐鸽兼有北美血统和亚洲血统[9],本研究中遗传距离参数测算分析的结果与之较为吻合,即相同区域内的鸽品种(系)间遗传距离较近,不同区域间鸽品种遗传距离较远。遗传距离测算分析表明,石岐鸽与欧洲肉鸽Ⅰ、银羽王鸽与欧洲肉鸽Ⅰ、泰深鸽与银羽王鸽间遗传距离较远,在后期杂交优势利用、配套系培育方面有较大的遗传选育空间。
参考文献:
[1]Liu R Y,Yang G S,Lei C Z. The genetic diversity of mtDNA D-loop and the origin of Chinese goats[J]. Acta Genetica Sinica,2006,33(5):420-428.
[2]雷初朝,陈 宏,杨公社,等. 中国驴种线粒体DNA D-loop多态性研究[J]. 遗传学报,2005,32(5):481-486.
[3]赖松家,王 玲,刘益平,等. 中国部分牦牛品种线粒体DNA遗传多态性研究[J]. 遗传学报,2005,32(5):463-470.
[4]Guo S,Savolainen P,Su J,et al. Origin of mitochondrial DNA diversity of domestic yaks[J]. BMC Evolutionary Biology,2006(6):73.
[5]胡永胜,方梅霞,何丹林,等. 鸽mtDNA遗传多样性及系统进化研究[J]. 中国家禽,2010,32(14):23-26.
[6]李 辉,吴 婵,熊光源,等. 黔东南小香鸡Cyt b基因遗传多态性及系统进化研究[J]. 江苏农业科学,2013,41(1):21-22.
[7]皮秀霜. 中国奶山羊线粒体DNA Dloop区遗传多样性与起源进化研究[D]. 泰安:山东农业大学,2013.
[8]张学余,苏一军,李国辉,等. 11个鸡品种产蛋性能与产区生态的聚类分析[J]. 家畜生态学报,2012,33(2):50-54.
[9]卜 柱,厉宝林,赵振华,等. 中国肉鸽主要品种资源与育种现状[J]. 中国畜牧兽医,2010,37(6):116-119.
[10]卜 柱,赵宝华.图说高效养肉鸽关键技术[M]. 北京:金盾出版社,2012.
