龙脑樟叶片光合特性对氮磷钾肥的响应
2015-05-28陈周琴李燕山彭招兰曹福亮郁万文
陈周琴,张 露*,李燕山,苏 恒,彭招兰,曹福亮,郁万文,杨 亮
(1.江西农业大学 林学院,江西 南昌 330045;2.吉安市林业科学研究所,江西 吉安 343101;3.南京林业大学 林学院,江苏 南京 210037)
龙脑樟(Cinnamomum camphora chvar.Borneol)是樟科樟属樟树 的5个化学类型之一 ,其鲜叶挥发油富含右旋龙脑(天然冰片)。目前,国内外对天然冰片的需求量远高于现有的生产能力,它的价格一直飚升,龙脑樟产业具有较大的发展前景[4]。因此龙脑樟的栽培技术和合理的开发利用越来越受到重视,目前已在种苗繁育[5-6]、组织培养[7-8]、精油成分分析[9-10]、龙脑含量[12]和医药应用[13-15]等方面开展了大量研究,其中施肥能够显著提高龙脑樟矮林叶片生物量得到证实[16],但其相关的一些生理机制,尤其是施肥对叶片光合特性的影响机制还不清楚。
近几年,有不少的研究者开展了许多有关氮磷钾施肥对植物光合作用影响的试验并取得了一定的成果,舒翔等分别对香樟(Cinnamomum camphora(L.)Presl)幼苗[17]、毛白杨(Populus tomentosa Carr)幼苗[18]、桑树[19](Morus L.)和文冠果[20](Xanthoceras sorbifolia Bunge)进行了氮磷钾配施处理的试验,结果表明氮、磷、钾肥的配施对其光合作用有促进作用,并且存在显著的差异,并且不同水平氮磷钾配比施肥处理对植物的叶绿素含量和酶活性等均有影响,从而对其光合作用以及生长发育产生不同的影响。
本试验采用“3414”不完全最优回归的设计方案,该设计具有处理少、效率高的特点,被广泛应用于国内外的肥料效应试验研究中。本文研究了氮磷钾不同的施肥处理对龙脑樟叶片光合特性的影响,旨在探讨不同氮磷钾施肥处理对龙脑樟光合作用和生长发育的影响,为龙脑樟的合理施肥提供科学依据。
1 材料与方法
1.1 试验材料
以龙脑樟为研究材料,龙脑樟以矮林作业方式经营,将距地面10 cm以上的枝叶在每年的10月下旬至11月上旬的时候全部砍除,待来年春季又重新萌发枝叶。试验地位于吉安市青原区吉安市林业科学研究所经营山场内,土壤 PH 4.5,有机质18.15 g/kg,碱解氮71.16mg/kg,有效磷53.7mg/kg,速效钾84.71 mg/kg。
1.2 研究方法
1.2.1 试验设计 采用“3414”不完全正交回归设计方案,设置N、P、K的3个因素,各因素均设4个水平用量,其中0水平是不施肥,2水平当地最佳施肥量,1水平=0.5×2水平,3水平=1.5×2水平(过量施肥水平),共14个处理,每个处理10株,3次重复。试验设计及施肥处理详见表1,其中为氮肥为尿素(N%=46%),磷肥为钙镁磷(P205%=12%),钾肥为氯化钾(K20%=63%),因在5月、6月和9月为龙脑樟生长的两个高峰期,故在4月中旬采用每株沟施法进行施肥。
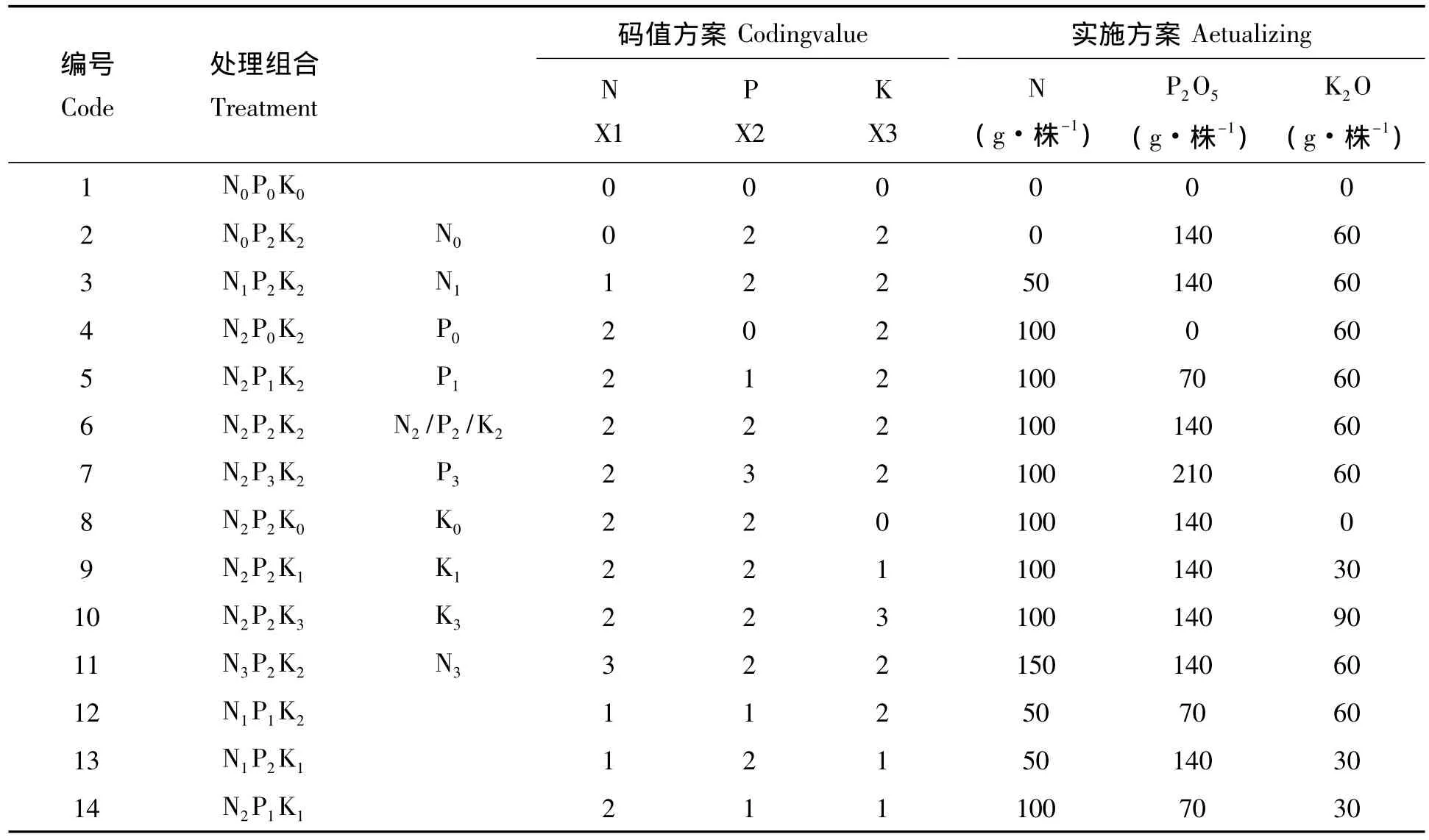
表1 施肥处理的试验设计Tab.1 The experimental design of fertilizing treatments
1.2.2 试验设计指标测定 试验于2014年7月中旬晴天上午09:00—11:00,每个处理选择3株,用美国生产的Li-6400便携式光合测定仪测定同一高度龙脑樟成熟叶片的光合作用参数,包括净光合速率(Pn)、胞间CO2浓度(Ci)和蒸腾速率(Tr)等。
同时,用美国生产的Li-6400便携式光合测定系统的自动光曲线程序来测定龙脑樟的光响应曲线。将光合有效辐射(PAR)梯度设定为:2 000、1 750、1 500、1 250、1 000、7 250、500、250、200、100、50、40、20、10、0μmol/(m2·s),测定在不同光合有效辐射下的净光合速率(Pn),绘制N、P、K不同施肥处理下龙脑樟的的光响应曲线。根据光响应曲线得到拟合参数(最大净光合速率Amax、暗呼吸速率Rd、光饱和点LSP、光补偿点LCP和表观量子效率AQY),并比较不同施肥处理下的差异。不同施肥处理下龙脑樟的表观量子效率(AQY)[21-22]可通过光合-光响应曲线在低光量子通量密度下的净光合速率做直线回归求得。在测定净光合速率后,取其叶片测定叶绿素含量[23]。
1.3 数据分析
采用SPSS 17.0进行数据统计分析,采用Origin 8.0制图。
2 结果与分析
2.1 不同水平N、P、K对龙脑樟叶片光合特性的影响
从表2可以看出,N、P、K肥不同施用量都对龙脑樟叶片的净光合速率(Pn)产生显著影响。对龙脑樟净光合速率影响最大的是N肥,其次是K肥,最后是P肥。在N2P2K2处理时净光合速率达到了最大值(14.37μmol/(m2·s)),该处理的净光合速率显著高于 N0P2K2、N1P2K2、N3P2K2、N2P0K2、N2P2K0和N2P2K1的处理。龙脑樟净光合速率随着N肥量的增加呈现“先增大后减小”的趋势,可见合理施N肥可以提高龙脑樟叶片净光合速率,不足或过量都会抑制龙脑樟的光合作用。而P肥和K肥对龙脑樟的净光合速率的影响不如氮肥显著。
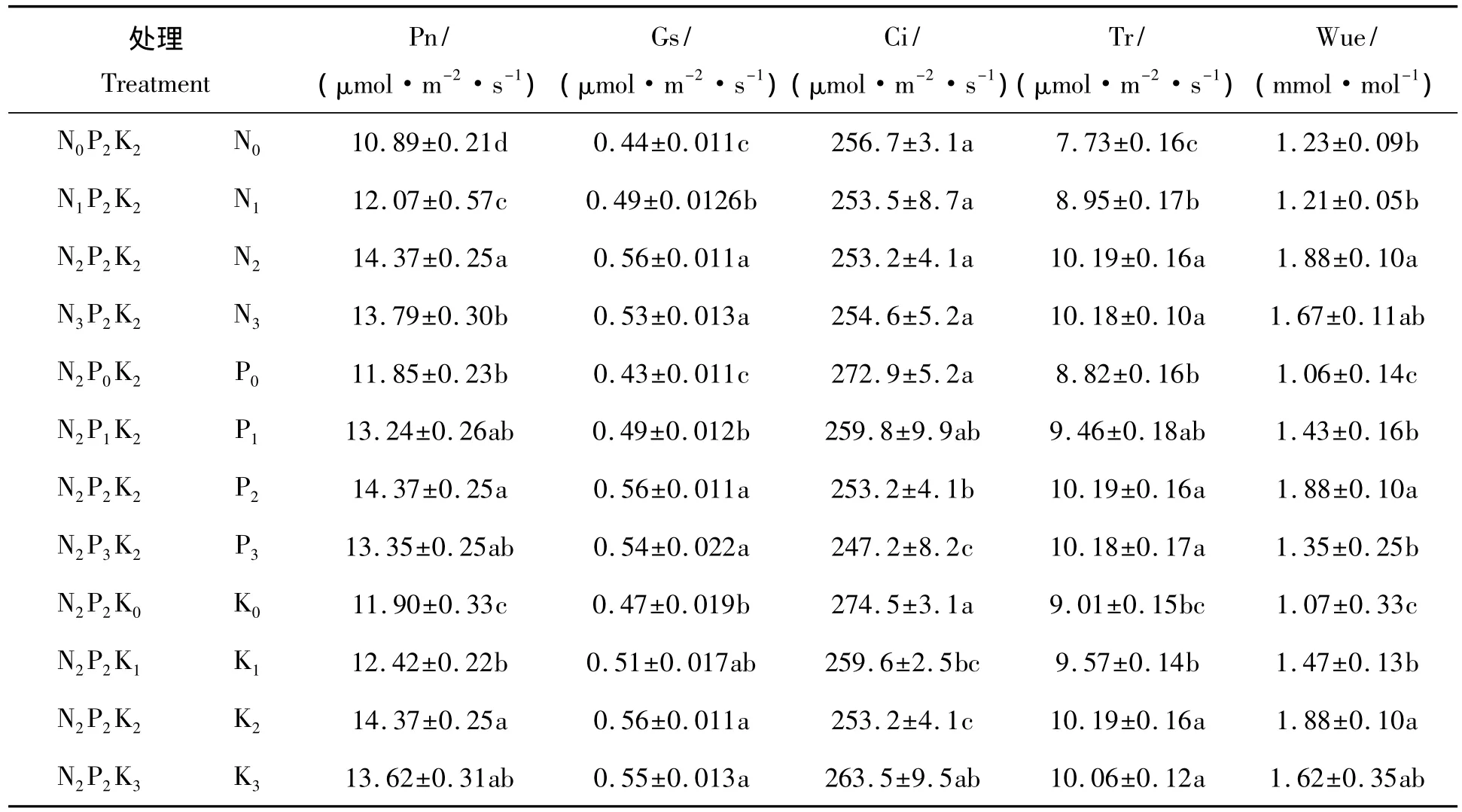
表2 N、P、K不同配施对龙脑樟叶片光合特性影响的单因素分析Tab.2 Single-factor analysis of photosynthetic characteristics by different N,P and K supp ly level in C.camphora
施肥处理对龙脑樟叶片气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)等的影响差异显著,但其变化趋势却各有差异(表2)。通过表3可知所有施肥处理中,净光合速率(Pn)与气孔导度(Gs)、水分利用效率(WUE)大小呈正相关,相关系数分别为0.784和0.516;气孔导度(Gs)与蒸腾速率(Tr)大小呈正相关,相关系数为0.416,与胞间CO2浓度(Ci)大小呈负相关,相关系数为-0.410。根据“3414”试验设计的特点,把N、P和K 3个因素中的1个固定为2水平,对另两个因素用量效果可以进行交互效应分析,由此可知N、P、K肥之间交互效应不明显,但龙脑樟净光合速率的大小受N肥施用量的影响大于P肥和K肥。
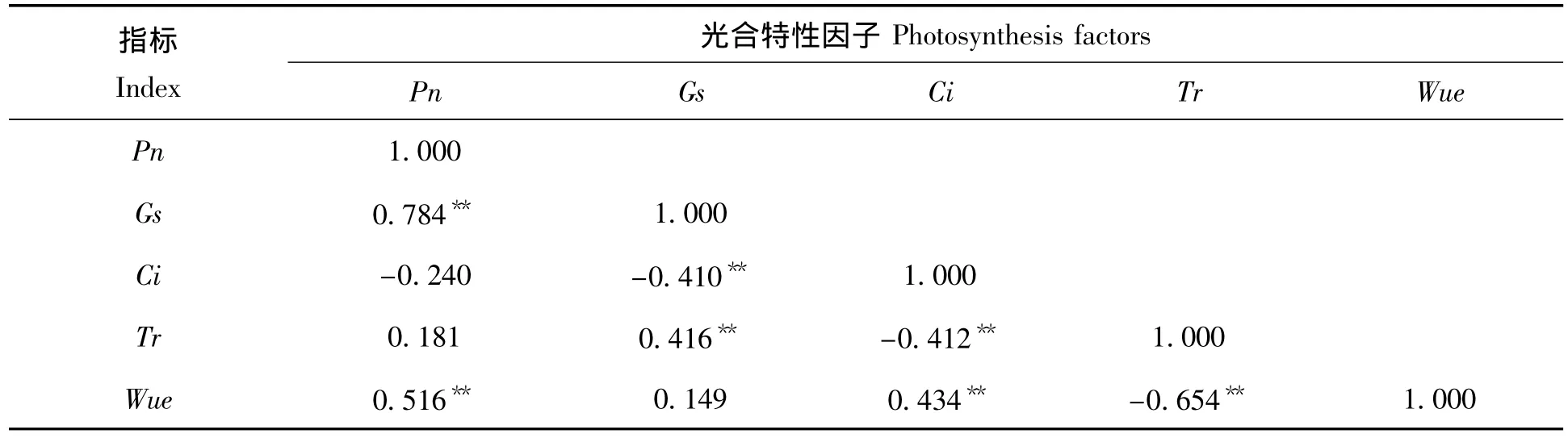
表3 主要光合特性因子间的相关性分析Tab.3 Correlation analysis among themajor photosynthesis factors
2.2 N、P、K对龙脑樟光合-光响应曲线的影响
不同施肥处理龙脑樟叶片对光照强度的响应存在一定的差异。根据实际观测结果拟合光合作用曲线如图1所示。从图1可知龙脑樟叶片的光合速率随光强上升而升高,超过一定范围后,光合速率不再上升。龙脑樟不同施肥处理下的光响应曲线特征参数(表4)可以通过光响应曲线结合低光强度下线性回归分析而得到。由表4可知龙脑樟叶片的暗呼吸速率(Rd)随着氮肥的增加而降低,N0(3.194 1 μmol/(m2·s))>N1(2.405 μmol/(m2·s))>N2(2.067 μmol/(m2·s))> N3(1.911 μmol/(m2·s));最大净光合速率(Amax)、光饱和点(LSP)和表观量子效率(AQY)呈先增后降的趋势,均在N2水平达到最高,其值分别是 Amax为15.145 μmol/(m2·s),LSP 为1245 μmol/(m2·s),AQY 为0.044 16。光补偿点(LCP)则是逐渐降低的,磷肥钾肥对光合-光响应曲线拟合参数的影响趋势相似,其中,氮肥对这些参数的影响均比磷肥和钾肥显著。
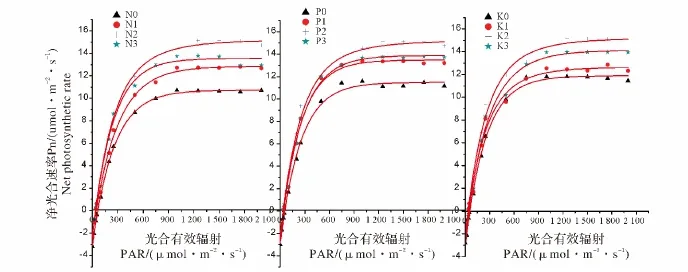
图1 龙脑樟光合-光响应曲线Fig.1 Netphotosynthetic rate-light response curves in leaves of C.camphora
2.3 不同水平N、P、K对龙脑樟的叶绿素含量影响
从表5可以看出,龙脑樟叶片叶绿素含量在一定的范围内随施肥量的增加而增加。N、P肥施用量对龙脑樟叶片叶绿素含量有显著性影响,随着N、P施肥量的增加,龙脑樟的叶绿素含量增加,在施肥量为中等时达到最大,此后叶绿素含量随施肥量的增加反而有所下降,表明龙脑樟不耐高肥力。钾肥施用量对幼苗叶片叶绿素含量影响差异不显著。N、P、K肥施用量对叶绿素含量的影响趋势相似。
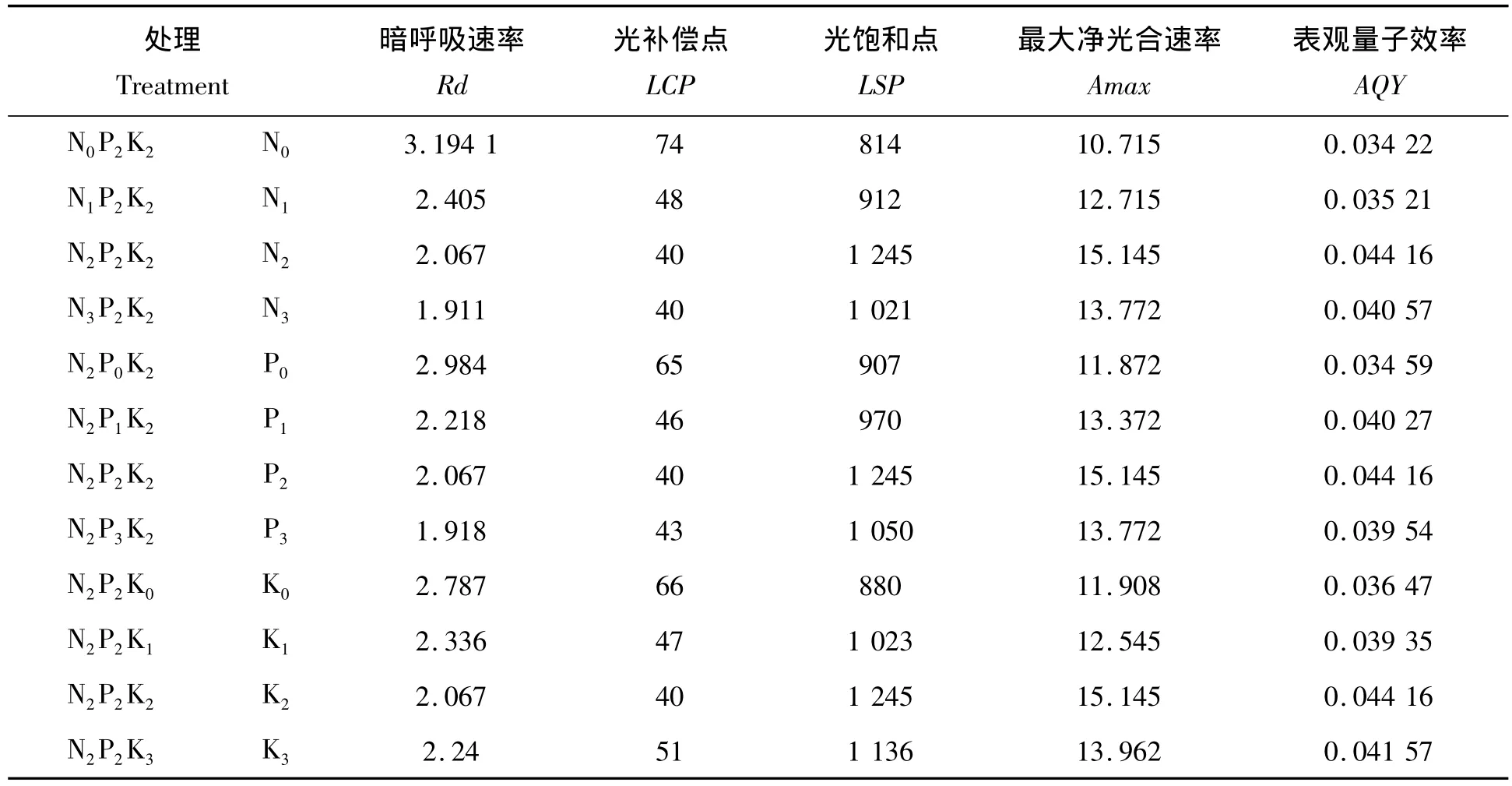
表4 N、P、K不同配施对龙脑樟光响应曲线特征参数的影响Tab.4 Effect of charactererstic parameters of light response curve under different N,P and K supply level in C.camphora
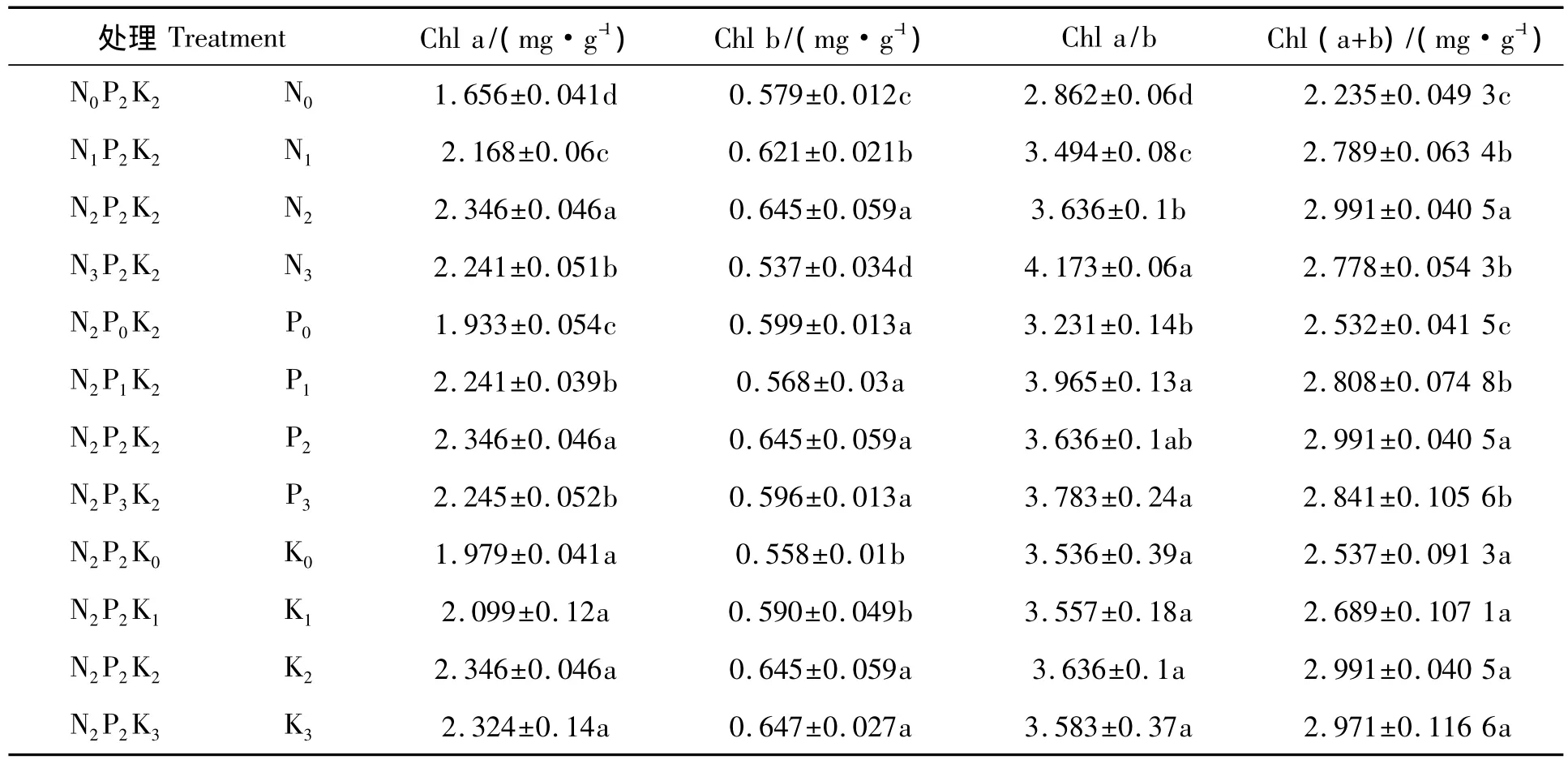
表5 N、P、K不同配施对龙脑樟叶绿素含量的影响Tab.5 The effect of chlorophyll content by different N,P and K supp ly level in C.camphora(n=3)
3 讨论与结论
氮肥能显著增加龙脑樟叶片的净光合速率和叶绿素含量,它们变化趋势基本是一致的,即在一定的范围内,龙脑樟的净光合速率(Pn)和叶绿素含量(Chl)随着施氮量的加大而增大,过量时反而减小,适量的氮肥可以增加叶绿素含量及光合作用,氮肥过量使后期叶绿素含量及光合作用降低,在N2(2.991 mg/g和14.37μmol/(m2·s))水平均达到了最大值。得出这种结果的原因可能是氮肥能提高对龙脑樟氮素的供应,使叶片可以合成更多的叶绿素,从而可以促进其光合作用[24];施肥量过高导致龙脑樟植株内氮素浓度持续上升,加快了叶片的衰老,致使参与光合作用的一些蛋白质和Rubisco酶含量有所提高[25],从而使光合速率下降。光合作用是决定作物产量的主要因素,提高光合速率是龙脑樟高产栽培的生理基础,叶绿素是植物光合作用的物质基础,龙脑樟叶片的净光合速率和叶绿素含量呈正相关关系,与魏典典[20]等人的研究结果一致。在本研究中,当氮(N 100 g/株)、磷(P2O5140 g/株)、钾(K2O 60 g/株)时,龙脑樟叶片的净光合速率和叶绿素含量显著高于其他施肥处理。
适量氮肥在一定程度上提高了龙脑樟的渗透调节和气孔调节能力,增加了龙脑樟叶片的气孔导度。在本研究中,处理 N2P0K2、N2P1K2、N2P2K2、N2P3K2、N2P2K0、N2P2K1、N2P2K3皆为 N2水平处理,但是只有N2P2K2处理时其光合速率和叶绿素含量皆为最高,可见氮施肥量相同,而磷、钾肥不同的施肥配比则有不同的结果。导致这种结果的原因其一是适量的增施磷肥能促进植株体内的氮素向叶片转移,进而提高叶片叶绿素含量和净光合速率[26];其二是钾元素可提高植株叶片中可溶性蛋白和叶绿素含量[27],从而可以使叶片光合能力增强;并且钾对镁元素吸收的影响,表现在低浓度条件时为协同作用,而高浓度条件下则是拮抗作用[28],而镁元素是合成叶绿素的重要元素;其三是磷肥与钾肥对植株光合作用的影响与施氮肥的量有着密切的联系,科学合理的配施是增强光合作用的关键[29-30]。
施肥对龙脑樟叶片的光饱和点(LSP)和表观量子效率(AQY)产生了显著的促进作用,且在适宜的范围内随施肥量的增加其值越大,在N2P2K2处理达到最高(1 245,0.044 16),并且其光补偿点最低(40),如表4所示。光补偿点低且光饱和点高的植物能适应多种光环境。光饱和点增大,光补偿点降低可以使龙脑樟叶片的光合时效延长,从而积累更多的光合产物,有利于龙脑樟的生长发育。表观量子效率可反映叶片转化光能的效率,表观量子效率值越大说明叶片转化光能的能力越高。施肥量过高,龙脑樟的光饱和和表观量子效率有所下降,由此可推测,适度水平的氮磷钾肥配施能提高林木的光饱和点,增大表观量子效率,降低光补偿点,过高水平氮磷钾肥配施对光合作用的促进作用则会减弱,具体原因有待进一步研究。
施肥处理显著影响了龙脑樟叶片的光合特性,合理的施肥使龙脑樟叶片的光合速率显著高于缺素施肥处理,如表2所示的缺氮处理中,N1、N2和N3时龙脑樟叶片的光合速率显著高于N0处理,光合速率分别是N0水平的111%、132%、127%。氮磷钾肥不同处理对龙脑樟叶片的净光合速率(Pn)、叶片气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等都产生了显著影响,其影响大小为:氮肥、磷肥、钾肥。本文分析了氮磷钾肥不同处理对龙脑樟的影响,得到龙脑樟光合生理促进作用最显著的是N2P2K2处理。本试验对氮磷钾不同配比施肥对龙脑樟叶片净光合速率(Pn)、胞间CO2浓度(Ci)等光合特性的影响进行了探讨,在今后的研究中,还需进一步分析龙脑樟的需肥规律和分配等对其生长和生理的影响,设置出更优化的氮磷钾配比,为龙脑樟产业的发展提供有效的科学依据。
[1]祁承经,汤庚国.树木学:南方本[M].北京:中国林业出版社,2005.
[2]陈美兰,华永丽,黄璐琦,等.龙脑樟有性繁殖后代叶油分析[J].中国中医药信息志,2010,17(8):37-40.
[3]陈红梅,孙凌峰.江西吉安龙脑樟资源开发与利用前景[J].林业科学,2006,42(3):94-98.
[4]刘冰.自营龙脑樟原料林生产天然冰片的经济效益分析[J].江西林业科技,2013(3):49-50.
[5]郑小春,郭志文,龙光远,等.龙脑樟容器扦插育苗技术研究[J].江西林业科技,2012(3):35-36.
[6]郭照光,孙秀泉,周晓勤,等.龙脑樟树快繁育苗技术的研究与应用[J].湖南林业科技,2003(3):44-46.
[7]陈美兰,叶正良,欧阳少林,等.龙脑樟愈伤组织的诱导及龙脑的产生[J].中国中药志,2012,35(5):558-560.
[8]邹娜,曹福亮,张露,等.龙脑樟愈伤组织诱导及增殖研究[J].江西林业科技,2014(4):1-3.
[9]胡文杰,江香梅,杨海宽,等.脑樟不同部位精油成分及其含量变化研究[J].西北农林科技大学学报:自然科学版,2014,42(10):126-132.
[10]张宇思,王成章,周昊,等.不同产地龙脑樟叶挥发油成分的GC-MS分析[J].中国实验方剂学志,2014,20(10):54-58.
[11]欧阳少林,赵小宁,李楚文,等.龙脑樟果实挥发油成分气相色谱-质谱分析[J].中国中医药信志,2013,20(11):58-60.
[12]刘塔斯,龚力民,郭英,等.GC-MS测定龙脑樟植物不同部位右旋龙脑的含量[J].中国中药志,2009,34(13):1692-1694.
[13]李菲菲,方静,马琼,等.龙脑液导致癌细胞凋亡的实验研究[J].中国生物工程杂志,2013,33(5):22-27.
[14]Damian Badillo LM,Martinez Mu oz R E,Salgado Garciglia R,et al.In vitro antioomycete activity of Artemisia ludoviciana extracts against Phytophthora spp[J].Boletin Latinoamericano y del Caribe de Plantas Medicinalesy Aromaticas,2010,9(2):136-142.
[15]Jadhav M V,Sharma R C,Rathore Mansee G A K.Effectof Cinnamomum camphora on human sperm motility and sperm viability[J].JClin Res Lett,2010,1(1):01-10.
[16]龙光远,彭招兰,郭德选,等.龙脑樟矮林基地施肥试验研究[J].江西林业科技,2000(1):14-16.
[17]舒翔,范川,李贤伟,等.施肥对香樟幼苗光合生理的影响[J].四川农业大学学报,2013.31(2):157-161.
[18]赵燕,董雯怡,张志毅,等.施肥对毛白杨杂种无性系幼苗生长和光合的影响[J].林业科学,2010,46(4):70-77.
[19]黄露,甘润朕,张涛,等.施肥对桑树夏季生长叶片的光合特性影响[J].蚕业科学,2013,39(6):1056-1065.
[20]魏典典,张刚,刘淑明,等.配方施肥对文冠果光合作用的影响[J].西北林学院学报,2014,29(3)27-31.
[21]Yu SW.Plantphysiology andmolecular biology[M].Beijing:Science Press,1992.
[22]XuD Q.Non-uniform stomatal closure and non-stomatal limitation of photosynthesis[J].Plant Physiology Communications,1995,31(4):246-252.
[23]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,1999.
[24]邹振华,党宁,王惠群,等.不同氮素水平对营养生长期南荻植株光合特性的影响[J].作物研究,2012,26(3):255-259.
[25]王冉,何茜,丁晓纲,等.N素指数施肥对沉香苗期光合生理特性的影响[J].北京林业大学学报,2011,33(6):58-64.
[26]陈琳,曾杰,贾宏炎,等.林木苗期营养诊断与施肥研究进展[J].世界林业研究,2012,25(3):26-31.
[27]胡春梅,王秀峰,季俊杰.钾对瓜尔豆光合及胚乳中糖类含量的影响[J].植物营养与肥料学报,2006,12(6):858-863.
[28]徐刚,彭天沁.不同基质含水量和钾肥施用量对黄瓜生长及光合作用的影响[J].江苏农业学报,2014,30(5):1109-1114.
[29]罗凡,张厅,龚雪蛟,等.不同施肥方式对茶树新梢氮磷钾含量及光合生理的影响[J].应用生态学报,2014,25(12):3499-3506.
[30]蔡伟建,窦霄,高捍东,等.氮磷钾配比施肥对杂交鹅掌楸幼林初期生长的影响[J].南京林业大学学报:自然科学版,2011,35(4):27-33.
[31]张恩平,李天来,葛晓光,等.钾营养对番茄光合生理及氮磷钾吸收动态的影响[J].沈阳农业大学学报,2005,36(5):532-535.
