三北地区冷水鱼常见病原菌的分布及药敏试验
2015-05-28连浩淼李绍戊卢彤岩
连浩淼,李绍戊,张 辉*,卢彤岩
(1.东北农业大学 动物科技学院,黑龙江 哈尔滨 150030;2.中国水产科学研究院 黑龙江水产研究所,黑龙江 哈尔滨150070)
冷水性鱼类以鲑鳟鱼类为代表的,只能在洁净、清冷、无污染的流动水域中生长的名贵鱼类的总称。我国冷水性鱼类约有88种(亚种),其中可开发利用的经济品种较多,主要有大麻哈鱼、鲟鱼、鳇鱼、河鳟、红点鲑、哲罗鲑、细鳞鲑、虎嘉鱼、黑龙江茴鱼、雅罗鱼、狗鱼、大银鱼、裂腹鱼类、重唇鱼、江雪等 50 多种[1]。
我国冷水域开阔,这为我国冷水性鱼类的养殖和生产提供环境优势。但随着冷水鱼养殖密度不断增大,养殖技术欠缺及管理调控水平低,冷水鱼病害问题愈加严重,给冷水鱼养殖带来严重的经济损失。密集养殖的情况下给鱼类养殖环境造成压力,长期密集养殖会造成免疫抑制和疾病暴发[2],其中细菌性疾病成为冷水鱼养殖中重要的病害之一。有研究曾报道常见细菌性疾病包括烂鳃、烂鳍、疥疮、细菌性败血症、肠炎等,已有学者对一个地区冷水鱼细菌病病原菌进行研究[3]以及多个地区一种病原菌的分离研究[4],2013年5—8月笔者于三北地区冷水鱼养殖场进行流行病学调查,对患病的虹鳟、大西洋鲑、银鲑、西伯利亚鲟等进行病原菌的分离鉴定及药敏试验,以期为三北地区冷水鱼细菌病防控提供参考。
1 材料与方法
1.1 病鱼症状及样品采集
2013年5—8月,在辽宁本溪、牡丹江、青海、北京房山地区分别采集患病银鲑鱼、大西洋鲑、虹鳟、西伯利亚鲟,采集的具体信息见表1。

表1 2013年5月至2013年8月份采集样品信息Tab.1 Information of fish sam ples collected in 2013.5—2013.8
1.2 培养基及主要试剂
实验用胰蛋白胨大豆琼脂培养基(TSA)/胰蛋白胨大豆肉汤(TSB)/脑心浸出液肉汤(BHI)/BHI琼脂培养基/MH肉汤/Mueller-Hinton琼脂培养基购于青岛科技园海博生物技术有限公司;PCR所用试剂及特异性引物购自于上海生工生物工程有限公司;API 20NE、API 20E、API 20Strep试剂条购自于北京威泰科生物技术有限公司;细菌基因组DNA提取试剂盒购自于天根生化科技有限公司;革兰染液、抗菌药物药敏纸片购于杭州天和微生物有限公司。
1.3 病原菌分离与接种
在无菌条件下从患病鱼肝脏、脾脏、大肠处取样划线接种于培养基琼脂平板上,28℃倒置培养24 h,分别挑取单个优势菌落在琼脂平板上纯化培养,获得纯培养的89株菌株。肉眼观察菌落形态、大小、颜色等,同时革兰染色光学显微镜观察细菌形态特征;各项理化指标的测定参照法国生物梅里埃公司API 20NE、API20E、API20 Strep试剂条进行。挑取形态较好的单菌落接种于3mL肉汤培养基中28℃200 r/min震荡12 h后取菌液和甘油4∶1的比例混合均匀,保存于-80℃冰箱备用。
1.4 16S rRNA基因序列测定
无菌条件下,将89株菌接种于TSB肉汤培养基中,28℃ 200 r/min震摇16 h后离心集菌,按照细菌基因组DNA提取试剂盒说明书提取细菌基因组DNA作为PCR模板DNA。利用细菌16S rRNA通用引物F27:5′-AGAGTTTGATCMTGGCTCAG-3′和 R1492:5′-GRTACCTTGTTACGACTT-3′对 89 株菌 16S rRNA 基因进行扩增[5]。PCR反应条件为:94℃预变性5min;94℃变性1min,54℃退火1 min,72℃延伸2 min,共35个循环;72℃延伸10min。PCR产物经10 g/L琼脂糖凝胶电泳检测,电压120 V,电泳25 min,保存图片用于查看分析。PCR产物经纯化后由上海生工生物工程股份有限公司进行序列测定。
1.5 药敏试验
1.5.1 供试抗菌药物 药敏实验中测定了常见18种抗菌药物:氨苄西林(ampicillin)、阿莫西林(amoxicillin)、诺氟沙星(norfloxacin)、氧氟沙星(ofloxacin)、左氧氟沙星(levofloxacin)、环丙沙星 (ciprofloxacin)、恩诺沙星(enrofloxacin)、萘啶酸(nalidixic acid)、庆大霉素(gentamicin)、链霉素(streptomycin)、卡那霉素(kanamycin)、四环素(tetracycline)、红霉素(erythromycin)、氯霉素(chloramphenicol)、复方新诺明(compound sulfamethoxazole)、呋喃唑酮(furazolidone)、利福平(rifampicin)、新霉素(neomycin),以上药敏纸片均购自于杭州天和微生物有限公司。
1.5.2 药敏试验操作 药敏试验采用K-B纸片法进行。无菌条件下将89株菌挑入MH肉汤培养基中,28℃ 200 r/min震荡培养16 h后将菌液浓度调至1.0×107CFU/mL。取0.1 mL培养菌液均匀涂布于9 cm的MH琼脂平板上,然后等距离贴上药敏纸片4片/平板,分别测定分离菌株对18种常见药物的敏感程度。将贴合好的平板倒置于培养箱中,28℃培养24 h,测定其抑菌圈直径(平行重复3次),并根据CLSI标准进行药敏结果判定[6]。
2 结果与分析
2.1 菌株分离鉴定结果
本研究从辽宁本溪分离病原菌13株,牡丹江分离病原菌32株,青海分离病原菌23株,北京房山分离病原菌21株,共分离菌株89株,并应用API20试剂条对部分菌株进行生化鉴定,菌株分离及生化鉴定信息见表2。

表2 部分菌株分离鉴定的相关信息Tab.2 Isolation and identification of part bacterial strains in this study
2.2 16S rRNA基因的PCR扩增及比对结果
选取2013年5—8月采集分离病原菌的89株,采用PCR方法扩增细菌16S rRNA基因,通过测序比对鉴定方法对其进行初步鉴定。图1为部分病原菌16S rRNA基因PCR扩增后的电泳图片。从图中可以看出,扩增片段大小约为1 500 bp,与目标片段大小相同。对所得的16S rRNA片段进行测序,将所得到的序列通过Genbank进行比对分析,对所分离的病原菌得到初步鉴定结果。
图1 16S rRNA PCR扩增产物电泳图谱Fig.1 Electrophoresis pattern of 16S rRNA PCR amplified products
2.3 药敏试验结果
89株病原菌作为供试菌株进行药敏实验,结果表明相同地区同种菌株药物敏感程度结果基本一致;不同地区相同菌株对药物的敏感程度有差别;其中,氨苄西林除对青海地区黄杆菌、鲁氏耶尔森菌,北京房山海豚链球菌抑菌外,对其他菌株不抑菌;阿莫西林除对青海黄杆菌,北京海豚链球菌和停乳链球菌抑菌外,对其他菌株均没有抑菌作用。地区代表菌株的抑菌圈直径见表3。
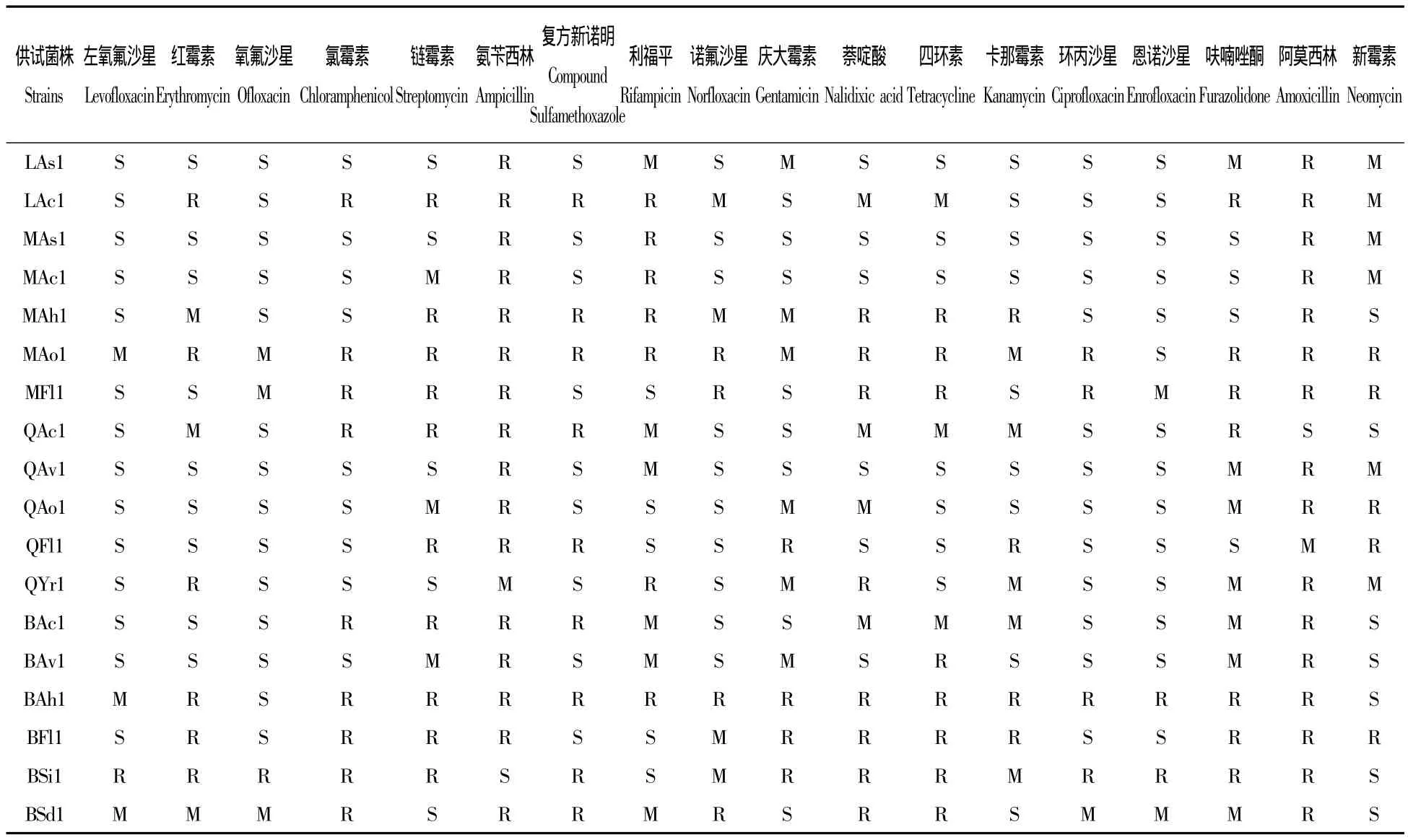
表3 部分菌株药敏实验结果Tab.3 Antimicrobial susceptibility test of parts of isolated bacteria in this study
根据药敏实验结果,统计了89株病原菌的药物敏感比例,不存在菌株对庆大霉素、呋喃唑酮、阿莫西林和新霉素高度敏感,所有菌株对左氧氟沙星均敏感(表4)。

表4 89株菌的药物敏感程度百分比Tab.4 The percentage of drug sensitivity to 89 strains
根据表4病原菌对抗生素敏感程度的统计,得出细菌耐药比率分析图(图2),菌株对不同种类抗生素耐药的情况可以看出,供试的8个种类89株菌种中,有92.13%和96.63%的菌株分别对氨苄西林和阿莫西林具有很强的耐药性,有100%、93.26%、91.10%、92.13%、91.01%和94.38%的菌株分别对左氧氟沙星、氧氟沙星、庆大霉素、卡那霉素、环丙沙星和恩诺沙星敏感,其中对左氧氟沙星无耐药菌株。
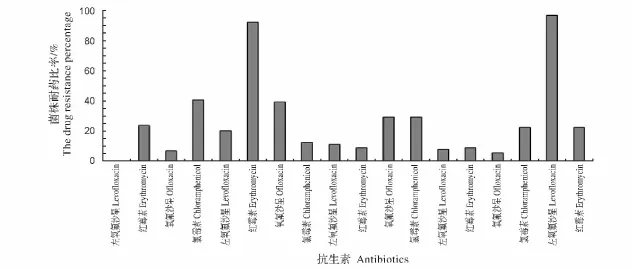
图2 细菌耐药所占比率Fig.2 The drug resistance percentage of isolated bacteria
3 结论与讨论
2013年5—8月由于气温逐渐升高,是冷水鱼细菌性疾病的多发时期。本研究从三北地区地区患病鱼体的肝脏、肾脏、脾脏、肠道采集并分离得到了89株病原菌,根据形态学特征和理化特性试验并结合16S rRNA基因检测对病原菌进行了初步鉴定,分离到的菌种为杀鲑气单胞菌、嗜水气单胞菌、温和气单胞菌、维氏气单胞菌、不动杆菌、黄杆菌、海豚链球菌及停乳链球菌,从辽宁本溪银鲑与牡丹江大西洋鲑分离得到的病原菌多为杀鲑气单胞菌,从青海虹鳟体内分离得到的菌株多为温和气单胞菌和鲁氏耶尔森菌,从北京房山西伯利亚鲟中分离到的菌株主要以停乳链球菌为主,已有学者对上述病原菌流行症状进行研究。
杀鲑气单胞菌分布地域广,宿主范围也很广,是多种水产动物的原发性病原菌,可感染各个年龄段的海淡水鱼类,每年对鲑鳟鱼类造成十分严重的损失[7-8],曹成易等[9]报道了杀鲑气单胞菌能引起大西洋鲑体表溃烂;温和气单胞菌广泛存在于水、土壤中,属条件性致病菌,其主要的致病因子是产生各种溶血素和肠毒素,具有发病率高、感染快、传播快、死亡快的特点[10-11]。刘敏等[12]报道了温和气单胞菌引起了鲤鱼体表溃烂疾病;维氏气单胞菌为人鱼共患病致病菌,人食用了该细菌污染的水产品时,可引发腹泻、脑膜炎和败血症等,免疫力低下的患者甚至能死于此菌的感染[13]。Sung等[14]报道了维氏气单胞菌能引起对虾体表严重出血、死亡的病症;鲁氏耶尔森菌为冷水性鲑鳟鱼的常见病原菌[15-16],几乎对所有养殖的鲑鳟鱼类都有感染性[17]。并且范方玲等[18]有过斑点叉尾鮰感染鲁氏耶尔森菌引起体表及内脏出血的报道;司力娜等[19]报道了嗜水气单胞菌引起鲤鱼败血症,目前嗜水气单胞菌仍然是引起鲤鲫鱼细菌性败血症的罪魁祸首;舍晓丽等[20]有过斑点叉尾鮰感染海豚链球菌的报道;顾天钊等[21]曾报道过鲍氏不动杆菌引起鳜鱼肝脏严重病变至死亡;潘厚军等[22]报道了停乳链球菌引起西伯利亚鲟鱼内脏出血及腹水等症状。根据有关报道这些病原菌已经成为水产动物的致病菌,对水产的健康养殖带来了严重危害。此次流行病学调查过程表明,每种病鱼分别感染多种细菌而致病。
研究表明,不同地区的相同菌株API 20的鉴定结果有所差异,而造成这种差异的原因可能是地域环境差异等方面造成的,这与刘礼辉等[23]阐述的分离的柱状黄杆菌有所差异是与地区、气候、水质条件及实验室培养条件等方面的不同而产生的结果相似。16S rRNA序列法是一种现代的基于细菌遗传特性的细菌鉴定方法[24],并且也广泛应用于水产细菌分子鉴定。本研究中不同地区相同菌株的16S rRNA结果基本一致,此结果验证了王荻等[25]的研究,不同地区的同种菌株保守区片段变化较小,同源性极高聚类不明显,没有形成明显的区域性或特异性变化。
国内对水产养殖中细菌性疾病的防治仍然依赖于抗生素。药敏实验结果表明,分离得到的89株菌株对氨苄西林和阿莫西林有较强的耐药性,对左氧氟沙星、氧氟沙星、庆大霉素、卡那霉素、环丙沙星和恩诺沙星高度敏感,对链霉素、利福平、诺氟沙星、萘啶酸、四环素、呋喃唑酮、新霉素均有不同程度的耐药,其中没有菌株对左氧氟沙星耐药,表现为最敏感。并且不同地区的相同病原菌对抗生素的敏感程度也有一定程度的差异。这也验证了林春友等[26]淡水养殖鱼类病原菌药敏试验中的结论,不同抗菌药物对同一病原菌的MIC不同,同一抗菌药物对不同病原菌的MIC也不同。青霉素类抗生素药物对革兰氏阳性菌较为敏感,而冷水鱼中的病原菌以革兰氏阴性菌为主,因此表现为不敏感。除了左氧氟沙星以外其他种类抗生素都有不同程度的耐药情况出现,这样高耐药情况提示在水产养殖过程中不能盲目使用抗生素药物,要对症下药,并且科学控制单次用药的时间和剂量,避免再次用药的高耐药情况,在用药的同时,做好饲养过程的调控和消毒工作,并且保持饲料的新鲜度,以及积极进行冷水鱼细菌性疾病疫苗的研发,这样更能达到防治细菌性疾病的目的。本研究通过对三北地区冷水鱼常见病原菌分离鉴定,表明不同地区同种菌株有共同特性同时也存在着差异,结合耐药性研究结果,以期对三北地区冷水鱼疾病防治以及实现冷水鱼健康养殖提供参考。
[1]纪锋,王炳谦,孙大江,等.我国冷水性鱼类产业现状及发展趋势探讨[J].水产学杂志,2012,25(3):63-68.
[2]Luo Lin,Cai Xuefeng,He Chunan,et al.Immune respone,stress resistance and bacterial challenge in juvenile rainbow trouts Oncorhynchusmykiss fed diets containing chitosan-oligosaccharides[J].Current Zoology,2009,55(6):1-14.
[3]高阳,王俊刚,顾燕玲,等.新疆额尔齐斯河流域冷水鱼肠道耐低温乳酸菌的分离筛选及其遗传差异[J].微生物学报,2013,53(1):82-91.
[4]Clifford E,Starliper.Bacterial coldwater disease of fishes caused by Flavobacterium psychrophilum[J].Journal of Advanced Research,2011,2(2):97-108.
[5]Lane D J.16S/23S rRNA sequencing[M]//Stackebrandt E,Goodfellow M(eds).Nucleic acid techniques in bacterial systematics.Chichester:Wiley,1991:115-175.
[6]National Committee for Clinical Laboratory Standards.Performance standards for antimicrobial disc susceptibility tests[M].Villanova,PA,USA:Approved standard NCCLS publications M2-A5,1993.
[7]丁雷,岳永生,宋憬愚.虹鳟皮肤溃烂病的病原研究[J].淡水渔业,2002,32(3):28-30.
[8]Wiklund T,Dalsgaard I.Occurrence and significance of atypical Aeromonas salmonicida in non-salmonid and salmonid fish species:A review[J].Diseases of Aquatic Organisms,1998,32:49-69.
[9]曹成易,汪开毓,王玲,等.大西洋鲑杀鲑气单胞菌的分离鉴定[J].淡水渔业,2009,39(1):54-57.
[10]胡琳琳,房文红,梁思成.金鱼温和气单胞菌的分离鉴定及药敏试验[J].上海水产大学学报,2008,17(3):285-290.
[11]韩继卫,罗文,郑大恒.黄颡鱼温和气单胞菌的分离鉴定和药敏试验[J].绍兴文理学院学报,2011,31(9):19-21.
[12]刘敏,韩英.鲤鱼温和气单胞菌的分离鉴定和药敏试验[J].东北农业大学学报,2005,36(4):486-489.
[13]Fraisse T,Lech iche C,Sotto A,et al.Aeromonas spp.infections:retrospective study in nimes university hospital,1997-2004[J].Pathologie Biologie,2008,56:70-76.
[14]Sung H H,Hwang S F,Tasi F M.Responses of giant fresh water prawn(Macrobrachium rosenbergii)to challenge by two strains of Aeromonas spp.[J].Journal of Invertebrate Pathology,2000,76:278-284.
[15]Fouz B,Zarza C,Amaro C.First description of non-motile Yersinia ruckeri serovar I strains causing disease in rainbowtrout,Oncorhynchusmykiss(Walbaum),cultured in Spain[J].Journal of Fish Diseases,2006,29:339-346.
[16]Tobback E,Decostere A,Hermans K,et al.Yersinia ruckeri infections in salmonid fish[J].Journal of Fish Diseases,2007,30:257-268.
[17]张晓君.鲁氏耶尔森氏菌及鱼类相应感染症[J].河北科技师范学院学报,2004,18(3):77-80.
[18]范方玲,汪开毓,耿毅,等.斑点叉尾鮰 (Ictalunes punctatus)源鲁氏耶尔森氏菌的分离鉴定及系统发育分析[J].海洋与湖沼,2010,41(6):862-868.
[19]司力娜,李绍戊,王荻,等.东北三省15株致病性嗜水气单胞菌分离株的药敏实验分析[J].江西农业大学学报,2011,33(4):786-790.
[20]余晓丽,陈明,李超,等.斑点叉尾暴发性海豚链球菌病的研究[J].大连水产学院学报,2008,22(3):581-191.
[21]顾天钊,陆承平,陈怀靑,等.鲍氏不动杆菌-鳜鱼爆发性死亡的新病原[J].微生物学通报,1997,24(2):104-106.
[22]潘厚军,刘晓勇,常藕琴,等.西伯利亚鲟停乳链球菌的分离、鉴定与致病性[J].中国水产科学,2009,16(6):891-904.
[23]刘礼辉,李宁求,石存斌,等.斑点叉尾鮰触烂鳃病病原柱状黄杆菌的分离及鉴定[J].安徽农业科学,2008,36(17):7124-7126.
[24]Eddy F,Powell A,Gregory S,et al.A novel bacterial diseaseof the European shore crab,Carcinusmaenas-molecular pathology and epidemiology[J].Microbiology,2007,153:2839-2849.
[25]王荻,李绍戊,刘红柏,等.东北三省9株嗜水气单胞菌16S rDNA序列比较研究[J].水产学杂志,2010,23(2):11-15.
[26]林春友,王云祥.淡水养殖鱼类病原菌药敏实验[J].水利渔业,2004,24(3):58-59.
