密度对乐昌含笑幼苗的生长和生物量的影响
2015-05-22
(华南农业大学 林学院,广东 广州 510642)
密度通过植物对地上光照和地下养分资源的利用[1]改变植物间的竞争[2],制约着植物体内的养分分配格局,引起植物个体资源的重新划分[3],影响植物的生长[4]和生物量积累与分配[5]。目前,国内外关于种群密度的研究较多,主要是针对杉木Cunninghamia lanceolata、马尾松Pinus massoniana、杨 树Populus×euramericana、 桉 树Eucalyptus urophylla等树种[6-10]。林分密度研究对象多为多年生人工林,而对幼苗种植密度效应的研究较少[11]。
乐昌含笑Michelia chapensis是热带和亚热带地区的重要绿化造林树种,前人对该树种的研究主要集中在生理、混交林、遗传性质、土壤等方面[12-16]。目前我国关于这类造林树种的幼苗密度效应研究较少。本研究以乐昌含笑幼苗为研究对象,研究其不同种植密度下的生长和生物量变化,可以丰富该树种的种群动态知识。
1 材料与方法
1.1 试验地概况
幼苗盆栽密度试验地设在广州市华南农业大学跃进北苗圃(113º18′E,20º6′N),属于南亚热带季风气候,温暖多雨,夏长冬短。年平均温度、最冷月(1月)和最热月(7月)温度分别为21.8 ℃、13.3 ℃和28.1 ℃。年降雨量1 714.4 mm,4~9月降雨量占年降雨量的82%,年均相对湿度79%[17]。
1.2 试验材料
以广东省国森苗圃所提供的乐昌含笑1年生实生苗为试验材料。以1∶1的腐殖质土与黄土为基质,采用直径35 cm,深30 cm无纺布美植袋种植。设置4个种植密度:10、20、40、80株/m2(美植袋中分别种植1、2、4、8株幼苗),分别用密度I、II、III和IV表示。各种密度处理为15个重复以上。试验时间为2013年3月至2013年12月。幼苗的基本生长情况见表1。

表 1 幼苗的基本生长情况(平均值±标准差)Table 1 General growth of seedlings (cm)
1.3 研究方法
幼苗的形态生长包括地径、苗高和冠幅每个月测定一次。
在实验结束的时(2013年12月)测量各密度幼苗的生物量。树种每个密度随机选取15株幼苗,移出美植袋,清洗干净后用密封袋密封带回实验室。室后将植株分解为根、茎、枝和叶后称重。放入烘箱中于80 ℃烘48 h,称量各器官干重。
1.4 数据处理
对植物生长、幼苗生物量进行常规统计分析。主要应用Excel软件和SAS 9.3软件。
2 结果与分析
2.1 各密度幼苗的生长
在试验期内,乐昌含笑各月份的地径随着密度增大而减小,且各密度间的地径差在12月份达到最大(图1)。试验期内,密度I、II、III和IV的地径平均净生长量分别为7.79、7.23、6.01 mm和3.92 mm,年增长幅度分别为136%、127%、105%和69%。密度I平均生长率分别为密度II、III和IV的1.09、1.30和1.98倍。
在整个试验周期内,苗高随着密度增大而减小,密度I、II、III和IV的平均苗高生长分别为11.95、11.06、11.30 cm和6.94 cm(图2),增加幅度分别为31%、28%、29%和18%。
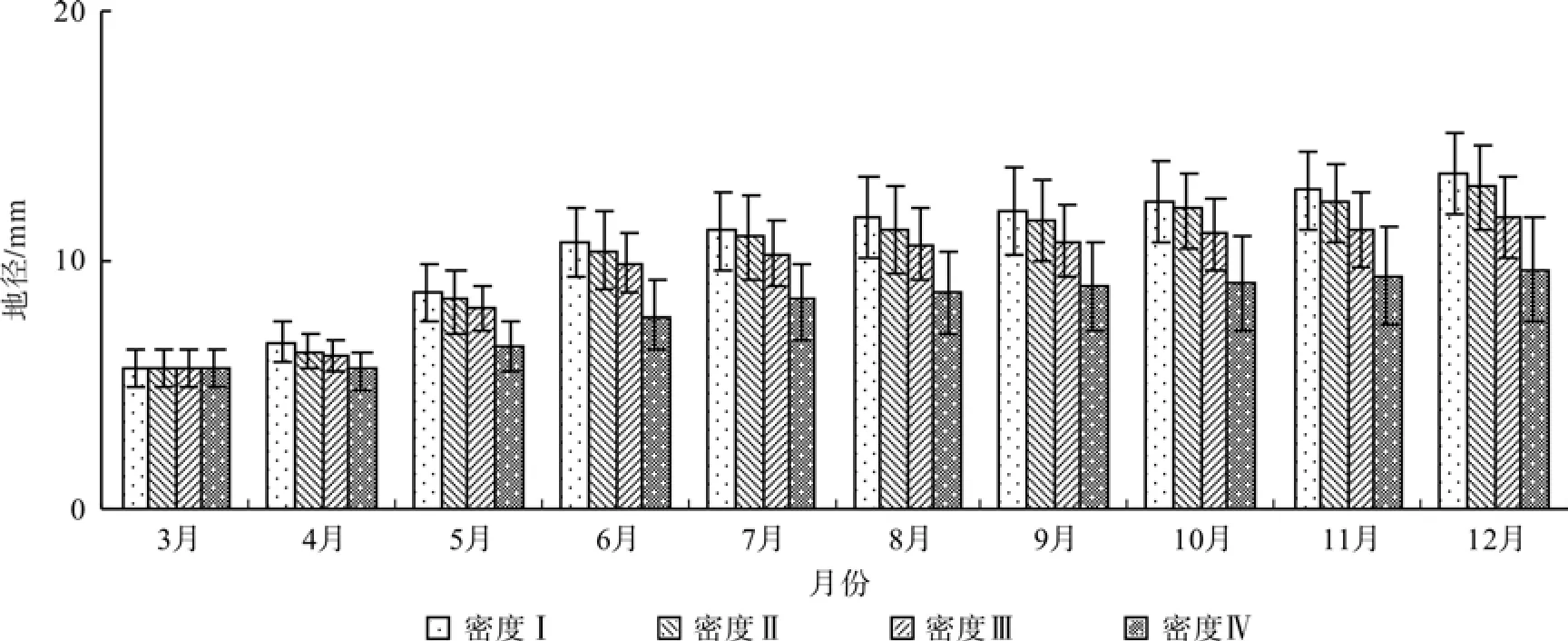
图1 不同密度幼苗的地径生长Fig. 1 Growth of ground diameter of seedlings with different densities
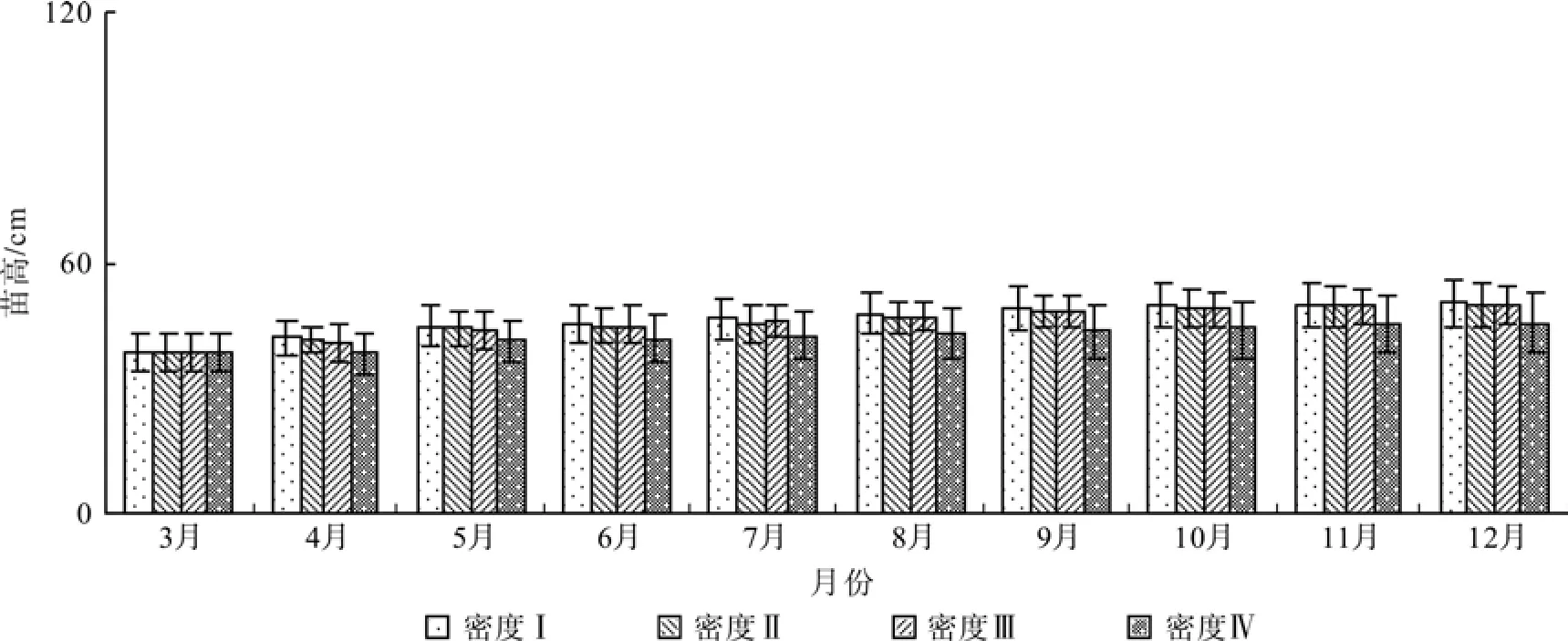
图2 不同密度幼苗的高生长Fig. 2 Growth of seedling height with different densities
2.2 各密度幼苗的单株生物量及其分配
幼苗各密度的干重(根、干、枝、叶)和总生物量均随着密度增大而减小(图3)。各密度的干重分配均表现为根>干>叶>枝(表2)。各密度的根干重比例为密度III >密度II >密度IV >密度I,密度I的叶干重分配比例较其他3个密度大。密度I、II、III和IV的每盆总干重分别为30.23、52.62、88.44、113.52 g,随密度增大而增加。根冠比为密度III >密度II >密度IV >密度I,均小于1。
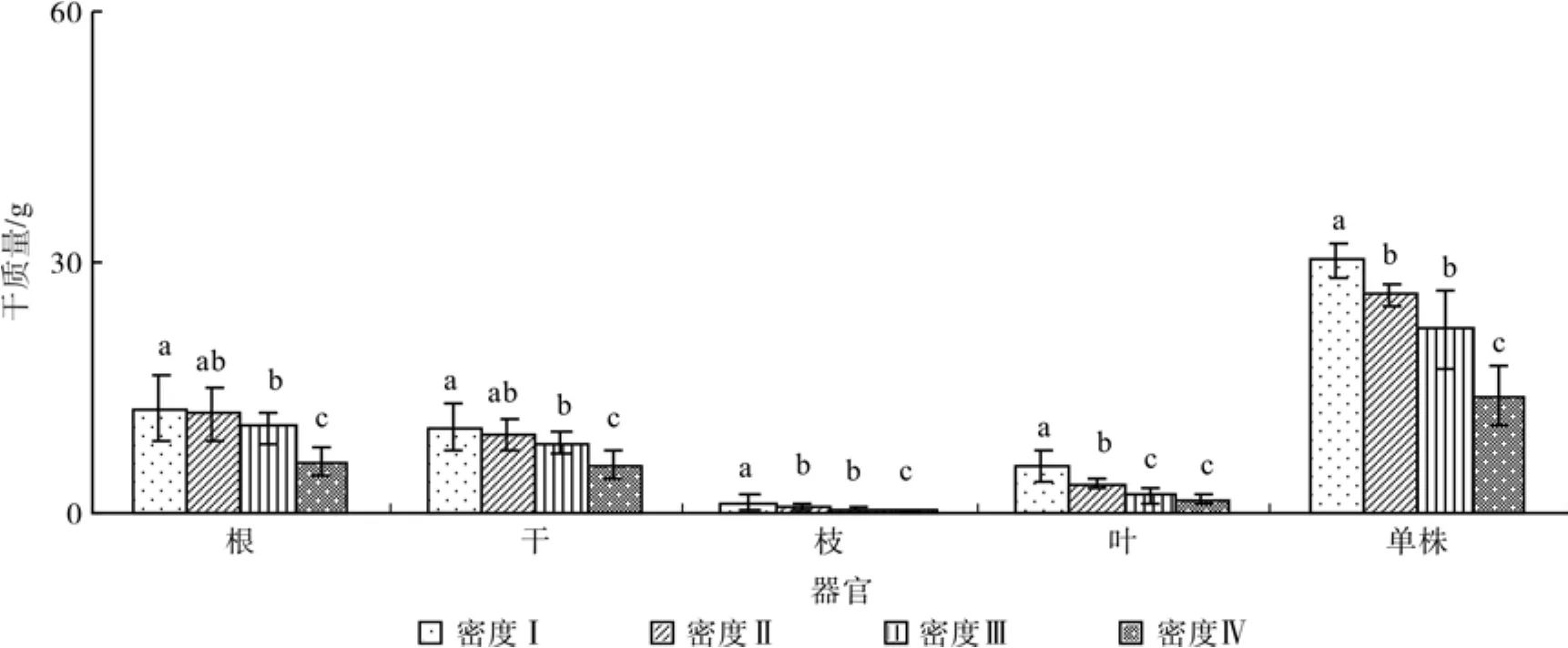
图3 不同密度单株器官干重和总干量Fig.3 Organ weight and tree weight of seedlings with different densities
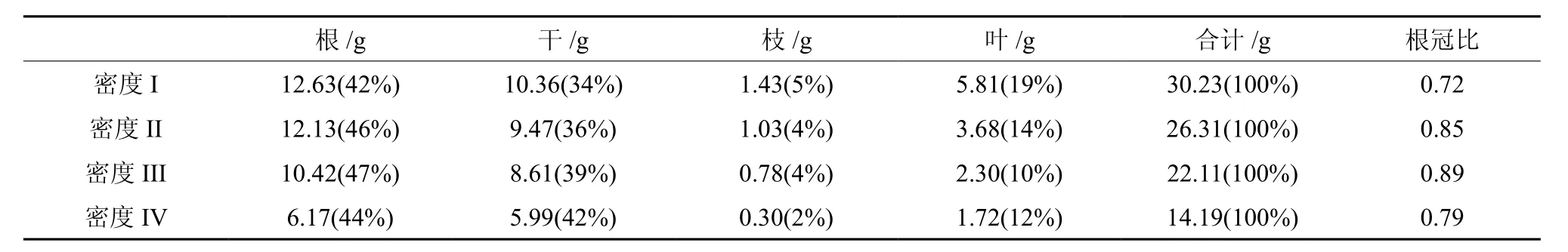
表 2 不同密度乐昌含笑的单株生物量分配及根冠比†Table 2 Biomass distribution and ratio of root to shoot of M. chapensis individual with different density
3 讨论
3.1 密度对幼苗形态生长的影响
密度影响着植物生长可利用的环境空间,密度越大,对光线、水分及营养的吸收利用越受到限制[18]。本研究中,乐昌含笑阔叶幼苗的地径随着密度的增加而减少。孙楠[19]对杂种落叶松进行研究,得出杂种落叶松的密度越大,其林分的平均胸径越小的结论。张春华等[20]对川楝Melia toosendan、新银合欢Leucaena leucocephala、苏门答腊金合欢Acacia suma等幼苗进行研究后发现,幼苗的地径都是随密度增大而减小。在乐昌含笑阔叶幼苗生长前期,密度间的地径差异较少,后期差异增大,可能是试验前期,密度大和密度小的植株间均有一定生长空间,可以收到足够的光照以满足幼苗光合作用,这导致生长前期的地径差异不大。生长后期,幼苗对光照和养分等资源的需求量大,高密度影响了幼苗个体获取的环境资源。随着幼苗的生长,密植的个体营养空间越小,幼苗间的竞争日益激烈,不利于生长发育。
本研究中,乐昌含笑的苗高随着密度的增加而减小,苗高的最高密度(I)仅为最小(IV)密度的1.1倍,标明该密度范围密度对乐昌含笑苗高的影响较小。我国一些学者对于杉木C. lanceolata[21]、桉树Eucalyptus robusta[6]等树种的研究也大多得出相同的结论。Xue et al.[9]的研究也发现密度对树高生长的影响小于胸径。
3.2 密度对幼苗的生物量及其分配影响
本研究中,各器官干重均随密度的增加而减少。这是由于密度小时,植物处于良好营养、水分、光照条件下,单株个体生物量大,然而随着密度的增加,植株平均营养面积缩小,单株个体生物量生长受到制约而与密度呈负相关关系[22]。安慧和上官周平[11]研究了密度对刺槐Robiniapseudoacacia幼苗生物量的影响,发现单株刺槐幼苗的根、茎、叶的生物量随着密度减小而呈增大趋势。赖文胜等[23]研究长序榆Ulmus elongata幼苗后发现,随着幼苗密度的增大, 苗木地上和地下部分重均有逐步减小的趋势。
乐昌含笑幼苗的根干重所占比例随着密度增大达到最大值(密度III最大),然后减小,其叶干重所占比例和根的规律相反。另外,该幼苗各密度的器官干重均为根>干>叶>枝。以上结果表明,密度会改变幼苗的生物量分配格局。1~4株/盆密度范围内,根干重所占比例增加,叶器官的减小,表明幼苗对地下资源的竞争较对激烈,密度会导致幼苗增加对根生长的投入。黄丽铭等[18]的研究也认为生物量会随密度的增大更多地分配到树干和树根。但密度增加到8株/盆时,其根比例反而减小,叶的增加,可能是由于密度过大,植株水平生长拥挤,对光资源的获取受到限制,所以增加对叶器官生物量的投入。密度会改变植物生物量的分配比例,但幼苗的生物量分配对密度程度的规律并不一致。乐昌含笑器官的生物量分配比例影响不大,各密度均为根>干>叶>枝。所以,密度对植物各器官生物量分配的影响因物种而异,一般被认为由基因型决定的[24]。
植物地下部分和地上部分生物量的大小反映了植物对土壤养分或光照的需求和竞争能力[25]。地下与地上生物量之比大于 1,说明植物对养分的需求和竞争能力强;反之其比值小于 1,说明植物对光照的需求和竞争能力强[26]。本研究中,乐昌含笑地下与地上生物量之比小于1,说明其植株对光照的需求和竞争能力较强。密度会改变幼苗地上与地下生物量的比值,但变化规律不明显,且各密度间的差异不大,可能是因为本研究的试验时间短所致。
[1] 薛 立, 傅静丹. 影响植物竞争的因子[J]. 中南林业科技大学学报, 2012,32(2): 6-15.
[2] 陈 伟, 薛 立. 根系间的相互作用—竞争与互利[J]. 生态学报, 2004, 24(6): 1243-1251.
[3] 杨元武, 王根轩, 李希来,等. 植物密度调控及其对环境变化响应的研究进展[J]. 生态学杂志, 2011, 30(8) : 1813-1821.
[4] 曾祥艳,陈金艳,廖健明,等. 不同栽培密度对多穗柯幼林生长的影响[J]. 经济林研究, 2014, 32(1): 113-116.
[5] 高 祥,丁贵杰,翟帅帅,等. 不同林分密度马尾松人工林根系生物量及空间分布研究[J]. 中南林业科技大学学报, 2014,34(6): 71-75.
[6] 黄宝灵, 吕成群, 蒙钰钗,等. 尾叶桉人工林种群密度的研究[J]. 应用生态学报, 2000, 11(1): 31-33.
[7] Xue L, Hagihara A. Growth analysis on the competition-density effect in Chinese fir (Cunninghamia lanceolata) and Masson pine(Pinus massoniana) stands[J]. Forest Ecology and Management,2001, 150: 331-337.
[8] Xue L, Hagihara A. Growth analysis on the C-D effect in selfthinning Masson pine (Pinus massoniana) stands[J]. Forest Ecology and Management, 2002, 165: 249-256.
[9] Xue L, Pan L, Zhang R,et al.Density effects on the growth of self-thinning Eucalyptus urophylla stands[J]. Trees,2011, 25(6):1021-1031. [6]
[10] Xue L, Jacobs D F, Zeng S C,et al. Relationship between aboveground biomass allocation and stand density index inPopulus×euramericanastands[J]. Forestry, 2012, 85(5): 611-619.
[11] 安 慧, 上官周平. 密度对刺槐幼苗生物量及异速生长模式的影响[J]. 林业科学, 2008, 44(3): 151-155.
[12] 姜景民, 滕花景, 袁金玲, 等. 乐昌含笑种群遗传多样性的研究[J]. 林业科学研究, 2005, 18(2):109-113.
[13] 田大伦, 尹刚强, 方 晰, 等. 湖南会同不同退耕还林模式初期碳密度、碳贮量及其空间分布特征[J]. 生态学报, 2010,30(22): 6297-6308.
[14] 许松葵,薛 立. 6种阔叶树种幼林的林地土壤特性[J]. 华南农业大学学报, 2010, 31 (4): 76-81.
[15] 郭淑红, 薛 立,张 柔,等. 4种幼苗对低温胁迫的光合生理响应[J]. 华南农业大学学报,2012, 33(3):373-377 .
[16] 叶龙华,杨振意,薛 立,等. 3种幼苗对水淹胁迫的生理响应[J].华南农业大学学报,2012,32(3):368-372.
[17] 薛 立,张 柔,奚如春,等. 华南地区6种阔叶幼苗叶片形态特征的季节变化[J]. 生态学报,2012, 32(1): 123-134 .
[18] 黄丽铭, 薛 立, 王相娥,等. 不同密度下大叶相思幼林的生长和生物量分配格局[J].华南农业大学学报,2008,29(3):52-55.[19] 孙 楠. 不同造林密度杂种落叶松人工林动态研究[D]. 东北林业大学, 2012.
[20] 张春华, 廖声熙, 李 昆,等. 金沙江干热河谷3个造林树种裸根 1 年生苗的育苗密度研究[J]. 西部林业科学, 2006,36(4): 80-86.
[21] 丁贵杰, 周政贤, 严仁发, 等. 造林密度对杉木生长进程及经济效果影响的研究[J]. 林业科学, 1997, 33(1): 67-75.
[22] 姚东和, 杨民胜, 李志辉. 林分密度对巨尾桉生物产量及生产力的影响[J]. 中南林学院学报,2000,20(3):20-23.
[23] 赖文胜,邹高顺,张纪卯. 不同密度对长序榆苗期生长的影响[J] 福建林业科技, 2001, 28(2): 74-76, 80.
[24] Weiner J. Allocation, plasticity and allometry in plants[J].Perspectives in Plant Ecology, Evolution and Systematics, 2004,6: 207-215.
[25] Aikio S, Rämö K, Mannin S. Dynamics of biomass partitioning in two competing meadow plant species[J]. Plant Ecology, 2009,205: 129-137.
[26] 张 莹. 不同育苗密度对千年桐幼苗养分吸收利用和生物量分配的影响[D]. 福建农林大学, 2011.
