太湖梅梁湾藻蓝蛋白转录间隔区基因和伪空胞基因表达及其影响因子*
2015-05-10张金丽GEXiuchun李玉燕
张金丽,虞 龙,GE Xiuchun,李玉燕,于 洋
(1:南京工业大学生物与制药工程学院,南京211816)
(2:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京210008)
(3:Virginia Commonwealth University, Richmond 23284)
太湖梅梁湾藻蓝蛋白转录间隔区基因和伪空胞基因表达及其影响因子*
张金丽1,2,虞 龙1,GE Xiuchun3,李玉燕1,于 洋2**
(1:南京工业大学生物与制药工程学院,南京211816)
(2:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京210008)
(3:Virginia Commonwealth University, Richmond 23284)
以室内铜绿微囊藻7806(Microcystisaeruginosa7806)为对照,运用反转录定量PCR技术研究太湖梅梁湾水体蓝藻藻蓝蛋白转录间隔区基因(PC-IGS)和伪空胞基因(gvpC)在2013年1月至2014年1月的相对表达量,并分析它们与环境因子的关系. 结果表明, PC-IGS相对表达量在2013年1—5月逐渐上升,并在5月达到最大值;gvpC相对表达量从2013年1月开始持续上升并在3月达到最大值;gvpC的表达早于PC-IGS. 相关分析表明,PC-IGS的相对表达量与硝态氮、溶解氧浓度均呈极显著正相关,与亚硝态氮、铵态氮以及正磷酸盐浓度均呈显著正相关,与pH值呈显著负相关,与温度、总氮和总磷浓度均无显著相关性;gvpC的相对表达量与硝态氮、铵态氮以及正磷酸盐浓度均呈极显著正相关,与总氮和亚硝态氮浓度呈显著正相关,与温度和pH值均呈显著负相关,与溶解氧和总磷浓度均无显著相关性.
藻蓝蛋白转录间隔区基因;伪空胞基因;反转录定量PCR;环境因子;相对表达量;太湖
湖泊富营养化和蓝藻水华的发生是目前全世界共同面临的重大环境问题之一[1],而在太湖和许多富营养化湖泊的藻类类群中,微囊藻(Microcystisspp.)是一个分布广泛的水华蓝藻种类[2]. 有研究指出蓝藻水华的形成存在3个基本要素:一是水体中蓝藻能在种间竞争中形成优势;二是蓝藻能够形成较大的生物量;三是具备合适的水力及水文气候条件,蓝藻能通过浮力调节在水体表层聚集[3]. 大量蓝藻群体通过调节自身浮力改变它们在水柱中所处的深度,以获取适宜的光能,得到充足的营养盐,这使得它们在与其它藻类竞争时具有明显的优势. 因此,研究蓝藻生物量以及浮力调控和垂直迁移特征对于探明水华形成机制有着十分重要的意义.
通常用蓝藻体内参与光合作用的藻蓝蛋白浓度表征蓝藻生物量,对于蓝藻上浮而言,藻细胞通过其细胞内结构——伪空胞的破裂与合成,调节自身在水体中的垂直分布. 然而富营养化湖泊水质背景复杂,传统的藻蓝蛋白分析方法(如分光光度法、高效液相色谱法等)及伪空胞分析方法(如压力毛细管法)不能区分死活细胞,难以真正反映具有活性藻类的状态. 随着分子生物学技术的发展,从环境样品中提取和处理核酸的技术逐渐被应用到微生物形态学中. 在研究微生物群落关键基因的动态变化时,以DNA为模板扩增目的基因序列往往会受到死亡细胞或游离DNA残体的影响,因而难以判断其生态学影响[4]. 而分析RNA序列则能针对微生物群落中具有活性的生物,尤其是mRNA表达的研究分析可以提供更多的针对特定微生物的信息. 目前发展起来的实时荧光定量反转录PCR技术能够很敏感地检测到含量很低的RNA,并对目的基因进行扩增定量化研究[5]. 根据设计特异性引物,可以对水体蓝藻藻蓝蛋白和伪空胞的组成蛋白在转录水平上进行研究,检测蓝藻在不同外部环境下数量和浮力的动态变化,为预防和控制蓝藻水华提供重要技术支持.
本研究通过对太湖梅梁湾野外水体蓝藻RNA进行提取,利用反转录和实时荧光定量PCR技术,分别以微囊藻藻蓝蛋白转录间隔区基因(PC-IGS)和伪空胞合成基因(gvpC)为研究对象,对其转录进行监测,探讨代谢因子之间的相互关系,并确定环境因子对PC-IGS和gvpC表达的影响.
1 材料与方法
1.1 采样点与样品采集
梅梁湾(31°25′53″N,120°12′42″E)位于太湖最北部,北接无锡市,是近年来太湖富营养化最为严重的湖区. 2013年1月至2014年1月(缺少2013年8月数据)每月采样. 用2.5 m长的PVC管采集整水柱,均匀混合. 同时用多功能水质参数仪(YSI6600-V2,Yellow Spring Instruments,USA)原位测定水体理化参数,包括溶解氧(DO)、电导率、浊度、水温、pH值等. 现场立即用GF/C滤膜过滤100ml水样用于RNA提取,然后将GF/C滤膜放入2ml冻存管中,立即向冻存管中加入一定体积的RNA later保护液,迅速将装有样品的2ml冻存管放入液氮中保存. 带回实验室放在-70℃中保存直到RNA提取.
1.2 营养盐的测定

1.3 藻蓝蛋白的测定
取水样100ml,用GF/C滤膜过滤,所得滤膜置于研钵中仔细研磨2~5 min,加入pH值为7.0 的Tris缓冲液6ml,然后转移到离心管中,4℃下避光保存10 h,然后在4000转/min下离心5 min,将上清液转移,定容. 藻蓝蛋白的测定用岛津分光光度计RF-5301,测定条件采用激发波长620 nm,发射波长647 nm,扫描波长设为60 nm/min,激发和发射光栅宽度均设为5 nm,响应时间2 s,增益置于normal,以0.05 mol/L pH值为7.0 的Tris缓冲液作为荧光参比液进行校零[7]. 每个样品重复3次.
1.4 水样中蓝藻RNA的提取
将2013年1月-2014年1月(缺少2013年8月数据)共12份保存在-70℃下的GF/C滤膜,分别转移到相对应编号的裂解管B(Lysing Matrix B,玻璃珠直径0.1 mm)中,然后每个裂解管B中加入500 μl Buffer RLT,将裂解管B放置于Fast Prep-24样品处理仪器中. 设置Fast Prep-24样品处理仪器参数为6 m/s,35 s. 击打破碎之后,立即取出裂解管B并放于冰上. 然后将裂解管B在4℃、12000 转/min条件下离心10 s,紧接着将上清液转移至无RNase的2ml离心管中. 由于裂解管B在离心之后很多未打碎的藻细胞下沉到管底部,再一次向裂解管B中加入500 μl Buffer RLT,再次打碎,离心. 将上清夜与第1次离心得到的上清液混合. 然后根据RNeasy Plant Mini Kit RNA提取试剂盒的操作说明书,并对试剂盒的操作步骤进行微小调整,在加入70%无水乙醇清洗之后,加入DNaseI去除基因组DNA. 将提取的无DNA污染的RNA在-70℃下贮存备用. 利用NanoDrop 2000/2000c微量紫外分光光度计测量RNA浓度和纯度(OD260/280).
1.5 反转录定量PCR
利用室内培养的铜绿微囊藻(Microcystisaeruginosa7806)作为对照,培养基为BG-11,培养温度为25℃,光照强度为40 μE/(m2·s),光照周期为12 h ∶12 h,离心收集处于对数生长期的藻细胞. 按照1.4节提取野外水体蓝藻RNA的方法提取室内微囊藻RNA.
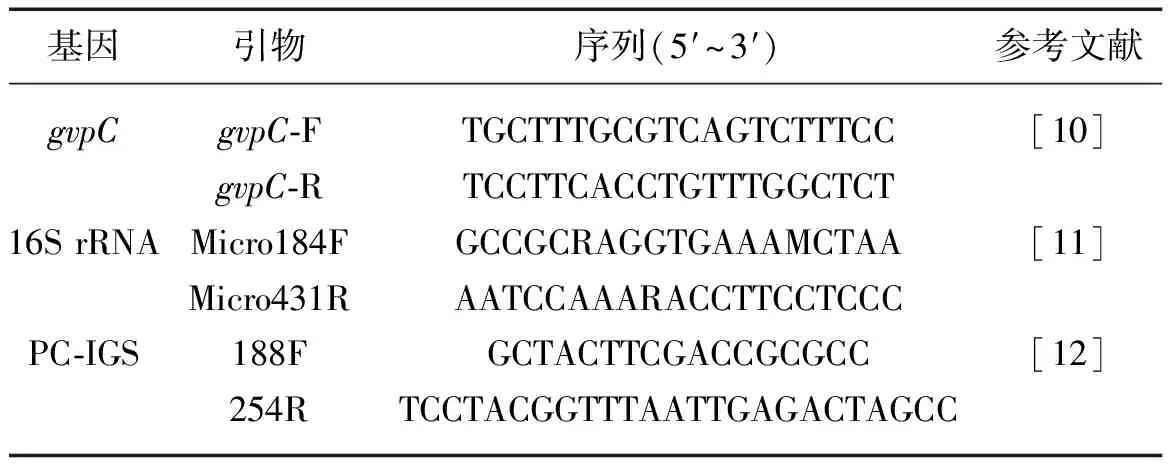
表1 引物序列
根据从不同样品中提取的RNA浓度,将RNA统一定量至400 ng,用Transciptor First Strand cDNA Synthesis Kit反转录试剂盒合成单链cDNA模板. 本实验选取微囊藻特异性16S rRNA基因为内参基因, PC-IGS和gvpC为定量检测目的基因. 针对各个目的基因所设计的引物见表1. 将以16S rRNA为引物的cDNA模板稀释10倍,其它的cDNA模板不稀释. qRT-RCR反应体系为25 μl,其中含12.5 μl 2×QuantiFast SYBR Green PCR Master Mix,2 μmol/L引物,1 μl模板,无RNA酶的水10.5 μl. PCR反应在replex4(Eppendorf)荧光定量PCR仪上完成. 扩增程序如下:95℃预变性3 min,95℃变性15 s,60℃退火30 s,72℃延伸45 s,40个循环. PCR完成后,导出相应的阈值循环数值(threshold cycle value,简写为Ct值,定义为荧光信号到达设定的阈值时所经历的循环数,目的基因的拷贝数与Ct值呈负相关). 以16S rRNA为阳性内对照基因来校正PCR模板的细胞拷贝数(ΔCt目的基因=Ct目的基因-Ct同一样本16S),从而消除组间加样量. 实验重复3次. 每个样品的ΔCt平均值减去室内ΔCt平均值得到每个目的基因相对循环数(ΔΔCt值),即ΔΔCt目的基因=野外ΔCt目的基因-室内ΔCt目的基因). 相对量(表达变化倍数)的计算采用2-ΔΔCt方法[8]. 最后进行熔解曲线分析,为了证明目的基因(PC-IGS和gvpC)和内参基因(16S RNA)的扩增效率(amplification efficiency)一致,将室内样品的cDNA模板稀释成一系列梯度,利用内参基因(16S rRNA)和目的基因(PC-IGS和gvpC)引物扩增,通过cDNA浓度梯度的lg值对ΔCt值(ΔCt=Ct目的基因-Ct内参基因)作图,如果所得直线斜率绝对值接近于0,说明扩增效率相同[9].
1.6 数据处理和分析
采用Origin 8.5和SPSS 16.0软件对数据进行处理和分析.
2 结果
2.1 太湖梅梁湾水体理化参数和营养盐的月变化

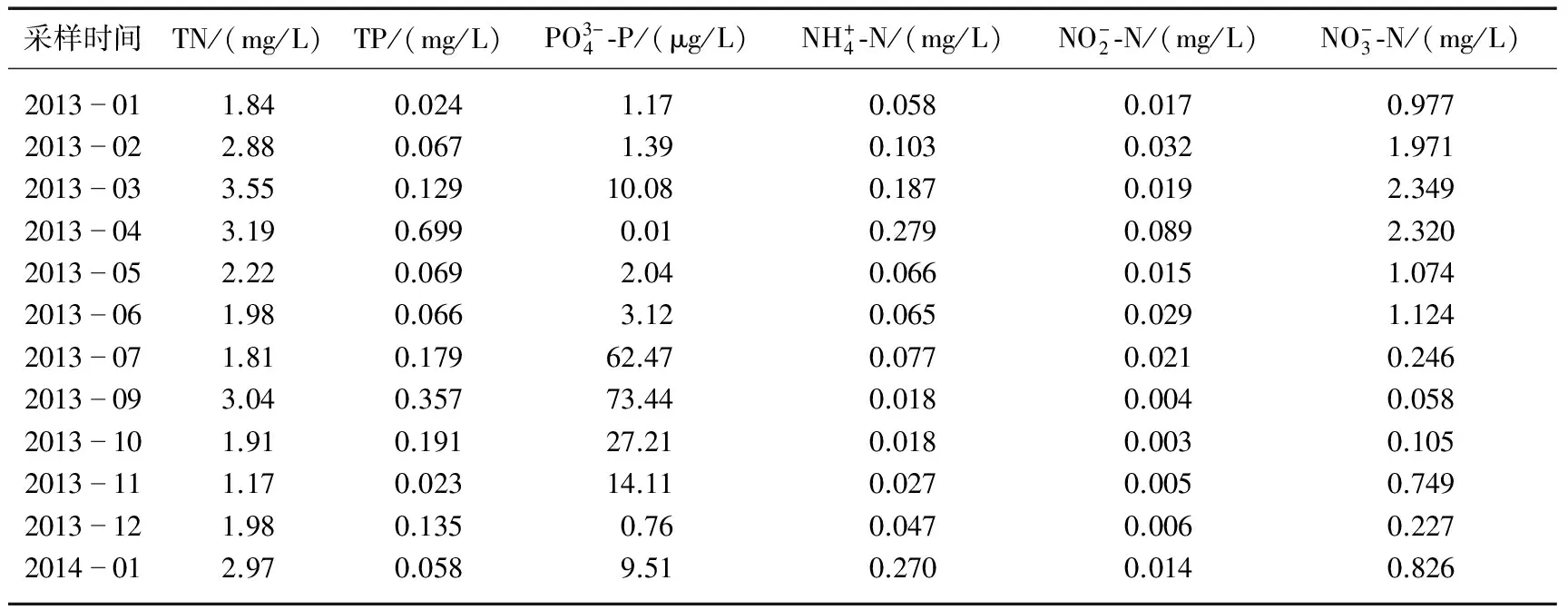
表2 2013年1月-2014年1月太湖梅梁湾水体的每月营养盐浓度
2.2 太湖梅梁湾水体中藻蓝蛋白浓度的月变化
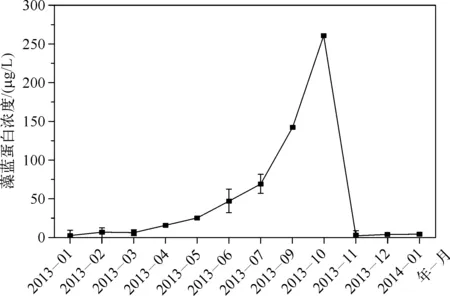
图1 2013年1月至2014年1月太湖梅梁湾水体藻蓝蛋白浓度Fig.1 Phycocyanin concentration of water in Meiliang Bay, Lake Taihu from January 2013 to January 2014
在2013年1—4月期间,太湖梅梁湾水体藻蓝蛋白浓度呈缓慢升高的趋势,而在5—10月藻蓝蛋白浓度大幅度升高,并在10月达到最大值(260.1 μg/L),之后,藻蓝蛋白浓度急剧下降并在11月达到全年最低值(2.9 μg/L),春季和冬季藻蓝蛋白浓度明显低于夏季和秋季(图1).
2.3 荧光定量反转录PCR测定
提取所有样品的RNA,OD260/280值均在1.8~2.2之间,说明提取的样品RNA纯度均符合要求. 目的基因PC-IGS和gvpC经过一系列梯度稀释的cDNA模板扩增后,通过cDNA浓度梯度的对数转化,对ΔCt值作图,得到的直线方程分别为y=-0.016x+10.023和y=-0.0931x+10.109. 两条直线斜率的绝对值都接近0,2个目的基因(PC-IGS和gvpC)和内参基因的扩增效率一致.
2.4 太湖梅梁湾水体中蓝藻PC-IGS相对表达量的月变化
根据实时荧光定量反转录PCR技术和2-ΔΔCt计算方法,得到2013年1月至2014年1月(缺少2013年8月数据)期间太湖梅梁湾水体中PC-IGS相对于室内微囊藻PC-IGS的相对表达量. 结果表明,在不同时期内PC-IGS相对表达量有很大差异,从1—5月藻蓝蛋白基因相对表达量逐渐上升,并在5月达到最高,5月的相对表达量是1月的7倍. 但到6月藻蓝蛋白基因相对表达量急剧下降至只有5月的1/4,和2月的相对表达量几乎相同. 9月是全年中藻蓝蛋白基因相对表达量最低的月份,从10月份开始藻蓝蛋白转录水平逐渐上升,总的来看冬季相对表达量始终低于春季(图2).
2.5 太湖梅梁湾水体中蓝藻gvpC相对表达量的月变化
2013年1—3月太湖梅梁湾水体中蓝藻gvpC相对表达量呈升高趋势,并在3月达到全年表达量的最高值. 1月和2月gvpC的相对表达量很接近,2—3月其相对表达量却急剧上升,3月相对表达量几乎是1、2月份的3倍,到4月时gvpC相对表达量直线下降到只有3月份的1/2. 5—10月这段时间内gvpC相对表达量呈下降趋势,并在9月和10月达到最低值,10月份的相对表达量几乎检测不到. 从结果还可以看出,gvpC相对表达量从10月就开始上升(图2).
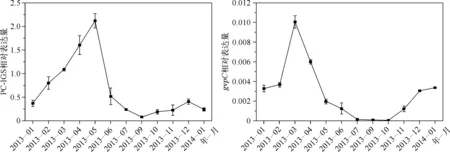
图2 2013年1月至2014年1月太湖梅梁湾水体中蓝藻PC-IGS和gvpC相对表达量的月变化Fig.2 Relative expressions of PC-IGS and gvpC in cyanobacteria in Meiliang Bay, Lake Taihu from January 2013 to January 2014
2.6 相关性分析
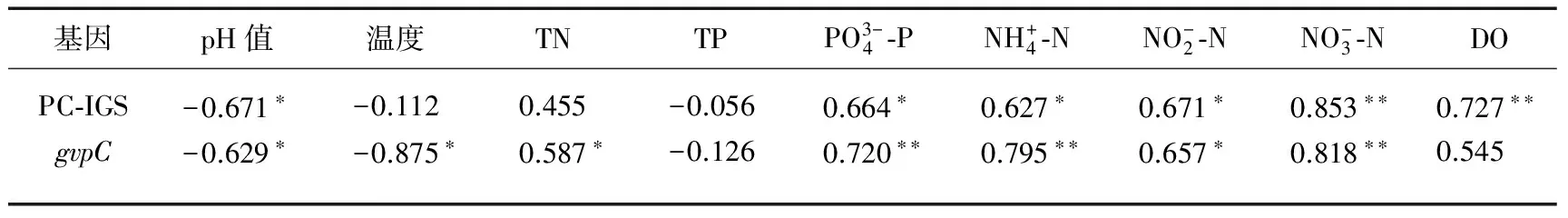
表3 环境因子与PC-IGS以及gvpC基因相对表达量的相关性分析
*表示P<0.05,**表示P<0.01.
3 讨论
荧光定量PCR技术在蓝藻水华研究中的应用已相当广泛. 目前,基于目标DNA片段的定量PCR技术已经成熟而且被广泛应用于估算野外环境蓝藻丰度和产毒微囊藻丰度[13-14],这项技术能够快速确定蓝藻水华毒性,并对蓝藻水华的生态风险做出评价. 而本研究运用反转录定量PCR技术检测了太湖梅梁湾水体蓝藻PC-IGS和gvpC在2013年1月至2014年1月不同时间内的相对表达量,在转录水平上检测蓝藻在不同外部环境下数量和浮力的动态变化. 在实时荧光定量PCR结果分析中采用2-ΔΔCt方法分析基因表达的相对变化,它的优点是无须知道起始模板的拷贝数,只须保证PCR扩增目标基因和内参基因的扩增效率相同,分析基因相对表达量即可. 在本实验中将室内样品的cDNA模板稀释成一系列梯度,利用内参基因(16S rRNA)和目的基因(PC-IGS和gvpC)引物扩增,通过cDNA浓度梯度的对数值对ΔCt值构造曲线,所得到的2条直线斜率的绝对值(0.016和0.0931)都接近于0,说明2个目的基因(PC-IGS和gvpC)和内参基因的扩增效率一致,从而保证了实验结果的准确性.
结合2013年1月到2014年1月藻蓝蛋白浓度的月变化以及蓝藻生活史来看,11月到次年2月是蓝藻整个生活史中一个非常特殊的时期——越冬期,藻体经历了大量消亡后逐渐下沉,所以导致水体中蓝藻数量逐渐减少,并一直处于低位. 随着春季(3—5月)温度上升、光照加强,在冬季下沉到湖泊底泥以及上覆水中的水华蓝藻细胞及群体开始进入水柱,逐渐复苏生长,所以看到2013年3—5月梅梁湾水体蓝藻数量逐渐增加. 与阎荣等[15]和吴晓东等[16]对太湖、巢湖和玄武湖等湖泊的周年野外观测研究和室内模拟实验发现的结果略有不同:他们指出在秋季,随着温度的降低,不同湖泊及不同湖区的水体中表征蓝藻生物量的藻蓝蛋白浓度逐渐下降. 而本研究结果发现梅梁湾水体藻蓝蛋白浓度从5月份逐渐升高并持续到10月,这可能是由于2013年夏、秋季的温度高于往年同期,导致蓝藻水华完全消失的时间推迟.
从基因表达水平上看,PC-IGS和gvpC的表达在时间上存在差异,gvpC的表达早于PC-IGS,可能是由于随着gvpC表达量的升高,蓝藻迅速合成伪空胞,从而带动蓝藻从水体上浮到水体表面,进而使得蓝藻获得适宜的生长条件和营养盐,这时藻蓝蛋白基因才开始迅速表达. 在蓝藻水华形成的各个阶段,伪空胞是否一直为蓝藻提供浮力并起到决定性作用并不清楚,本研究发现蓝藻在越冬复苏阶段,gvpC转录水平逐渐升高并在3月份达到最大值,但是在蓝藻生物量增加以及上浮阶段,gvpC表达水平逐渐降低. 这表明gvpC的表达及伪空胞的形成极有可能是微囊藻摆脱休眠状态、重新获得浮力的一个标志性转变,当藻细胞重新恢复成正常细胞后即不需要gvpC的持续表达. 这一结果与张永生等[17]的室内研究结果一致:在微囊藻上浮过程中,伪空胞的气体相对含量先增加后减少,gvpC基因转录水平也随着微囊藻上浮逐渐降低,他的研究还表明伪空胞是微囊藻上浮的主要浮力提供者,该浮力仅能使微囊藻悬浮在水中,不能使其漂浮到水面形成蓝藻水华. 与gvpC表达不同,PC-IGS相对表达量从1—5月逐渐升高,并在5月迅速达到最大值,贯穿整个越冬复苏期. 当形成水华后,PC-IGS的表达才逐渐下降. 这可能是由于当微囊藻进入水华期后,可以上浮至湖面获得最大光照,无法继续合成藻蓝蛋白. 而水体藻蓝蛋白浓度却从3月逐渐增加并持续到10月,主要是因为微囊藻的复苏可能是一个渐进的过程,加之底泥表面的微囊藻种源不断进入水体,上浮至水体表面,直至盛夏藻浓度才到达最高值.

致谢:葛秀春、余丽、杜明勇等对本实验提供指导与帮助,特此致谢!
[1] 孔繁翔, 马荣华, 高俊峰等. 太湖蓝藻水华的预防、预测和预警的理论与实践. 湖泊科学, 2009, 21(3): 314-328.DOI 10.18307/2009.0302.
[2] 孙慧群, 朱 琳, 高文宝. 淡水湖泊中微囊藻水华的成因分析. 生物学通报, 2005, 40(8): 23-24.
[3] 杨 波. 蓝藻伪空胞的特征及其浮力对氮、磷和温度的响应机制[学位论文]. 长沙: 湖南农业大学, 2007: 12-13.
[4] Miskin IP, Farrimond P, Head IM. Identification of novel bacterial Lineages as active members of microbial populations in a freshwater sediment using a rapid RNA extraction procedure and RT-PCR.Microbiology, 1999, 145(8):1977-1987.
[5] Saleh-Lakha S, Miller M, Campbell RGetal. Microbial gene expression in soil: methods, applications and challenges.JournalofMicrobiologicalMethods, 2005, 63(1): 1-19.
[6] 金相灿, 屠清瑛. 湖泊富营养化调查规范. 北京:中国环境科学出版社, 1990: 160-185.
[7] 季 健, 孔繁翔, 于 洋等. 太湖越冬蓝藻空间分布的初步研究.湖泊科学, 2009, 21(4): 490-494.DOI 10.18307/2009.0406.
[8] 王秀敏, 邵敏华, 卢大儒等. 应用高通量实时荧光定量PCR方法筛选FL细胞对低剂量微囊藻毒素LR暴露的应答基因. 水生生物学报, 2007, 31(6): 909-911.
[9] Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCtmethod.Method, 2001, 25(4): 402-408.
[10] Mlouka A, Comte K, Casters AMetal. The gas vesicle gene cluster fromMicrocystisaeruginosaand DNA rearrangements that lead to loss of cell buoyancy.JournalofBacteriology, 2004, 186(8): 2355-2365.
[11] Rinta-Kanto JM, Ouellette AJA, Boyer GLetal. Quantification of toxicMicrocystisspp. during the 2003 and 2004 blooms in the western Lake Erie using quantitative real-time PCR.EnvironmentalScienceandTechnology, 2005, 39(11): 4198-4205.
[12] Kurmayer R, Kutzenberger T. Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacteriumMicrocystissp.AppliedandEnvironmentalMicrobiology, 2015, 81(12): 6723-6730.
[13] Pearson LA, Neilan BA. The molecular genetics of cyanobacterial toxicity as a basis for monitoring water quality and public health risk.CurrentOpinioninBiotechnology, 2008, 19(3): 281-288.
[14] Martins A, Vasconcelos V. Use of qPCR for the study of hepatotoxic cyanobacteria.ArchivesofMicrobiology, 2011, 193(9): 615-627.
[15] 阎 荣, 孔繁翔, 韩小波. 太湖底泥表层越冬藻类群落动态的荧光分析法初步研究. 湖泊科学, 2004, 16(2):163-168.DOI 10.18307/2004.0204.
[16] 吴晓东, 孔繁翔, 张晓峰等. 太湖与巢湖水华蓝藻越冬和春季复苏的比较研究. 环境科学, 2008, 29(5):162-167.
[17] 张永生, 李海英, 孔繁翔等. 群体细胞间空隙在微囊藻水华形成过程中的浮力调节作用. 环境科学, 2011, 32(6):1602-1607.
[18] 许 敏. 微囊藻伪空胞基因丛的研究[学位论文]. 武汉: 中国科学院水生生物研究所, 2006: 49-52.
[19] 周 贝, 毕永红, 胡征宇. 温度对铜绿微囊藻细胞浮力的调控机制.中国环境科学, 2014, 34(7):1847-1854.
[20] Damerval T, Xastets AM, Houmard Jetal.Pseudanabaenasp: Occurrence of a single photoregulated gene.Molmicrobial, 1991, 5: 657-664.
[21] Parrish J. The role of nitrogen and phosphorus in the growth, toxicity, and distribution of the toxic Cyanobacteria,Microcystisaeruginosa. San Francisco: University of San Francisco Scholarship Repository, 2014: 8.
Environment factors and expression of PC-IGS andgvpCgenes in Meiliang Bay of Lake Taihu
ZHANG Jinli1,2, YU Long1, GE Xiuchun3, LI Yuyan1& YU Yang2
(1:CollegeofBiotechnologyandPharmaceuticalEngineering,NanjingUniversityofTechnology,Nanjing211816,P.R.China)
(2:StateKeyLaboratoryofLakeScienceandEnvironment,NanjingInstituteofGeographyandLimnology,ChineseAcademyofSciences,Nanjing210008,P.R.China)
(3:VirginiaCommonwealthUniversity,Richmond23284,USA)
The relative expression of intergenic spacer region-within the Phycocyanin(PC-IGS) and gas vesicle gene(gvpC) in cyanobacteria in Meiliang Bay, Lake Taihu from January of 2013 to January of 2014, was investigated using reverse transcription-quantitative PCR, compared to those in interiorMicrocystisaeruginosa. In addition, we analyzed the influence of environmental factors on the relative expressions of PC-IGS andgvpCgene. The results showed that the relative expression of PC-IGS increased from January to May of 2013, and reached the maximum in May of 2013. The relative expression ofgvpCgene rose from January of 2013, and reached the maximum in March of 2013. The expression ofgvpCgene increased earlier than that of PC-IGS. The data also showed that the relative expression of target gene PC-IGS was correlated significantly and positively with the concentrations of nitrate nitrogen, dissolved oxygen, ammonia, nitrite nitrogen and orthophosphate salt, negatively with pH value, and not significantly with temperature, total nitrogen and total phosphorus. The relative expression of target genegvpCwas correlated significantly and positively with the concentrations of ammonia, nitrate nitrogen and orthophosphate salt. They are positively correlated with the concentrations of total nitrogen and nitrite nitrogen, but negatively with pH value and temperature, and not significantly with the concentrations of dissolved oxygen and total phosphorus in water.
PC-IGS;gvpC; reverse transcription-quantitative PCR; environmental factors; relative expression; Lake Taihu
*国家水体污染控制与治理科技重大专项(2012ZX07506-001)和太湖流域(江苏)水生态功能分区与标准管理工程建设课题(2011ZX07534-001)联合资助. 2014-09-11收稿;2014-12-12收修改稿. 张金丽(1989~),女,硕士研究生;E-mail:18761681572@163.com.
J.LakeSci.(湖泊科学), 2015, 27(5): 895-901
DOI 10.18307/2015.0516
©2015 byJournalofLakeSciences
**通信作者;E-mail:lzsswgc@163.com.
