基于粪便纤维组织法探究异地驯养藏羚的食性
2015-04-19郭玉萍熊建利张纪亮王占彬赵新全
孙 平,郭玉萍,熊建利,张纪亮,王占彬,赵新全
(1.河南科技大学 动物科技学院,河南 洛阳 471003;2. 安阳师范学院 体育学院,河南 安阳 455000;3. 中国科学院 西北高原生物研究所,青海 西宁 810001)
基于粪便纤维组织法探究异地驯养藏羚的食性
孙 平1,郭玉萍2,熊建利1,张纪亮1,王占彬1,赵新全3*
(1.河南科技大学 动物科技学院,河南 洛阳 471003;2. 安阳师范学院 体育学院,河南 安阳 455000;3. 中国科学院 西北高原生物研究所,青海 西宁 810001)
基于粪便纤维组织法探讨异地(铁卜加草原站)驯养藏羚的食性。结果表明,在基地生长的50种植物中,伊凡苔草、火绒草、川青早熟禾、洽草等是藏羚最喜爱食用植物,所占比例依次为18%、13%、11%和8%,其中伊凡苔草的茎、火绒草的叶、菥冥的果和迷果芹的花是其喜食部位。在基地驯养藏羚的食物组成中,各科所占比例依次为:禾本科(43%)> 莎草科(25%)> 菊科(14%)> 十字花科(7%)> 豆科(3%)。与可可西里地区类似,铁卜加地区藏羚以单子叶植物为主要食物。
藏羚;粪便纤维组织法;异地保护;食性
草食动物是影响植物群落组成和生态系统稳定的重要决定者。植食性哺乳动物食物选择是觅食生态学研究最为活跃的领域,动物食物选择对策主要有营养选择、植物次生化合物假设、营养平衡假设、最优觅食理论和条件性气味回避假设[1]。我国有蹄类食性研究在1978年后才有了长足进展,1979年郑生武等[2-5]采用直接观察法对马麝进行了试验,最近几年更是取得了重大突破,方法也日趋先进,通常主要用直接观察法、利用法、胃内容物分析法和粪便显微分析法。随着分子生物学技术的发展,粪便DNA分析法、稳定性同位素法[2]等更准确的测定方法相继出现。粪便显微组织法因为取样没有伤害性,成为近年来国内研究有蹄类食性的最主要方法[3-5]。
藏羚(Pantholops hodgsoni)为我国特有珍稀濒危动物,是青藏高原特有物种,野生动物中最独特的类群,同时也是世界上最珍贵和稀有的物种之一[6]。藏羚主要分布在我国西藏、青海、新疆西部近80万平方公里的高寒荒漠地区,其中80%区域及其70%种群数量在西藏羌塘境内。近年来由于遭到疯狂盗猎,藏羚成为濒危动物,已被列入中国一级重点保护野生动物和《濒危野生动植物种国际贸易公约》附录I中[7]。
目前,关于藏羚的研究涵盖其生物学特征[8]、种群生态学[6, 7, 9-15]、保护生物学[16-20]和行为学[15, 21-24]等方面。而动物的食性分析是研究动物生境需求的核心内容。基于此,有学者运用粪便显微组织法对可可西里自然保护区藏羚的食性进行研究,并从营养学角度探讨了它们之间竞争与共存的关系[25]。还有学者对铁卜加地区驯养藏羚在枯草期对干物质消化率进行了研究[26]。另外,Harris & Miller[27]以及Schaller[15]分别对青海野牛沟地区和西藏羌塘地区藏羚的食性进行了研究,其食性分析仅做到了科的分类,但对异地驯养藏羚的食物构成至今未见报道。因此,本研究主要探讨异地驯养藏羚食物结构的适应性变化,为该物种异地保护时环境的选择提供科学参考。
1 材料与方法
1.1 研究样地
捕自可可西里的藏羚被安置在青海省铁卜加草原站。该区位于东经99°35′,北纬37°05′,海拔3 240~3 270 m,地处青藏高原北部、青海湖西北侧,是青海省海南、海北、海西三州交界处,属典型的高原半干旱草原区,年降水量377.10 mm,年蒸发量1 484.5 mm,年均气温-0.5 ℃,牧草生长期125 d,牧草生长季日照时数1 100 h (5~9月)。草地类型为草原化草甸类,草地植物主要有早熟禾(Popalpigenalindm)、针茅(Stipaspp)、赖草(Leymussecalinus)、猪毛蒿(Sabrotanoidesbunge)、萎陵菜(Potentillaspp)、狼毒(Stellerachamaejasme)等[28]。该区夏秋凉爽微润,冬春干冷多风,昼夜温差较大,霜期长。自然环境和气候条件独特,具青藏高寒牧区典型特征。自2005年12月,在该牧场提供的30 hm2围栏高寒草原化草甸驯养基地进行藏羚人工驯养[26]。
1.2 试验方法
1.2.1 藏羚粪便及参照植物样本采集 2006年9月,在铁卜加草原站藏羚驯养基地收集新鲜粪便67份,每一堆粪样为一份,单独装入塑料袋并标记取样时间、地点等信息,车载冰箱带回备用。同时采集其活动区域内所有植物完整植株,记录相关信息带回实验室,用于制作藏羚粪样显微分析的标准切片。
1.2.2 粪样植物碎片制备、装片和识别 植物标准玻片的制备方法主要参照Anthony(1974)[29],将植物阴干后,分别将每种植物的花、茎、叶等粉碎,放入试管中。然后加入5%硝酸和5%铬酸(1∶1),酒精灯加热煮沸约3 min,倒入200目网筛中冲洗,将筛上物用水冲入平皿中。用吸管将碎片滴入载玻片中央并覆以盖玻片。制作的标准玻片均在生物数码显微镜的20倍物镜下观察和拍照,并对植物表皮细胞进行测量,包括表皮细胞的形状(细胞的大小、密度及细胞壁的特征),气孔(气孔器的类型、数量和大小),表皮毛(单一细胞毛、多列细胞毛或腺毛的形状,长度和密度)。表皮细胞的性状特征及鉴别特征参照相关文献的描述[30-32]。
藏羚新鲜粪便在60 ℃烘干24 h,取粪便2 g经粉碎机粉碎,将粉碎的材料依次经20目、60目、100目网筛筛选,使碎片大小介于0.15 mm和0.30 mm之间。取100目的筛上物经自来水冲洗约2 min,移入平皿,加入次氯酸钠并用解剖针搅拌,3 h后倒入200目的网筛中自来水冲洗3 min,去除漂白液、尘土或极小的植物碎片,作为待检材料。
显微组织分析法的粪样植物碎片装片制作方法参考曹伊凡和苏建平等[5]。利用粪便显微分析法,在九月份所收集的全部藏羚羊粪样中,随机选取编号为06、08、10、14、15、22、26、27、28、33、34、35、39、44、50等共15份样品,共拍摄粪样图片1312张,共镜检到21科50种(属)植物碎片。
1.2.3 藏羚羊食物中植物种类的相对密度 采用下式计算粪便中各植物种类的相对密度(relative density, RD),RDi可作为植物的被利用频率。
RDi =(每种植物可辨认的碎片数 / 各种植物可辨认的碎片数量之和)× 100%。
式中:i表示物种种类。在九月份所收集的全部藏羚粪样中随机选取的样品中,先计算出编号06、08、10、14、15、22、26、27、28、33、34、35、39、44、50等样品中每种植物可辨认的碎片数,再将各种植物可辨认的碎片数相加,最后依据上述公式计算出50种植物中每种植物所占的相对密度,即各种植物的RD值。
2 结果与分析
2.1 藏羚的食物组成
在青海湖铁卜加藏羚羊驯养基地(以下简称"基地"),共采集21科50种植物(表1)。其中禾本科9种,莎草科2种,紫草科2种,菊科7种,豆科5种,蔷薇科3种,玄参科3种,毛茛科2种,伞形科2种,十字花科3种,唇形科2种,石竹科1种,百合科1种,龙胆科1种,鸢尾科1种,蓼科1种,瑞香科1种,茜草科1种,车前科1种,藜科1种,麻黄科1种。其中,禾本科占全部镜检植物的43%,其次为莎草科,占25%,再次为菊科、十字花科及豆科等植物,分别占14%、7%、3%。上述5科植物共占全部粪样可识别植物的 92%。
2.2 藏羚羊食物中植物种类的利用程度
由表2知,禾本科、莎草科以及菊科、十字花科、豆科等5科植物为藏羚羊的主要食物类群。在50种植物中伊凡苔草(Carextristachya)、火绒草(Leontopodiumleontopodioides)、川青早熟禾(Poaindattenuata)分别占镜检植物的18%、13%及11%,藏羚最喜食食物,其次为洽草8%(koeleriacristata)、菥冥7%(ThlaspiarvenseL)、矮嵩草7%(Kobresiahumilis)、冰草6%(AgropyroncristatumLinn)和赖草6%(LeymussecalinusGeorgi)。

表1 藏羚羊在九月份的食物谱
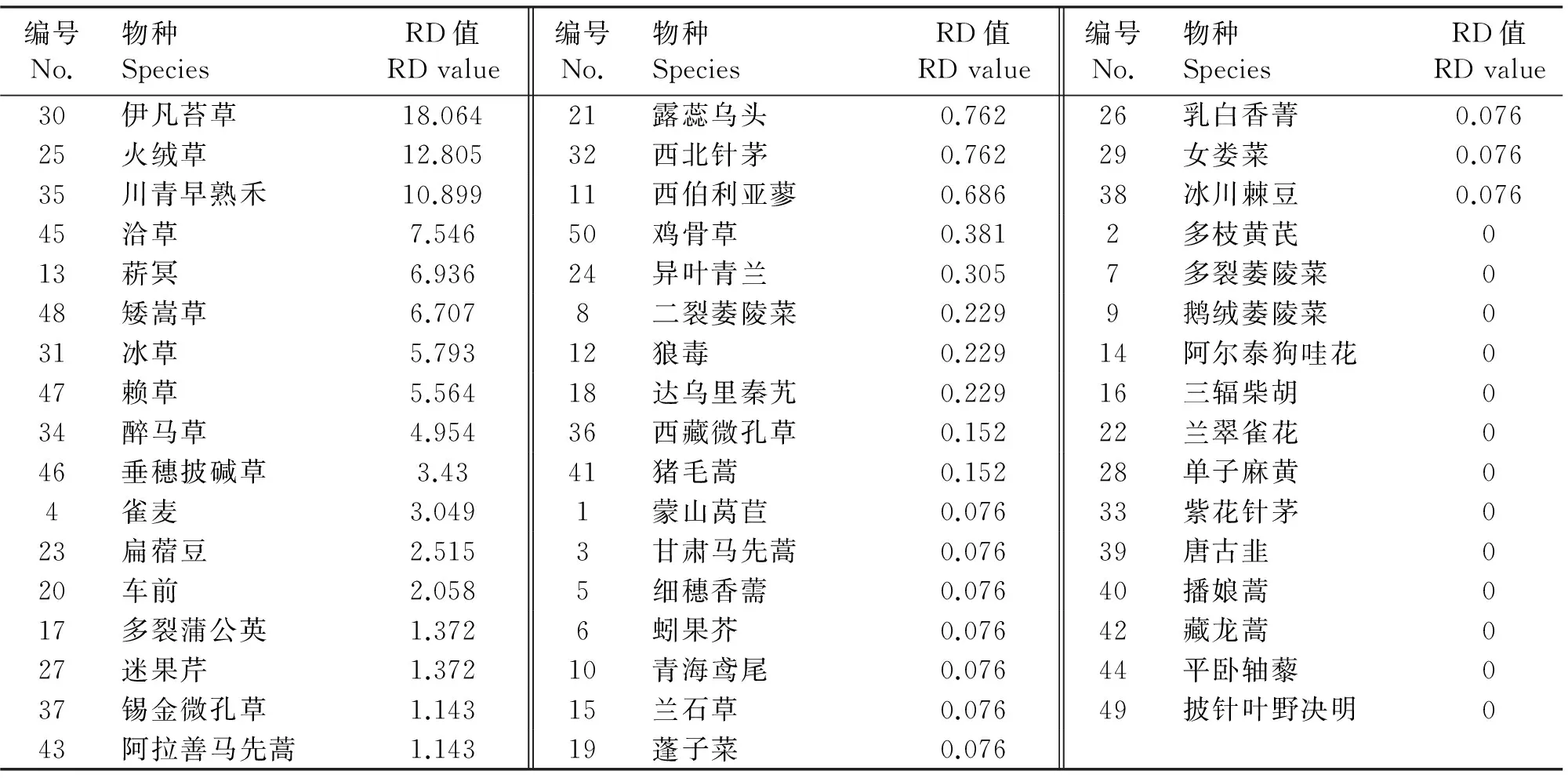
表2 粪样中各植物种类的相对密度
注:小数点后保留三位有效数字,物种名前的数字表示在九月份收集的标本编号。
Note:Three valid decimal numbers are kept,and numbers before speciecs are sample codes marked in september.
由表3可知,雀麦、菥冥、多裂蒲公英、车前、火绒草、迷果芹、伊凡苔草、矮嵩草、醉马草、川青早熟禾、洽草、垂穗披碱草、赖草、冰草等14种的RD值共计90.55,由此可见,这14种植物是其主要食物,其中伊凡苔草的茎为最高比例,达到34.2%;火绒草叶的比例最高,达到41.7%;菥冥的果为最高比例,达到44.3%;迷果芹的花为最高比例,达到14.3%(见表3)。
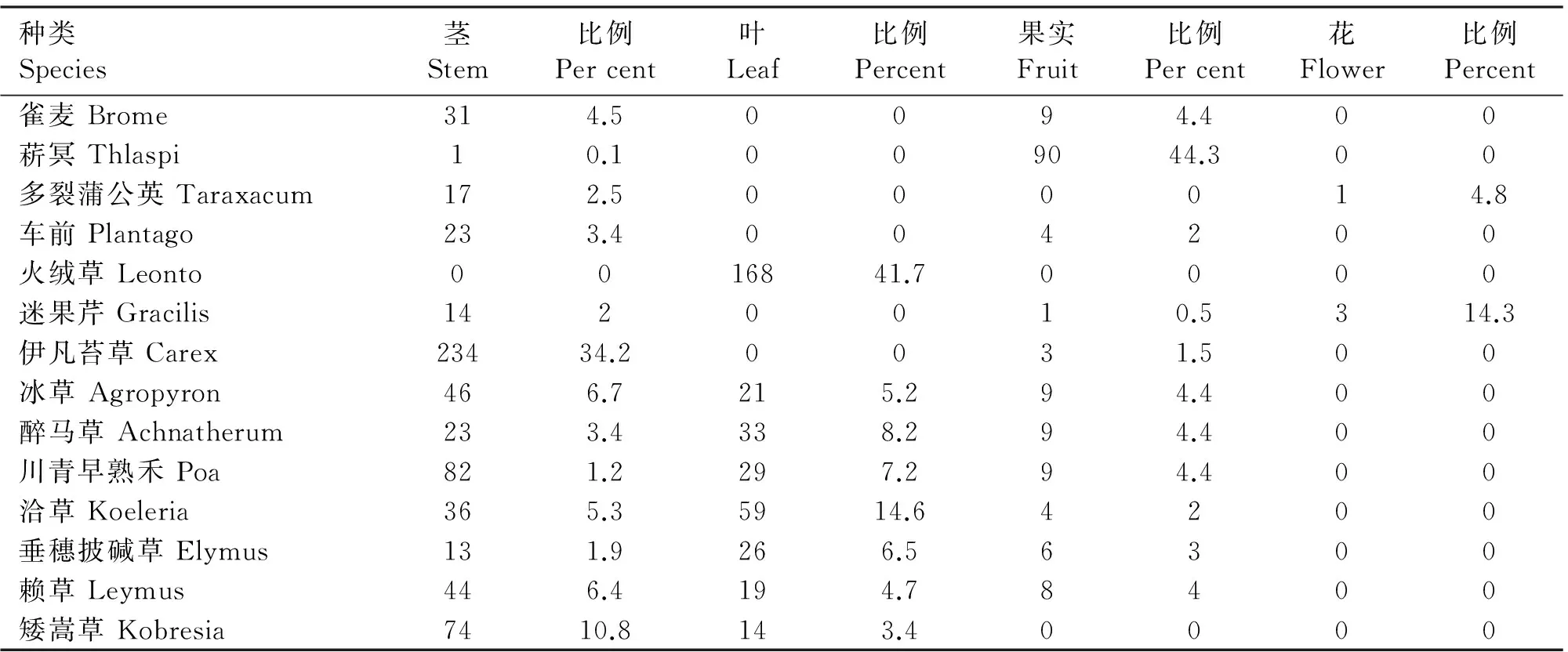
表3 14种植物的茎、叶、果、花相对比例
3 讨 论
动物与植物界面上的采食活动是草地放牧系统的核心特征。动物的食性分析是研究动物生境需求的核心内容,有蹄类动物食性的研究是一项基础生态学研究,它可以对栖息地评价、容纳量估计、能量代谢以及种间关系等研究提供有价值的基础资料,同时也为有蹄类动物尤其是濒危物种的异地保护、人工养殖和资源管理等提供理论依据,直接关系到该类动物资源科学管理水平[1, 3, 33-35]。
有蹄类动物的采食对策包括对食物种类、采食时间和采食地的选择,而这种对策的形成是长期进化、适应环境的结果[36]。异地驯养藏羚在受到气候因子影响的同时,还要受食物资源可利用性改变的制约,而且受异地物种之间的相互作用关系,尤其是异地物种之间的竞争作用影响。本研究主要分析了异地驯养藏羚的食性,结果表明,基地内藏羚的食物主要由禾本科、莎草科、菊科、十字花科和豆科等植物组成;其中,禾本科是其最主要的食物类群,占据了整个食物资源的43%。Schalle等[15]对西藏羌塘地区藏羚食性依据其粪便显微组织和胃内容物的研究表明,藏羚食性主要由禾本科、非禾本科和灌木组成,其中禾本科植物在暖季和冷季分别占其食物构成的40%和50%。曹伊凡等[4]人在可可西里地区分别于 2005年1月和7月在藏羚羊集聚地点随机收集其新鲜粪样,暖季藏羚羊粪样中镜检到15科24种(属)植物碎片,冷季镜检到 10科 18种 (属),且各科百分比依冷暖季的不同而不同。以上研究结果与本试验非常相似,即藏羚的食物组成中:禾本科(43%)> 莎草科(25%)> 菊科(14%)> 十字花科(7%) > 豆科(3%)。
在本试验的结果中,伊凡苔草、雀麦、菥冥、迷果芹、醉马草、川青早熟禾、垂穗披碱草、矮嵩草、赖草、冰草、洽草、车前、火绒草、多裂蒲公英等占了食物组成的近90%,以禾本科、莎草科为主,这和可可西里地区的藏羚羊的主要食物基本一致,不同之处在于多了十字花科的植物,这可能与海拔的降低和温度的升高有关,而且相同的季节由于海拔的影响,植物生长以及分布情况也对异地驯养藏羚造成一定的影响。藏羚食物种类的多寡以及采食比例的变化则可能与植物的营养元素成分、次生化合物含量和适口性有关[37]。
本研究采用粪便显微组织法分析了藏羚食性。该方法是食草动物食性研究中的常规方法,具有易于取样和对动物干扰小的优点,在食草动物食性研究中应用广泛。基本原理是根据粪便中未被消化的植物角质表皮碎片的细胞结构,鉴定动物取食种类,但该技术的难点之一是植物碎片的识别,尤其是残留于粪便中的植物碎片识别。曹俊虎等[26]研究发现藏羚对干季牧草的消化率高于其它反刍动物,这有可能增加对粪样中植物碎片的识别难度,虽然有些植物种类的表皮细胞因具有特定的形态结构可以识别到种,如唐古韭等但大多数植物只能识别到属[4]。本试验采用没有粘性的饱和 NaCl溶液代替Hoyer's装片介质进行装片和镜检,显著地减少了植物碎片卷曲率和重叠率,有效提高了植物碎片的识别效果[4,5]。作为食性定量分析方法,粪便组织方法存在一定范围的偏差,而这与动物对各种植物的不同消化程度和食草动物自身的生理差异有关。同时,野外条件下,藏羚活动范围非常大,兼具迁徙习性,其食物组成相对难以确定到种。本试验中藏羚被圈养于驯养基地内,活动范围相对较小,而根据已经掌握藏羚活动范围内所有的植物种类,研究其在驯养基地的食物组成更有针对性,因此具有更高的属、种鉴定的准确性及说服力。
动物能够通过自己的适应和行为调节,而使得它们更能适应食物匮乏的生境,进而得以存活并不断生息,藏羚在这一方面更加突出。因此,其仍能对严酷的自然环境产生良好的适应。综上所述,基地藏羚的食物主要由禾本科、莎草科、菊科、十字花科和豆科等植物组成,其食物谱的组成与可可西里地区相比在科的分类水平上没有明显变化,不管是在可可西里还是在基地,禾本科植物在藏羚的食物谱中仍然占有较大的比例。因此,藏羚的食性没有发生明显变化,铁卜加地区可以作为藏羚异地驯养的基地。
[1] 李俊年,刘季科. 植食性哺乳动物食物选择的生态学意义及行为机制[J].应用生态学报,2003,14(3):439-442.
[2] 李由明,黄翔鸽,刘楚吾. 碳氮稳定同位素技术在动物食性分析中的应用[J]. 广东海洋大学学报,2007,27(4):99-103.
[3] 郑荣泉,鲍毅新. 有蹄类食性研究方法及研究进展[J]. 生态学报,2004,24(79):1532-1539.
[4] 曹伊凡,苏建平,连新明,等. 可可西里自然保护区藏羚羊的食性分析[J]. 兽类学报,2008,28(1):14-19.
[5] 曹伊凡,苏建平.一种用于食草动物粪便显微组织分析的临时装片新技术[J].兽类学报,2006,26(4):407-410.
[6] 武素功,冯祚建.青海可可西里地区生物与人体高山生理[M]. 北京:科学出版社,1996.
[7] 郑生武. 中国西北地区珍稀濒危动物志[M]. 北京:中国林业出版社. 1994.
[8] Wang S, Yang C F, Zheng G M. China red data book of endangered animal[M]. Beijing: (Mammalia) Science Press, 1998:341-344.
[9] 冯祚建. 喀喇昆仑山-昆仑山地区兽类资源的现状与保护[J]. 自然资源学报, 1990, 5(4):343-353.
[10] 冯祚建,郑昌林,蔡桂全. 西藏东南部兽类的区系调查[J]. 动物学报,1980,26(1):91-97.
[11] 谷景和. 新疆东昆仑-阿尔金山地区的有蹄类动物[J]. 干旱区研究,1987(3):56-68.
[12] Cai G Q, Liu Y S,O'ara B W. Observations of large mammals in the Qaidam Basin and its peripheral mountainous areas in the People's Republic of China[J]. Canadian Journal of Zoology, 1990, 68: 2021-2024.
[13] Harris R B, Pletscheer K H. Status and trend of Tibetan plateau mammalian fauna, Yeniugou China[J]. Biological Conservation, 1999, 87: 13-19.
[14] Ruan X, He P, Zhang J, et al. Evolutionary history and current population relationships of the chiru (Pantholops hodgsonii) inferred from mtDNA variation[J]. Journal of Mammalogy, 2005, 86: 881-886.
[15] Schaller G B. Wildlife of the Tibetan Steppe[M]. Chicago: University of Chicago press,1998.
[16] 裘 丽,冯祚建. 青藏公路沿线白昼交通运输等人类活动对藏羚羊迁徙的影响[J]. 动物学报,2004,50(4):669-674.
[17] 杨奇森,夏 林,吴晓民. 青藏铁路线上的野生动物通道与藏羚羊保护[J]. 生物学通报,2005,40(5):15-17.
[18] 郑中朝. 藏羚羊纤维特性的研究[J]. 中国草食动物, 2001, 3(4):14-16.
[19] Fox J, Bardsen B J. Density of Tibetan antelope, Tibetan wild ass and Tibetan gazelle in relation to human presence across the Chang Tang Nature Reserve of Tibet[J]. China Acta Zoological Sinica, 2005, 51: 586-597.
[20] Schaller G B, Ren J R. Effects of a snowstorm on Tibetan antelope[J]. Journal of Mammalogy, 1988, 69: 631-634.
[21] 连新明,苏建平,张同作,等. 可可西里地区藏羚的社群特征[J]. 生态学报,2005(25):1 341-1 346.
[22] 孙 平, 于鸿浩, 赵新全, 等. 异地半圈养藏羚卧息行为适应性的初步研究[J]. 四川动物,2011, 31(1): 1-5.
[23] 孙 平, 于鸿浩, 赵新全, 等. 青藏高原异地半圈养藏羚警戒行为的适应性研究[J]. 动物学研究,2012, 32(5): 561-565.
[24] Schaller G B, Ren J, Qiu M. Observations on the Tibetan antelope (Pantholops hodgsoni) [J]. Applied Animal Behaviour Science, 1991, 29:361-378.
[25] 殷宝法,淮虎银,张镱锂,等. 可可西里地区藏羚羊、藏原羚和藏野驴的营养生态位[J]. 应用生态学报,2007,18(4):766-770.
[26] 曹俊虎,徐世晓,赵新全,等. 藏羚羊冷季对干物质的消化效率[J]. 兽类学报, 2007,27 (2):206-208.
[27] Harris R B, Miller D J. Overlap in summer habitats and diets of Tibetan plateau ungulates[J]. Mammalis, 1995, 59: 197-212.
[28] 拉 旦,马恩洲. 铁卜加地区大气降水、土壤湿度与牧草产量的关系[J]. 青海畜牧兽医杂志, 1998(3): 21-22.
[29] Anthony R G, Smith N S. Comparison of rumen and fecal analysis to describe deer diets[J]. Journal of Wildlife Management, 1974, 38: 535-540.
[30] 蔡联炳,王世金,李建华. 中国主要禾本科植物叶片表皮细胞、表皮附属物的常见类型及其分类价值的初步探讨[M]//高原生物学集刊.第10集. 北京:科学出版社,1991:1-14.
[31] 康 乐,陈永林. 草原蝗虫食料植物叶片表皮显微结构的研究[M]//草原生态系统研究.第4集. 北京:科学出版社,1992:125-139.
[32] 李正理,张新英.植物解剖学[M]. 北京:高等教育出版社,1983.
[33] 宋延龄,巩会生,曾治高,等. 鬣羚食性的研究[J]. 动物学杂志,2005,40(5): 50-56.
[34] 梁 亮,张代贵,徐 剑,等. 喀斯特灌丛草地武雪山羊的觅食行为与喜食性[J]. 生态学杂志,2006,25(12):1524-1527.
[35] 吴建平,单继红,李言阔. 小兴安岭地区原麝冬季食性研究[J]. 兽类学报,2007,27(1):58-63.
[36] 王 岭,王德利. 放牧家畜食性选择机制研究进展[J]. 应用生态学报,2007,18(1):205-211.
[37] Crawford H S. Seasonal food selection on digestibility by tame white-tailed deer in central in Maine[J]. Journal of wildlife management, 1982, 46: 974-982.
Eating Habits of Tibetan Antelope in ex situ Conservation Based on Fecal Micro-histological Analysis
SUN Ping1,GUO Yu-ping2,XIONG Jian-li1,ZHANG Ji-liang1,WANG Zhan-bin1,ZHAO Xin-quan3*
(1.AnimalScienceandTechnologyCollege,HenanUniversityofScienceandTechnology,LuoyangHenan471003,China;2.Collegeofphysicaleducation,Anyangnormaluniversity,Anyang,Henan455000,China;3.NorthwestPlateauInstituteofBiology,ChineseAcademyofSciences,Xining,Qinghai810001,China)
By fecal micro-histological analysis, we studied the diet of Tibetan antelope in Tiebujia area, Qinghai Province,then compared it with these in Kekexili Natural Reserve, and tried to find the possibility protecting antelope via ex situ conservation.Comparisen between fecal samplesshowed that four kinds of species Carex tristachya, Leontopodium leontopodioides, Poa indattenuata, and koeleria cristata were the main food diet of Tibetan antelopes in Tiebujia area,accoccnting for 18%,23%,11% and 8% of total,respectively. Anyway, stems of C. tristachya, leaves of L. leontopodioides, fruits of Thlaspi arvense, and flowers of Sphallerocarpus gracilis, were the most favorite parts in the food of antelopes. Other families accounting for higher proportion were as follows: Gramineae (43%), Cyperaceae (25%), Compositae (14%), Leguminosae (7%), and Cruciferae (3%), respectively. In conclusion, Tibetan antelope showed similar food diet of monocoty-leden in Tiebujia with that in Kekexili area.
Tibetan antelope; fecal micro-histological analysis; ex situ conservation; food habits
2014-03-08,
2014-04-26
国家自然科学基金资助项目(30500073,30870370);青海省重点科技攻关项目(No.2006-N-152)
孙 平(1975-),男,山东邹城人,博士,副教授,主要从事动物生态方面的研究。 E-mail: pingsunny@msn.com
*[通讯作者] 赵新全(1959-),男,陕西扶风人,博士,研究员,主要从事草地生态和动物生态方面的研究。 E-mail: xqzhao@yahoo.com.cn
S811.6
A
1005-5228(2015)01-0070-06
