IL-27与LTB4在非小细胞肺癌中的表达及意义*
2015-04-19杨晓东马雪梅孙云晖林杨鲍文华
杨晓东 马雪梅 孙云晖 林杨 鲍文华
肺癌是一种主要起源于支气管黏膜或腺体的恶性肿瘤,而非小细胞肺癌约占所有肺癌的80%,具有高度侵袭性与转移的生物学特征,但病因至今尚不完全明确,其发生是一个复杂的生物学过程。本项实验采用免疫组化S-P法对IL-27与LTB4在肺癌标本中表达情况和相互关系进行研究。探讨两者在非小细胞肺癌发生、发展中的作用,为肺癌的临床诊治提供一种理论依据及治疗方法。
1 资料与方法
1.1 一般资料 实验标本选取本院2012-2014年手术切除的肺组织石蜡标本60例,均已确诊为非小细胞肺癌。女28例,男32例,年龄43~77岁,平均58.3岁。按1999年WHO肺癌分类标准,其中鳞癌34例、腺癌26例;高分化20例、中分化24例、低分化16例;按UICC1997肺癌分期标准分期,Ⅰ+Ⅱ期38例、Ⅲ期22例。入选标准:所有分类及分期均执行目前国际通用标准;所有标本术前均未接受任何抗肿瘤治疗;所有标本均重新证实为非小细胞肺癌。同时选取25例正常肺组织作为对照研究。
1.2 主要试剂 IL-27抗体、LTB4抗体、S-P试剂盒为通用型。
1.3 方法 免疫组化采用链霉菌抗生物素蛋白-过氧化物酶(S-P)法。实验标本采用10%福尔马林固定,常规石蜡包埋,切片机上作4 μm厚连续切片,常规脱蜡、水化、抗原修复及血清封闭等处置,分别与IL-27、LTB4抗体免疫组化染色,DAB显色,同时把已知的阳性非小细胞肺癌组织切片在相同情况下染色作为阳性对照,所有操作方法均严格执行S-P试剂盒说明书。
1.4 结果判断 所有结果均由本院病理专业的医师统计。IL-27以胞核染色为主,LTB4以胞膜或胞浆为主,呈棕黄色颗粒状染色,以清晰的棕黄色颗粒作为阳性表达细胞。取五个不同的视野,在400倍光镜下各自计数200个肿瘤细胞,以阳性细胞/同类细胞%为标准,将免疫组化结果分为:阴性(-):阳性细胞数<10%;弱阳性(+):阳性细胞数10%~25%;中度阳性(++):阳性细胞数25%~50%;强阳性(+++):细胞数>50%。前两者为低表达,后两者为高表达。
1.5 统计学处理 实验数据采用SPSS 16.0统计软件进行分析,计数资料比较采用 X2检验,两种蛋白表达的相关性采用Spearman相关性分析,以P<0.05为差异有统计学意义。
2 结果
2.1 IL-27与LTB4在非小细胞肺癌和正常肺组织中的表达水平 在非小细胞肺癌组织中IL-27与LTB4总阳性表达率分别为41.67%、63.33%,各种蛋白阳性表达率在肺癌组织与正常肺组织间的差异均有统计学意义(P<0.05),见表1。
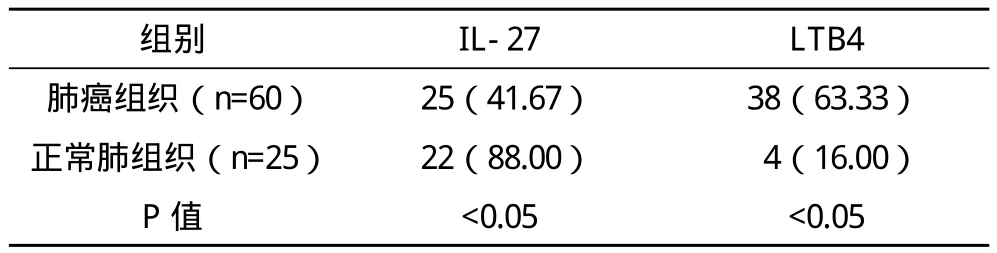
表1 IL-27与LTB4在肺癌和正常肺组织中表达情况的比较 例(%)
2.2 IL-27与LTB4的表达与非小细胞肺癌临床病理特征之间的关系 IL-27在肺癌组织中的阳性表达率为41.67%,其表达水平与肺癌的细胞分化程度和TNM分期相关(P<0.05),与肺癌患者的组织学类型无密切关系(P>0.05);LTB4在肺癌组织中的阳性表达率为63.33%,其表达水平与肺癌患者的组织学类型、TNM分期无密切关系(P>0.05),而与肺癌细胞的分化程度密切相关(P<0.05)。见表2。
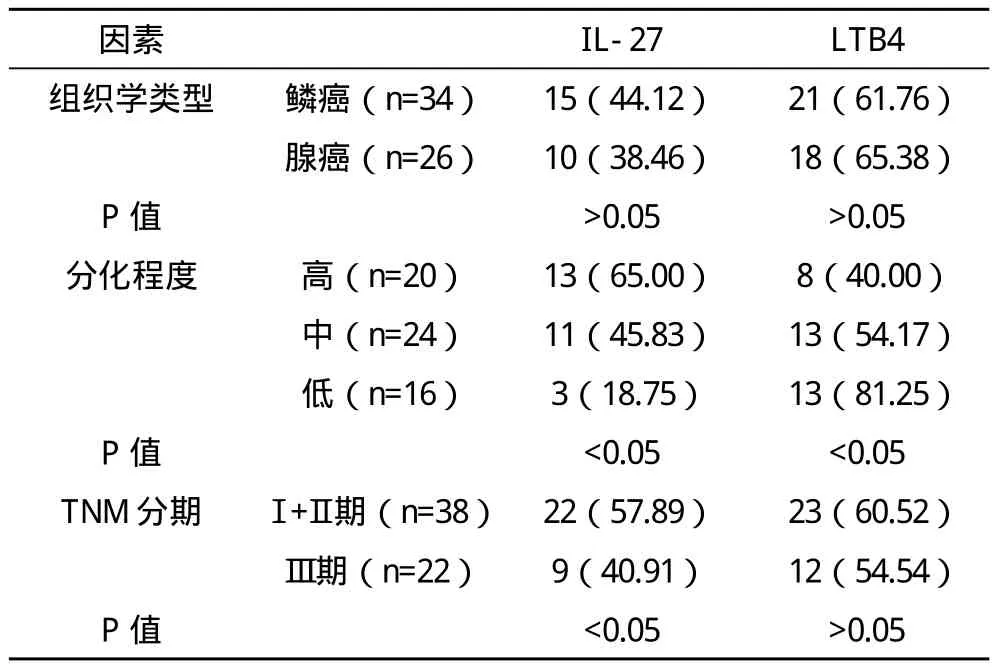
表2 IL-27与LTB4的表达与肺癌病理特征之间的关系情况
2.3 IL-27与LTB4表达的相关性 IL-27与LTB4之间相关性统计表明两者间有明显相关性,Spearman相关系数为-0.624(P<0.05)。两者在肺癌组织中的阳性共表达情况见表3。
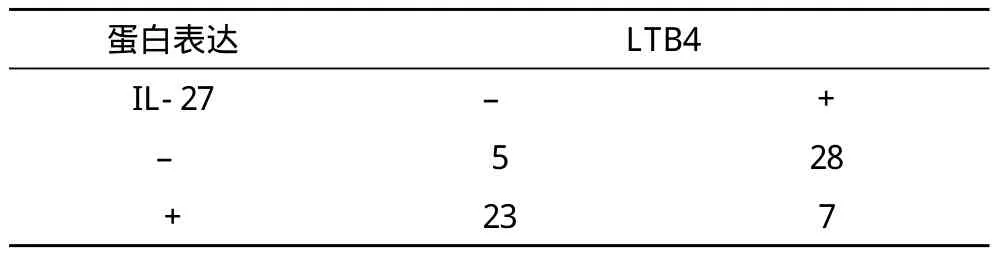
表3 IL-27与LTB4在肺癌组织中的阳性共表达情况 例
3 讨论
白 细 胞 介 素 27(interleukin-27,IL-27)是 在2002年Pflanz等发现的IL-6/IL-12家族的新型细胞因子[1],它由EB病毒诱导基因3亚基和p28亚基组成,主要通过调节Th1、Th2细胞及相关细胞因子参与自身免疫性疾病达到促进和抑制炎症的双重性质。目前关于IL-27与各种肿瘤发病机制的研究在我国日趋完善[2],IL-27与前列腺癌及其骨转移、食管癌淋巴转移、黑色素瘤等肿瘤密切相关[3],在肺癌方面,转染IL-27的小鼠肺腺癌实验已经进行,研究表明:肿瘤特异性CD8+T细胞通过IL-27调节参与机体抗肿瘤或者在活体直接起到抗肿瘤增殖作用[4]。血管形成作用和NK细胞被IL-27抑制,从而抑制了肿瘤生长和转移[5]。并且最近研究表明免疫炎症反应在肿瘤发生发展中起着重要作用[6],本项研究证实IL-27阳性表达率在非小细胞肺癌组织中为41.67%,显著低于正常肺组织(88.00%)(P<0.05),而且随着肺癌的细胞分化程度降低IL-27的阳性表达也随之下降(P<0.05),因此IL-27可能是一种潜在的抗肿瘤细胞因子,IL-27主要通过CD8+T细胞,分泌IFN-r发挥抗肿瘤作用,对免疫原性高、低不同的肿瘤细胞发挥抑制肿瘤生长和转移作用,尤其对免疫原性低的肿瘤发挥无可比拟的作用[7],其可以通过抑制血管形成来抑制肿瘤的增值。IL-27尚可不依赖免疫细胞而发挥抗肿瘤效应,并且目前研究一致认为在肿瘤细胞中过表达IL-27可以产生免疫保护和免疫记忆。因此IL-27基因可能成为肿瘤治疗的靶基因,认定是肿瘤治疗的新靶点[8],当前白介素药物治疗主要立足于感染性疾病治疗[9],但对于肺癌治疗提供了前提依据,并有望成为继手术治疗、化疗及放疗后的一种新的肺癌治疗模式。
白三烯B4(leukotriene B4,LTB4)作为花生四烯酸的代谢产物之一,是一种潜在的炎症介质和免疫反应调节剂,其代谢通路主要为:在各种酶的催化下,花生四烯酸转变成环化物白三烯A4(LTA4)。LTA4在 LTA4水解酶作用下形成LTB4。LTA4水解酶和5-脂氧合酶是白三烯合成过程中具有调节作用的酶[10]。LTB4受体有两种亚型即BLT1和BLT2,BLT1少量表达于脾脏和胸腺,其主要在白细胞中呈高表达。BLT2在脾脏中表达最高,卵巢、肝脏、白细胞中次之。另外,BLT2在CD+的淋巴细胞和单核细胞中也均有表达[11]。LTB4与食管癌、结肠癌、口腔癌、胰腺癌、人间变性大细胞淋巴瘤等多种肿瘤关系密切[12-13]。本项研究证实非小细胞肺癌组织中LTB4阳性表达率为63.33%,显著高于正常肺组织(16.00%)(P<0.05),而且LTB4的阳性表达随肺癌的细胞分化程度降低而升高(P<0.05),分析LTB4可能为一种致癌或促癌因子,在肺癌发生、发展中起着一定的作用。Chen等[14]对LTB4和肿瘤发生的机制研究认为与炎症相关的致癌作用有关:第一、炎症细胞、上皮细胞所引起的LTB4旁分泌生长刺激效应、自分泌生长刺激效应两者均作用于癌细胞和癌前病变。第二、LTB4受体与信号分子形成正反馈,从而加强了LTB4对炎症细胞的作用。其作用机制可能还有许多不明之处,但目前较确切的是LTB4参与肺癌发病中,而其受体拮抗剂则与其发挥作用相反。研究表明,LTB4受体抑制剂对肿瘤有明显抑制作用,因此如能应用于临床将对肿瘤治疗有着极大发展前景[15]。目前很多成熟的白三烯抑制剂已经在临床广泛应用[16],针对肿瘤的制剂亦在研制中。
IL-27与LTB4的阳性表达在本研究中表明与患者性别、年龄等无相关性。IL-27肺癌低分化组的阳性率为18.75%明显低于高、中分化组均数55.42%(P<0.05),其阳性表达随肿瘤病理分化程度的降低而降低(P<0.05),spearman相关性分析显示:在非小细胞肺癌组织中IL-27的表达与TNM分期成正相关(r=0.654,P<0.001),而与组织类型无相关性(P>0.05)。LTB4在非小细胞肺癌组织中呈高表达,其表达水平与肺癌的细胞分化程度负相关(r=-0.754,P<0.001),与病理学类型及TNM分期无相关性(P>0.05)。在此实验组非小细胞肺癌组织中,IL-27、LTB4表达经统计学分析结论表明两者之间呈负相关性(r=-0.624,P<0.001)。
本项研究证实了在肺癌复杂发病过程内,IL-27与LTB4均参与了肺癌发生、发展的病理生理过程,因此笔者对两者进行联合检测既丰富了肺癌发病机制理论也将对抗肿瘤药物研制起到一定启发,从而势必对肺癌的临床早期发现及早期诊治起到影响意义。
[1] Pflanz S,Timans J C, Cheung J, et al.IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein, induces proliferation of naive CD4(+)T cells[J].Immunity, 2002, 16(6):779-790.
[2] Hu T, Zhao H, Wang K, et al.Association of IL-27 polymorphisms and cancer risk in Chinese population[J].Recept Signal Transduct Res,2014, 22(7):1-4.
[3] Di Carlo E, Sorrentino C, Zorzoli A,et al.The antitumor potential ofinterleukin-27 in prostate cancer [J].Oncotarget, 2014, 5(21):10 332-10 341.
[4] Salcedo R, Stauffer J K, Lincoln E, et al.IL-27 mediates complete regression of orthotropic primary and metastatic murine neuroblastoma tumors:role for CD8+T cells[J].Immunol,2004, 173(12):7170-7182.
[5] Oniki S, Nagaih, Horikawa T, et al.Interleukin-23 and interleukin-27 exert quite different antitumor and vaccine effects on poorly immunogenic melanoma[J].Cancer Res, 2006, 66(12):6395-6404.
[6] Elinav E, Nowarski R, Thaiss C A, et al.Inflammation -induced cancer:crosstalk between tumous, immune cells and microorganisms[J].Nat Rev Cancer 2013,13(11):759-771.
[7] Wynick C, Petes C, Gee K.Interleukin-27 mediates inflammation during chronic disease[J] .Interferon Cytokine Res, 2014, 34 (10):741-749.
[8] Grivennikov S I, Greten F R,Karin M.Immunty, inflmmation and cancer [J] Cell,2010,140(6):883-889
[9]王雪斐,朱敏.白介素-11与维生素B12联合应用在化疗后口腔黏膜炎患者中的疗效观察[J].中国医学创新,2012,9(6):121-122.
[10] Wejksza K, Lee-Chang C, Bodogai M.Cancer-produced metabolites of 5-lipoxygenase induce tumor-evoked regulatory B cells via peroxisome proliferator-activated receptor α[J] .Immunol, 2013,190(6):2575-2584.
[11] Watanabe M, Machida K, Inoue H.A turn on and a turn off:BLT1 and BLT2 mechanisms in the lung [J] .Expert Rev Respir Med, 2014,8(4):381-383.
[12] Kim H, Park G S, Lee J E, et al.A leukotriene B4 receptor-2 is associated with paclitaxel resistance in MCF-7/DOX breast cancer cells [J].Cancer, 2013,109(2):351-359.
[13] Seo J M, Park S, Kim J H.Leukotriene B4 receptor-2 promotes invasiveness and metastasis of ovarian cancer cells through signal transducer and activator of transcription 3 (STAT3)-dependent upregulation of matrix metalloproteinase 2[J].Bio Chem, 2012,287(17):13 840-13 849.
[14] Chen X, Wang S, Wu N, et al.Leukotriene A4 hydrolase as a target for cancer prevention and therapy[J].Curr Cancer Drug Targets,2004,4(3):267-283.
[15] Tong W G, Ding X Z, Talamonti M S, et al.Leukotriene B4 receptor antagonist LY293111 induces S-phase cell cycle arrest and apoptosis in human pancreatic cancer cells[J].Anticancer Drugs, 2007, 18(5):535-541.
[16]李剑,赵晓宇,吴泰华.白三烯受体拮抗剂对吸烟哮喘患者病情控制的影响[J].中国医学创新,2014,11(16):4-6.
