棉花、苜蓿细根生长和死亡的季节变化
2015-04-15任爱天鲁为华杨洁晶马春晖
任爱天,鲁为华,杨洁晶,马春晖
(石河子大学动物科技学院,新疆 石河子832003)
棉花、苜蓿细根生长和死亡的季节变化
任爱天,鲁为华*,杨洁晶,马春晖
(石河子大学动物科技学院,新疆 石河子832003)
以棉花和紫花苜蓿为研究对象,以现存量、生长速率(G, cm/cm3·d)、死亡速率(M, cm/cm3·d)和生死之比(R)为基本参数,采用微根管技术对棉花和苜蓿细根的生长、死亡以及周转进行动态研究。结果表明,在生长季内,棉花平均细根现存生物量为8.025 mm/cm3,2011年苜蓿为14.830 mm/cm3,高于2012年苜蓿(8.026 mm/cm3),棉花细根G和M分别为0.056和0.011 mm/(cm3·d),2011年苜蓿分别为0.026和0.007 mm/(cm3·d),2012年分别为0.041和0.028 mm/(cm3·d)。棉花G变化趋势为5月底-7月中旬迅速下降,7月中旬到9月初上升,此后下降,2011和2012年苜蓿G变化趋势一致,分别在6月中旬-7月中旬和8月初-9月初形成峰值。棉花和苜蓿M相似,均是在6月中旬到7月中旬降低,此后开始增加,在生长季末达到最大。R在生长季呈逐渐减少的趋势,但棉花在生长季末R>1,苜蓿则是R<1。用最大细根现存量和平均细根现存量估算得到细根周转率不同,以平均细根现存量得到细根周转率比以最大细根现存量增加了22%。通过对棉花和苜蓿细根比较研究,棉花和苜蓿在不同季节细根现存生物量和生长动态不同,棉花呈单峰曲线,苜蓿呈双峰曲线,而棉花和苜蓿死亡动态则是随着季节变化持续升高,同时棉花以较高的周转速率满足了植物对水分和养分的需求。
细根;生长率;死亡率;周转;微根管
植物根系作为植物与土壤接触的器官,有着非常重要的生理学功能,包括从土壤中吸收水分和养分等。此外,植物根通过自身的生长(封存的碳和养分)和死亡(释放碳和养分)对陆地生态系统中养分的循环做了很大的贡献[1]。植物的根系根据其直径可以分成不同层次,其中细根通常被定义为直径小于2 mm的根,在目前,关于细根生物量、生长、周转、季节动态及对陆地生态系统生物化学循环的影响已逐渐成为研究焦点[2]。细根虽然对环境因子(如温度、土壤养分等)特别敏感,并且有相对短的寿命[1]。但是在地下生态过程中起着非常重要的作用,主要表现在:细根有着巨大的吸收表面积,是植物吸收水分和养分的重要器官;虽然细根生物量占总根系生物量的一小部分,但是许多研究表明,细根的生长占到净初级生产的26%~33%[1];细根周转进入土壤中的碳氮等营养元素远高于地上凋落物的输入量。因此,估计细根的生物量以及生长和死亡能够很好地了解生态系统中碳循环和生物地球化学动态变化[3]。同时研究认为细根的周转有助于植物初级生产物的沉积,是植物生态系统营养动态变化和碳封存的重要组成部分[4]。因而准确估算细根生产和周转是研究陆地生态系统碳分配格局的核心环节[2]。
近年来,随着新疆养殖业快速发展,紫花苜蓿(Medicagosativa)种植面积逐年加大,但紫花苜蓿连续生长多年会导致下层土壤水分超常利用,从而导致土壤干旱层向下延伸。而且还会引发自身生长衰败。同时在新疆农作物的种植主要以棉花(Gossypiumhirsutum)为主,长期棉花的连作导致农田病虫害大面积发生,土壤肥力下降,部分养分亏缺,因此将苜蓿与棉花进行轮作,对后茬作物提供大量的营养物质,起到增产及改善作物品质的作用[5],可以缓解和恢复苜蓿地的水分亏缺状况[6],对维持生态系统营养的平衡和功能恢复发挥了重要的作用。而且种植多年的苜蓿其根瘤菌和大量死亡的细根可以增加土壤的腐殖质和有机质,苜蓿翻耕后,将有利于农田作物水分的利用,提高后茬作物土壤氮素的有效性。目前对苜蓿与棉花草田轮作的研究鲜有报道,尤其对草田轮作中作物根系的生产和周转研究很少,而根系特征是牧草水分和养分管理、刈割和放牧利用的基本依据,与土壤耕作及作物种植制度关系密切,因此,利用微根管技术对轮作系统中不同作物细根生长变化进行动态研究,为草田轮作系统中养分的循环以及农地资源的合理、高效、持续利用提供科学依据。
1 材料与方法
1.1 试验地概况和样地的设置
试验于2011年在石河子市西郊石河子大学试验场二连(东经 86°03′,北纬 45°19′,海拔412 m) 进行。试区棉花和苜蓿生长季节4-10月降水总量为139.8 mm,年平均日照时间达2865 h,大于10℃年积温为3463.5℃,大于15℃年积温为2960.0℃,无霜期达到170 d,供试土壤为灌耕灰漠土,pH值8.1,质地为砂壤土,有机质15.6 g/kg,全氮0.76 g/kg,全磷(P2O5)1.54 g/kg,碱解氮83.3 mg/kg,速效磷(P) 30.4 mg/kg。使用棉花品种新陆早13号,4月播种,4月29日出苗,60-30-60-30 cm宽窄行宽膜覆盖种植。使用苜蓿种子为阿迪那多叶苜蓿品种(Adrenalin,由加拿大BrettYoung公司生产),建植时布设研究小区,每个小区面积均为5 m×8 m,其中每个小区中间行苜蓿为观测行,两边为保护行。
1.2 微根管安装及数据采集
2011年建植期间,在棉花和苜蓿试验区随机各选择6块样地,每块样地均采用滴灌方式。每块样地微根管根据白文明等[7]介绍方法进行安装和数据采集。数据采集工作从2011年5月24日开始,棉花是到2011年10月初为止。由于苜蓿为多年生作物,而且2011年苜蓿是建植年,因此为了进一步比较不同年限苜蓿细根生长动态变化,对苜蓿进行为期两年的观测,分别是在2011年10月初和2012年10月初结束,两年期间采用同样的处理方法进行数据的采集,苜蓿生长过程中,当小区内有10%植株开花时开始收获,收获时间分别在5月底,7月中旬和9月初。
1.3 细根数据的整理
为了与常用的单位土壤体积为基础的细根动态数据进行比较,本文以单位土壤体积的根长密度(root length density production,RLD,cm/cm3)作为基本参数[8]。RLD=(RL×sinθ)/(W×DOF×D)。式中,RL(cm)为观测窗中观测到的细根根长(root length);W(cm)为扫描图片宽度(width);DOF(cm)为田间深度(depth of field),本实验中DOF为100 cm,D为土层厚度(摄像头扫描可见的),一般为0.2~0.3 cm,本研究中D(cm)取0.2[9-10]。计算RLD时,考虑到微根管的安装角度与地表呈45°,需要将计算结果转换成垂直高度的单位体积根长密度(乘以sin45°)。
以单位时间单位土壤体积的细根生长量和死亡量表示细根的生长速率(growth rate, G)和死亡速率(death rate, M)[11]。G(M)=ΔRLD增(减)/T。式中,ΔRLD增(减)为相邻两次观测值之间单位土壤体积的棉花、苜蓿细根生长量(死亡量),T为相邻两次观测之间的间隔天数(d),G和M的单位均为mm/(cm3·d)。
为了表示细根生长量与死亡量的相对大小,本研究同时引入细根生死之比(即细根生长速率与死亡速率之比,R)这一参数:R=G/M。由定义可见,R=1,生长量与死亡量相等;R>1,生长量大于死亡量;R<1,生长量小于死亡量。
参照Burton等[12]的方法对生长季细根生长量和死亡量进行估算,在本研究中,生长季细根生长量是指从5月至10月各次观测得到的细根(已有细根和新生细根)长度增加值之和(mm/cm3),生长季细根死亡量是指各次观测得到的细根(因细根死亡脱落和昆虫取食等)长度减少值之和(mm/cm3)。细根现存量以每次观测到的单位土壤体积活细根长度来表示。细根周转估计采用以下公式:
周转率(a)=年细根生长量(mm/cm3·a)/年细根平均现存量(年细根最大现存量)
1.4 数据处理
用Excel软件整理获得的微根管数据。采用方差分析方法分析细根生长差异和死亡差异,采用SPSS软件进行统计分析。
2 结果与分析
2.1 细根现存量
不同月份苜蓿和棉花细根现存量存在一定的差异(表1)。其中棉花的现存量出现了1次峰值,而苜蓿细根现存量出现了2次峰值。棉花细根现存量在8月达到最大(11.019±0.691) mm/cm3,8月之后开始下降,到10月达到最低(5.029±0.828) mm/cm3。2011和2012年苜蓿现存量变化趋势趋于一致。均先在6月达到1次峰值,分别为(14.846±0.260)和(9.831±0.403) mm/cm3,此后下降,在8月达到另1次峰值,为(16.469±0.594)和(10.993±0.656) mm/cm3。此后开始下降,到10月降到(14.823±0.475)和(4.386±0.388) mm/cm3。

表1 棉花和苜蓿细根现存量季节变化
注:同列不同小写字母表示不同处理之间差异显著(P<0.05),下同。
Note: Values followed by different small letters are significantly different at 0.05 level, the same below.
2.2 细根生长速率和死亡速率的季节动态
在生长季节,棉花生长速率在5月底最大,为(0.108±0.012) mm/(cm3·d),此后在6月中旬到7月11日细根生长速率达到最低,为(0.014±0.003) mm/(cm3·d)(P<0.05)。此后开始上升,在8月6日到9月8日出现1次峰值,显著高于6月15日到7月11日,但与其他时期差异不显著(P>0.05)。两年苜蓿生长速率均出现双峰曲线,分别在6月中旬到7月11日和8月6日到9月8日出现。2011年苜蓿细根第2次峰值生长速率显著高于其他时期(P<0.05),而2012年苜蓿第1次峰值生长速率均高于其他时期,在整个生长季内,2012年苜蓿生长速率(0.041 mm/cm3·d)显著高于2011年(0.026 mm/cm3·d)(表 2)。
在2011-2012年生长季内,棉花和苜蓿出现相似的细根死亡速率动态变化(表2)。细根死亡速率均在季末达到最大,显著高于其他时期(P<0.05)。总体来看,均在生长初期较高,然后开始下降,此后缓慢上升,在生长季末达到最高,分别为0.038,0.024和0.089 mm/(cm3·d)。2012年苜蓿死亡速率(0.028 mm/cm3·d)显著高于2011年(0.007 mm/cm3·d)。

表2 棉花和苜蓿细根生长速率和死亡速率的季节动态变化
2.3 棉花和苜蓿生死之比(R)
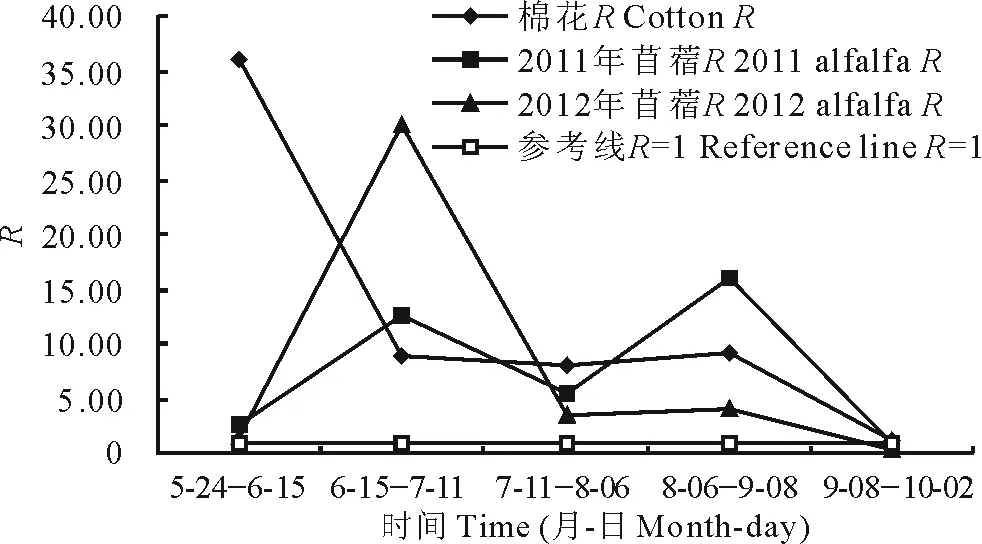
图1 细根生长速率与死亡速率的季节变化Fig.1 Seasonal changes of the ratio of fine root growth rate (G) and death rate (M)
就季节变化而言,棉花R呈逐渐减少的趋势(图1),自生长季节初到10月初,棉花R均大于1,说明在5月到10月之间,棉花的生长速率均大于死亡速率。然而苜蓿的R在生长季节内表现出一定动态变化,在9月8日到10月2日之前,R均大于1,在9月初以后,R开始小于1,说明除了生长季末之外,两年苜蓿细根均是生长速率大于死亡速率,由此可见,在生长期中,苜蓿的细根动态是一个以生长占优势的生死交织过程。
2.4 细根周转
由表3可知,棉花年细根生长量显著高于苜蓿,而2011和2012年苜蓿年细根生长量之间没有显著差异。细根现存量2011年苜蓿显著高于2012年苜蓿和棉花。细根周转棉花(1.03)高于苜蓿,2012年苜蓿周转率(0.76)高于2011年苜蓿(0.27)。利用细根平均现存量和最大现存量做分母估算得到细根周转率出现了一定的差异,使用平均现存量估算得到的周转率较使用最大现存量增加了22%。
3 讨论
苜蓿作为多年生作物,其根系生长发育决定了地上生物量以及利用年限[13],而细根的生长和死亡在根系生长中发挥巨大作用,但是在不同年际其细根变化存在一定差异。研究发现两年苜蓿细根现存量出现了一定的差异,可能是2011年是苜蓿建植年,到了第2年苜蓿为了获得更多的养分以及受刈割等因素的影响,主根进一步伸长变粗,横向生长加快[14-15],导致二级以及三级细根直径也逐渐变粗,形成较粗的根系,因此在第2年苜蓿细根现存量较第1年少。苜蓿生长速率以及死亡速率在2011和2012年均表现出相同的趋势,但是2012年苜蓿的生长速率和死亡速率均高于2011年,对细根周转研究也发现,2011年细根周转率较低,说明在2011年苜蓿低的周转就能满足植株对营养的需求,而在2012年随着苜蓿年限的增加和多次刈割等影响,苜蓿需要不断产生新的细根以替代旧的细根来吸收水分和营养物质,提高利用效率[16]。

表3 棉花和苜蓿年细根生长量、年现存量和周转
1) 年长度生长量/平均现存量Annual length production/Average length;2) 年长度生长量/最大现存量Annual length production/Max length.
在生长季节内,细根现存生物量出现一定程度的波动,在5-7月,棉花主要是主根和侧根的快速生长,此时细根生物量较低,到8月,棉花处于花铃期,此时棉花根系达到吸收的高峰期,细根的生长十分的旺盛,细根现存生物量达到了高峰,之后,9月直至10月,棉花处于吐絮期,根系的生理功能逐渐衰退[17],细根现存生物量逐渐下降,而苜蓿细根现存生物量在生长季节内出现两次峰值。在5月底,7月中上旬和9月初,由于苜蓿进行地上部分的收获,使得植株光合作用减弱,根系中分配到的碳水化合物降低,使得根系现存生物量降低,而在6月中旬和8月初,由于此时苜蓿地上部分生长旺盛,输入到根系碳水化合物较多,进而促进了根系的生长,而且夏季气温升高,土壤温度较高,利于细根的增加[18]。
已有研究表明,细根生长的季节动态表现为3种形态,即无明显季节变化,单峰曲线[19]和双峰曲线[20]。研究发现棉花的生长表现出单峰型,而苜蓿则表现出双峰曲线。在5月底,棉花幼苗开始生长,为满足叶和嫩枝的发育,需要提供大量的养分和水分,这就要迫使植物产生较高的细根生物量来满足自身生长需求[21]。因此在此阶段细根生产较高。然而苜蓿此时由于地上部分收获,使得供应细根的碳水化合物减少,细根的发生降低。在6月中旬到7月中旬,由于棉花主根和侧根的大量生长,此时细根发生减缓,此后,随着温度的上升,土壤温度的上升,促使细根的增加,同时此阶段棉花进入花铃期,根系吸收矿物元素和水的能力增加,促使细根的大量发生,而且此时期植物处于生长旺盛时期,促进光合产物的运输和分配[18],这一时期细根可以获得大量碳水化合物的供应。但是苜蓿由于在5月底地上部分收获,在6月中旬到7月中旬为了促使地上部分的快速生长,细根的发生增加,此后在7月中旬收获地上部分,细根的生长表现出相同的变化趋势。在9月以后,随着气温的降低,以及棉花处于吐絮期,地上叶片的脱落,同时细根获得的碳水化合物减少,细根生产下降,而苜蓿在此阶段又进行了地上部分的收获,因而细根生产迅速下降。
在本研究中发现在生长季节内棉花和苜蓿的细根死亡动态趋势基本一致。春季细根的死亡可能与储存的光合产物消耗有关,秋季细根大量死亡则与落叶和温度降低密切相关[11],而且在9月以后,由于棉花进入吐絮期,棉花需水强度降低,同时在此阶段根系生理功能减弱[17-22],但根系为维持细胞膜和酶活性所进行的呼吸作用依然消耗大量的碳,因此,植物为降低能量消耗而增加细根死亡量[18],而苜蓿又在9月以后进行地上部分收获,从而进一步加剧了细根的死亡。研究发现,无论是苜蓿还是棉花,细根M随生长季的变化表现出持续升高的趋势[19],对柠条(Caraganakorshinskii)和琵琶柴(Reaumuriasoongorica)群落细根的动态研究中也观察到了类似的现象[11],认为秋季土壤温度降低是导致细根大量死亡的主要原因[18]。
在生长季,棉花细根生死之比很悬殊 (5.09/1)。而苜蓿的生死之比为3.71和1.46。棉花生长速率在季末 (9月下旬) 仍很高(图 3),说明细根死亡节律较为滞后,这可能与地下气候滞后于地上气候以及地下环境因子比地上较为稳定有关[11]。而苜蓿在9月以后细根死亡速率超过生长速率,这可能是由于在生长季末,随着灌溉的停止,加之地上部分收获以后停止生长,导致了细根死亡的加快。随着生长季的迅速结束和冬季的到来,地下热量条件会更为不适。
在目前,估算细根周转通常用到细根最大现存量和细根平均现存量,而且大约2/3的研究采用最大现存生物量来估算细根周转[23]。本研究发现采用细根平均现存量得到的周转率较最大现存量增加了22%。2011年细根周转率较低,可能是细根管埋设后,在2011年土壤和微根管界面没有达到平衡,土壤紧实度、温度、湿度等的改变影响了根系生长[24],从而使得年细根生长量较低,同时在这一年苜蓿细根生长速率较低,所以不需要较高的周转来满足植株营养需求,而棉花和2012年苜蓿细根周转率较高,植物在生长过程中需要大量的营养物质和能量来保证高的细根生物量,但保持较高的细根周转可以使得植物以较低细根生物量满足植物旺盛的生命活动对水分和营养物质的需求[25],并保证对土壤水分和养分的有效吸收。
[1] Noguchi K, Sakata T, Mizoguchi T,etal. Estimating the production and mortality of fine roots in a Japanese cedar (CryptomeriajaponicaD. Don) plantation using a minirhizotron technique. Journal of Forestry Research, 2005, 10: 435-441.
[2] Ji W P, Wang J J, Zhao X C,etal. Fine root production and turnover ofAlhagisparsifoliacommunity in arid area of Xinjiang, Northwest China. Chinese Journal of Ecology, 2013, 32(10): 2635-2640.
[3] Zhi Q P, Chun W X, Dan D,etal. Comparison of the fine root dynamics ofPopuluseuphraticaforests in different habitats in the lower reaches of the Tarim River in Xinjiang, China, during the growing season. Journal of Forestry Research, 2012, 17:343-351.
[4] Gill R A, Jackson R B. Global patterns of root turnover for terrestrial ecosystems. New Phytologist, 2000, 147: 13-31.
[5] Liu P S, Jia Z K, Li J,etal. A study on optimal duration ofMedicagosativainM.sativa-grain crop rotation system in arid region of Southern Ningxia. Acta Prataculturae Sinica, 2008, 17(3): 31-39.
[6] Han L N, Ding J, Han Q F,etal. Effects of alfalfa-grain (oil) crop plowing rotation on soil moisture and crop yield in Loess Plateau. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2012, 28(24): 129-137.
[7] Bai W M, Cheng W X, Li L H. Applications of minirhizotron techniques to root ecology research. Acta Ecologica Sinica, 2005, 25(11): 3076-3081.
[8] Li J Y, Wang M B, Shi J W. Minirhizotron technique in measuring fine root indices: A review. Chinese Journal of Ecology, 2007, 26(11): 1842-1848.
[9] Shi J W, Yu Y Q, Yu L Z,etal. Application of minirhizotron in fine root studies. Chinese Journal of Applied Ecology, 2006, 17(4): 715-719.
[10] Bernier P Y, Robitaille G. A plane intersect method for estimating fine root productivity of trees from minirhizotron images. Plant and Soil, 2004, 265: 165-173.
[11] Wang M B, Chen J W, Shi J W,etal. The seasonal change patterns of production and mortality of fine roots in youngCaraganakorshinskiiplantation. Acta Ecologica Sinica, 2010, 30(19): 5121-5130.
[12] Burton A J, Pregitzer K S, Hendrick R L. Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests. Oecologia, 2000, 125(3): 389-399.
[13] Peng L Q, Li X Y, Qi X,etal. The relationship of root traits with persistence and biomass in 10 alfalfa varieties. Acta Prataculturae Sinica, 2014, 23(2): 147-153.
[14] Nan L L, Shi S L, Zhang J H. Study on root system development ability of different root-type alfalfa. Acta Prataculturae Sinica, 2014, 23(2): 117-124.
[15] Du M X, Zhou X R, Zhou Z Y,etal. Vertical root distribution characteristics ofAmorphafruticosaon southern Mu Us Sandy Land. Acta Prataculturae Sinica, 2014, 23(2): 125-132.
[16] Burke M K, Raynal D J. Fine root growth phenology, production and turnover in a northern hardwood forest ecosystem. Plant and Soil, 1994, 162(1): 135-146.
[17] Li Y Y, Liu H H, Chen J X,etal. Review of research on root of cotton. Crop Research, 2008, 22(5): 449-452.
[18] Pei Z Q, Zhou Y, Zheng Y R,etal. Contribution of fine root turnover to the soil organic carbon cycling aReaumuriasoongoricacommunity in an arid ecosystem of Xinjiang Uygur Autonomous Region, China. Chinese Journal of Plant Ecology, 2011, 35(11): 1182-1191.
[19] Chen J W, Wang M B, Shi J W. A comparative study of the spatial-temporal patterns of fine roots between young and matureCaraganakorshinskiiplantations. Acta Ecologica Sinica, 2011, 31(22): 6978-6988.
[20] Yong Z, Jia Q S, Janssens I A.etal. Fine root and litterfall dynamics of three Korean pine (Pinuskoraiensis) forests along an altitudinal gradient. Plant and Soil, 2014, 374: 19-32.
[21] Cheng Y H, Han Y Z, Wang Q C,etal. Seasonal dynamics of fine root biomass, root length density, specific root length and soil resource availability in aLarixgmeliniplantation. Acta Phytoecologica Sinica, 2005, 29(3): 403-410.
[22] Zuo Y B, Pang H C, Li Y Y,etal. Effects of plastic mulching on Etc, Kc and WUE of cotton in northern Shandong province. Chinese Journal of Agrometeorology, 2010, 31(1): 37-40.
[23] Brunner I, Bakker M R, Björk R G,etal. Fine-root turnover rates of European forests revisited: an analysis of data from sequential coring and ingrowth cores. Plant and Soil, 2013, 362: 357-372.
[24] Zhang X Q, Wu K H, Murach D. A review of methods for fine-root production and turnover of trees. Acta Ecologica Sinica, 2000, 20(5): 875-883.
[25] Wang Y J, Zhao X C, Lai L M,etal. Growth dynamic and turnover of fine root ofHaloxylonammodendroncommunities in Sangong river basin. Journal of Ecology and Rural Environment, 2013, 29(5): 666-670.
参考文献:
[2] 冀卫萍, 王健健, 赵学春, 等. 干旱区骆驼刺群落细根生产与周转. 生态学杂志, 2013, 32(10): 2635-2640.
[5] 刘沛松, 贾志宽, 李军, 等. 宁南旱区草粮轮作系统中紫花苜蓿适宜利用年限研究. 草业学报, 2008, 17(3): 31-39.
[6] 韩丽娜, 丁静, 韩清芳, 等. 黄土高原区草粮(油)翻耕轮作的土壤水分及作物产量效应. 农业工程学报, 2012, 28(24): 129- 137.
[7] 白文明, 程维信, 李凌浩. 微根管技术及其在植物根系研究中的应用. 生态学报, 2005, 25(11): 3076-3081.
[8] 李俊英, 王孟本, 史建伟. 应用微根管法测定细根指标方法评述. 生态学杂志, 2007, 26(11): 1842-1848.
[9] 史建伟, 于永强, 于立忠, 等. 微根管在细根研究中的应用. 应用生态学报, 2006, 17(4): 715-719.
[11] 王孟本, 陈建文, 史建伟, 等. 柠条人工幼林细根生长和死亡的季节变化. 生态学报, 2010, 30(19): 5121-5130.
[13] 彭岚清, 李欣勇, 齐晓, 等. 紫花苜蓿品种根部特性与持久性和生物量的关系. 草业学报, 2014, 23(2): 147-153.
[14] 南丽丽, 师尚礼, 张建华. 不同根型苜蓿根系发育能力研究. 草业学报, 2014, 23(2): 117-124.
[15] 杜明新, 周向睿, 周志宇, 等. 毛乌素沙南缘紫穗槐根系垂直分布特征. 草业学报, 2014, 23(2): 125-132.
[17] 李垚垚, 刘海荷, 陈金湘, 等. 棉花根系研究进展. 作物研究, 2008, 22(5): 449-452.
[18] 裴智琴, 周勇, 郑元润, 等. 干旱区琵琶柴群落细根周转对土壤有机碳循环的贡献. 植物生态学报, 2011, 35(11): 1182-1191.
[19] 陈建文, 王孟本, 史建伟. 柠条人工林幼林与成林细根动态比较研究. 生态学报, 2011, 31(22): 6978-6988.
[21] 程云环, 韩有志, 王庆成, 等. 落叶松人工林细根动态与土壤资源有效性关系研究. 植物生态学报, 2005, 29(3): 403-410.
[22] 左余宝, 逄焕成, 李玉义, 等. 鲁北地区地膜覆盖对棉花需水量、作物系数及水分利用效率的影响. 中国农业气象, 2010, 31(1): 37-40.
[24] 张小全, 吴可红, Murach D. 树木细根生产与周转研究方法评述. 生态学报, 2000, 20(5): 875-883.
[25] 王永吉, 赵学春, 来利明, 等. 三工河流域梭梭群落细根生长动态及周转. 生态与农村环境学报, 2013, 29(5): 666-670.
Seasonal change patterns in the production and mortality of fine roots in cotton and alfalfa
REN Ai-Tian, LU Wei-Hua*,YANG Jie-Jing, MA Chun-Hui
CollegeofAnimalScience&Technology,ShiheziUniversity,Shihezi832003,China
Roots have two functions, “source” and “sink”, which are important for substance and energy exchange between plants and soil. Understanding these processes requires studies of the dynamics of fine root growth and mortality. This study reports on the growth, death and turnover of fine roots in cotton (Gossypiumhirsutum) and alfalfa (Medicagosativa) using minirhizotron techniques over different seasons. The fine root growth rate [G, cm/(cm3·d)], death rate [M, cm/(cm3·d)] and the ratio of growth to death (R) were determined. Results showed that the average standing crop of cotton in the growing season is 8.0 mm/cm3, with that of alfalfa being 14.8 mm/cm3in 2011 and 8.0 in 2012. The averageGandMof cotton in the growing season are 0.056 and 0.011 mm/(cm3·d) respectively, with those of alfalfa being 0.026 and 0.007 mm/(cm3·d) in 2011 and 0.041 and 0.028 in 2012. TheGof cotton decreased rapidly from the end of May to mid-July, increased from mid-July to early September and then decreased. TheGof alfalfa showed a consistent trend in 2011 and 2012, peaking from mid-June to mid-July and from early August to early September. TheMof both cotton and alfalfa decreased from mid-June to mid-July and then increased to reach the maximum by the end of the growing season. TheRof cotton and alfalfa decreased gradually from May to October, with cottonR>1 and alfalfaR<1. The estimated turnover rate is different according to whether it is calculated in terms of maximum or mean biomass. Mean biomass calculations increased turnover rates by about 22% compared with maximum biomass. During the growing season, the standing crop andGof cotton and alfalfa showed unimodal and bimodal peaks respectively, but theMof both rose continuously. At same time, cotton had a higher turnover rate to meet water and nutrient requirements.
fine root; growth rate; death rate; turnover; minirhizotron
10.11686/cyxb2014319
http://cyxb.lzu.edu.cn
2014-07-21;改回日期:2014-10-09
兵团博士资金专项(2012BB017),国家牧草产业技术体系项目(CARS-35),石河子大学科学技术研究发展计划项目(RCZX201022)和石河子大学科技服务项目(4004103)资助。
任爱天(1989-),男,甘肃张掖人,在读硕士。E-mail: aitianren@163.om *通讯作者Corresponding author. E-mail: winnerlwh@sina.com
任爱天, 鲁为华, 杨洁晶, 马春晖. 棉花、苜蓿细根生长和死亡的季节变化. 草业学报, 2015, 24(6): 213-219.
Ren A T, Lu W H,Yang J J, Ma C H. Seasonal change patterns in the production and mortality of fine roots in cotton and alfalfa. Acta Prataculturae Sinica, 2015, 24(6): 213-219.
